
EFFECTS OF TWO ANTI-ESTROGEN DRUGS EXPOSURE ON LIPID METABOLISM OF FEMALE ZEBRAFISH (DANIO RERIO)
-
摘要: 为探究雌激素对雌鱼体内脂肪代谢的影响,研究分别使用50和250 μg/L的来曲唑(Letrozole、LET)与他莫昔芬(Tamoxifen、TAM)两种抗雌激素药物,构建了雌性斑马鱼(Danio rerio)雌激素缺乏模型和雌激素受体竞争抑制模型,并检测两种药物处理后斑马鱼肝脏、内脏和肌肉的甘油三酯(TG)含量变化以及肝脏内雌激素和脂肪代谢相关基因的变化。结果显示,低浓度LET处理后雌鱼肝脏和内脏TG显著上升(P < 0.05);高浓度TAM处理后肝脏TG含量显著降低(P < 0.05),其他各组处理TG均无差异。基因mRNA检测结果表明,两种浓度LET和TAM处理的雌性斑马鱼芳香化酶(CYP19A)表达均显著下调(P < 0.05),低浓度TAM暴露导致雌激素受体(ERα)表达显著下调(P < 0.05)。此外,两种浓度LET处理均引起了脂肪酸合成酶(FAS)表达显著上调,微粒体的TG转运蛋白(MTP)表达显著下调(P < 0.05);低浓度TAM引起了MTP表达显著下调(P < 0.05),而高浓度TAM组则引起了MTP表达显著上调(P < 0.05)。综合各相关指标,研究结果表明雌激素确实在雌性斑马鱼脂肪代谢中发挥作用,然而不同程度和方式的雌激素抑制会导致不同的脂代谢失调表现,这提示鱼体内雌激素紊乱所导致的脂代谢失调与雌激素浓度和作用通路上的受阻位点有关,并受到多重因子参与的内分泌调控网络的调节。Abstract: In higher animals, estrogen can regulate lipid metabolism of females; however, whether estrogen regulates fish lipid metabolism is unclear. To explore effects of estrogen on lipid metabolism in zebrafish, letrozole (LET) and tamoxifen (TAM) were treated female zebrafish (0.14±0.01) g for 5 weeks to measure triglyceride (TG) concentrations in liver, visceral and muscle and the expressions of the hepatic genes related to estrogen and lipid metabolism. The results showed that low dose LET significantly increased TG in liver and viscera but not in muscle. Low dose TAM did not impact No TG content in all tested tissues but high dose TAM significantly decreased liver TAG. LET and TAM significantly induced CYP19A mRNA, and low dose TAM decreased ERα. LET increased FAS but decreased MTP expression. DGAT had an increase tendency, and CPT-1 and PPARα had a decreased tendency. It suggests a dyslipi-demia symptom in the LET exposure groups. Low dose TAM reduced MTP and high dose TAM enhanced it (P < 0.05). These results showed important role of estrogen in lipid metabolism in female zebrafish with a complex dosageand tissue-dependent pattern.
-
Keywords:
- Estrogen /
- Lipid metabolism /
- Letrozole /
- Tamoxifen /
- Zebrafish
-
硫胺素, 也称维生素B1, 以焦磷酸硫胺素(TPP)的形式参与动物体内碳水化合物氧化分解及其转化为脂类等代谢反应[1], 是水生动物所必需的水溶性维生素之一, 如对鲤鱼(Cyprinus carpio)[2]、虹鳟(Salmo gairdneri)[3]、大菱鲆(Scophthalmus maximus)[4]和斑点叉尾鲙(Ictalurus punctatus)[5]等的研究表明, 当饲料中硫胺素满足不了需求时, 鱼类会有厌食、生长缓慢、运动失常、鳍出血和体色异常等症状。已有的关于鱼类对硫胺素需求量的研究表明硫胺素的最适需求量大菱鲆[4]为0.6-2.6 mg/kg、虹鳟[3]为1.0 mg/kg、斑点叉尾鲙[5]为1.0 mg/kg、奥尼罗罗非鱼(Oreochromis niloticus)[6]为3.5 mg/kg、建鲤(Cyprinus carpio var. Jian)[7]为1.02 mg/kg、军曹鱼(Rachycentron canadum)[8]为6.80-8.56 mg/kg、草鱼(Ctenopharyngodon idel-la)[9]为1.16-4.49 mg/kg、吉富罗非鱼(GIFTOreochromis niloticus)[10]为1.16-2.06 mg/kg。参照以往的试验方法, 通常将增重率[9]、血液或者血清中的乳酸和丙酮酸含量[3, 6, 10]等血液生化指标, 和组织中硫胺素的沉积量[10]作为评价水产动物硫胺素缺乏症的一些指标。
近年来, 团头鲂的养殖量已经攀升到我国淡水鱼类养殖的第六名[11, 12], 团头鲂以其肉质鲜嫩深受人们喜爱, 因此随着团头鲂养殖规模的继续扩大, 全价配合饲料的开发刻不容缓。目前, 有关团头鲂基本营养素需求的研究已有部分报道[13], 本实验室前期对团头鲂饲料蛋白、脂肪和糖[14-16]等大量营养素需求量进行了研究, 并考察了团头鲂对常见饲料原料的消化率。关于团头鲂对维生素需求量的研究已有报道的包括生物素[17]、吡哆醇[18]、胆碱[19]等, 目前仍未见关于团头鲂对硫胺素需求量的研究。本试验通过研究饲料中添加不同水平的硫胺素对团头鲂幼鱼生长、血浆生化指标和肝脏硫胺素沉积量的影响, 以得出团头鲂幼鱼的适宜硫胺素需求量, 为开发出团头鲂全价配合饲料提供理论依据, 并为团头鲂幼鱼营养素需求量提供基础数据。
1. 材料与方法
1.1 试验饲料
本试验采用单因素试验设计, 配制6组半纯合饲料, 分别添加硫胺素(Sigma-Aldrich公司)含量为0、0.5、1、1.5、2和2.5 mg/kg。试验饲料中蛋白源为酪蛋白、明胶和鱼粉, 糖源为玉米淀粉, 填充物为微晶纤维素, 以鱼油鲶豆油按照1鲶1添加量为脂肪源配制等氮等能的半纯合饲料(即基础饲料), 其营养成分按照团头鲂幼鱼的适宜需求量进行配制[13, 14]。并以羧甲基纤维素钠为黏结剂, 添加部分水, 将粉碎后的原料逐级混匀, 后用小型制粒机制粒, 将饲料颗粒风干后避光保存于-20℃冰箱冷藏备用。在饲料制作完成后, 测定各试验组饲料的硫胺素实际含量分别为0、0.51、0.98、1.59、2.13和2.68 mg/kg。饲料组成及营养成分见表 1。
表 1 基础饲料配方及营养组成(%以风干样为基础)Table 1. Formulation and proximate composition of the basal diet (% air-dry basis)成分Ingredient 含量Content (%) 营养成分Nutrient 含量Content (%) 鱼粉1Fish meal 10.30 粗蛋白Crude protein 31.02 酪蛋白2Casein 24.00 粗脂肪Ether extract 6.86 明胶3Gelatin 6.00 粗纤维Crude fiber 13.23 玉米淀粉1Corn starch 37.40 粗灰分Ash 8.54 鱼油1Fish oil 3.1 总能Gross energy (MJ/kg) 17.89 豆油1Soybean oil 3.1 纤维素4Cellulose 11.10 磷酸二氢钙5Calcium biphosphate 1.80 羧甲基纤维素6Carboxymethyl cellulose 2.00 预混料(无VB1)5 Premix without VB17 1.20 注:1购自江苏正昌饲料公司(其中鱼粉为进口的秘鲁鱼粉); 2购自甘肃华安生物科技集团; 3购自上海鲁博明胶有限公司; 4购自上海益丰食品添加剂有限公司; 5购自南京华牧动物科技研究所; 6购自国药集团化学试剂有限公司; 7预混料提供以下矿物质(mg/kg)和维生素(IU或者mg/kg): CuSO4·5H2O, 2.0 g; FeSO4·7H2O, 25g; ZnSO4·7H2O, 22 g; MnSO4·4H2O, 7 g; Na2SeO3, 0.04 g; KI, 0.026 g; CoCl2·6H2O, 0.1 g; 维生素A, 900000IU; 维生素D, 200000IU; 维生素E, 4500 mg; 维生素K3, 220 mg; 维生素B2, 1090 mg; 维生素B6, 500 mg; 烟酸, 2000 mg; 维生素B12, 1.6 mg; 维生素C, 5000 mg; 泛酸, 1000 mg; 叶酸, 165 mg; 胆碱, 60000 mg; 肌醇, 15000 mg; 生物素100 mg
Note:1 Provided by Zhengchang Feed Industry Co., Ltd (Jiangsu, China) (The fish meal is bought from Peru); 2 Provided by Huaan Biotechnology Co., Ltd (Gansu, China); 3 Provided by Bolu Casein Co., Ltd (Shanghai, China); 4 Provided by Yifeng Food Additives Co., Ltd (Shanghai, China); 5 Provided by Huamu Animal Science Research Institute (Nanjing, China); 6 Provided by Sinopharm Chemical Reagent Co., Ltd (Beijing, China); 7 Premix supplied the following minerals (g/kg) and vitamins (IU or g/kg): CuSO4·5H2O, 2.0 g; FeSO4·7H2O, 25 g; ZnSO4·7H2O, 22 g; MnSO4·4H2O, 7 g; Na2SeO3, 0.04 g; KI, 0.026 g; CoCl2·6H2O, 0.1 g; Vitamin A, 900000IU; Vitamin D, 200000IU; Vitamin E, 4500 mg; Vitamin K3, 220 mg; Vitamin B2, 1090 mg; Vitamin B6, 500 mg; Niacin, 2000 mg; Vitamin B12, 1.6 mg; Vitamin C, 5000 mg; Pantothenate, 1000 mg; Folic acid, 165 mg; Choline, 60000 mg; Myoinositol 15000 mg; Biotin 100 mg1.2 试验用鱼及饲养管理
试验鱼购自江苏扬州国家级良种场, 饲喂于江苏省淡水水产研究所禄口基地。驯化1周, 期间投喂不含硫胺素的基础日粮。驯化后进行分组, 挑选身体健壮、规格整齐的团头鲂720尾[初重为(0.30±0.01) g], 按随机原则分为6组, 每组4重复, 各重复30尾。试验鱼采用室内流水水族箱(规格为3 m×0.8 m×0.8 m)饲养, 每日投饲3次, 饱食投喂, 30min后观察摄食情况并吸取残饵, 养殖周期为8周。试验期间全程使用经过滤、紫外灭菌的池塘水, 24h充氧, pH为7.2-7.4, 水温为(28±4)℃, 流水速度为2 L/min, 溶氧 > 5.0 mg/L, 氨氮含量 < 0.5 mg/L, 亚硝酸盐 < 0.05 mg/L。
1.3 指标测定与方法
生产性能统计和样本采集 在养殖试验完成后, 将鱼饥饿24h, 统计每个水族箱的尾数并称总重, 从中随机取6尾鱼, 然后采用浓度为100 mg/L间氨基苯甲酸乙酯甲磺酸盐(MS-222, Sigma-Aldrich公司)将鱼麻醉。用一次性注射器从尾静脉采血, 采集的血液置于肝素抗凝管中, 于4℃下3500 r/min离心10min, 收集血浆, 保存于-20℃待测。肝脏用0.85%生理盐水冲洗, 用滤纸吸干, 迅速放入液氮罐中, 后转移至-70℃冷冻保存, 用于测定肝脏硫胺素的含量。
生长性能的计算公式如下:
增重率(Weight gain, WG, %)=(Wt -W0)×100/W0
特定生长率(Specific growth rate, SGR, %/d)=[ln (Wt )-ln (W0)]×100/t
饲料系数(Feed conversion ratio, FCR)=F/(Wt -W0)
式中W0为鱼体初重(g); Wt 为鱼体末重(g); F为摄食量(g); t为饲养天数(d)。
样品测定 饲料中粗蛋白采用全自动凯氏定氮仪测定; 粗脂肪采用索氏抽提法测定; 粗纤维采用纤维分析仪测定; 灰分采用高温灼烧法测定; 总能采用氧弹法测热仪测定。饲料和肝脏中硫胺素的含量采用高压液相色谱法测定[20], 具体步骤如下:称取0.5 g左右饲料和肝脏样品, 加入3 mL 0.01 mol/L盐酸(HCL)后匀浆, 再加入1 mL 10%三氯乙酸沉淀蛋白, 后将混合液离心10min (4℃, 12500×g)。用乙醚将上层清液清洗3遍, 然后将乙醚蒸发除去。经0.21 μm滤膜过滤后, 然后用高压液相色谱系统(HP 1100)检测, 色谱柱为OSD Hypersil柱(5 μm, 4 mm×160 mm)。流动相由25 mmol/L磷酸氢钾和甲醇按85鲶15组成, 流速为1.0 mL/min。检测系统为荧光检测器, 激发光360 nm, 发射光425 nm。每次进样前, 需先加入0.1%铁氰化钾和15%氢氧化钠进行柱前衍生。取适量硫胺素样品溶解在0.01 mol/L盐酸中作为标准样品。
血浆生化指标的测定 血糖、丙酮酸和乳酸, 均采用南京建成生物工程研究所的试剂盒检测。血糖测定采用葡萄糖氧化酶-过氧化物酶法, 丙酮酸测定采用丙酮酸-氧化酶法, 乳酸测定采用LAC酶显色法测定, 具体操作方法参见试剂盒说明书。
1.4 数据统计与分析
试验数据采用采用SPSS 16.0软件分析, 试验结果以平均值±标准误(Mean±SE)表示, 单因素方差分析方法(One-way ANOVA), 并用Duncan氏多重比较法对数据进行显著性检验, 显著性水平设为P < 0.05。
2. 结果
2.1 饲料中硫胺素含量对团头鲂幼鱼生长性能的影响
由表 2可知, 饲料硫胺素含量为0时, 相比添加量为0.98、1.59、2.13和2.68 mg/kg添加组的增重率显著降低(P < 0.05), 当硫胺素含量进一步升高时, 变化不显著(P > 0.05)。饲料硫胺素0添加组的特定生长率显著低于0.98、1.59、2.13和2.68 mg/kg添加组(P < 0.05), 当硫胺素含量继续升高时, 差异不显著(P > 0.05)。饲料硫胺素含量为2.13 mg/kg组的饲料系数最低, 且显著低于0组(P < 0.05), 但和其余组之间差异不显著(P > 0.05)。1.59和2.13 mg/kg硫胺素添加组成活率显著高于0和0.51 mg/kg硫胺素添加组(P < 0.05), 但和其他各组差异不显著(P > 0.05)。
表 2 不同硫胺素含量对团头鲂幼鱼增重和饲料转化率的影响Table 2. Effects of dietary thiamin levels on weight gain and feed conversion ratio of juvenileMegalobrama amblycephala实测值Measured
value (mg/kg)增重率WG (%) 特定生长率SGR (%) 饲料系数FCR 成活率Survival (%) 0 1030.64±87.16b 4.27±0.26b 3.58±0.09a 72.13±3.80b 0.51 1132.22±62.15ab 4.46±0.16ab 3.45±0.23ab 81.72±1.79b 0.98 1236.42±63.17a 4.60±0.16a 3.36±0.23ab 89.81±1.69ab 1.59 1285.03±41.42a 4.68±0.10a 3.19±0.09ab 94.25±1.97a 2.13 1293.05±72.38a 4.67±0.18a 3.15±0.09b 95.33±2.15a 2.68 1284.90±81.63a 4.65±0.20a 3.18±0.09ab 88.75±2.91ab 注:表中同列数据后标注不同小写字母表示差异显著(P < 0.05);下同
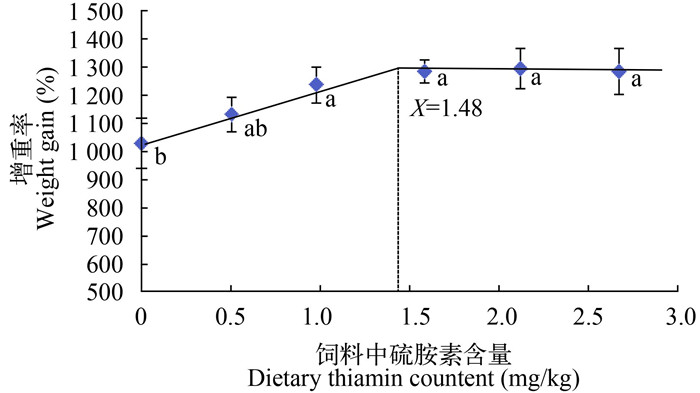
Note: Values with different small letter superscripts in the same column mean significant difference (P < 0.05). The same applies below根据增重率与饲料中硫胺素含量的关系, 以饲料中硫胺素含量为横坐标, 增重率为纵坐标, 进行双折线回归分析得出Y=1287.13-163.6(X-1.48)(R2=0.958), 对其求解, 当饲料硫胺素水平为1.48 mg/kg时, 团头鲂幼鱼增重率最大(图 1)。
![]() 图 1 饲料中硫胺素含量与团头鲂幼鱼增重率之间的关系Figure 1. The relationship between dietary thiamin levels and WG of juvenileMegalobrama amblycephala
图 1 饲料中硫胺素含量与团头鲂幼鱼增重率之间的关系Figure 1. The relationship between dietary thiamin levels and WG of juvenileMegalobrama amblycephala2.2 饲料中硫胺素含量对团头鲂幼鱼肝脏硫胺素沉积量的影响
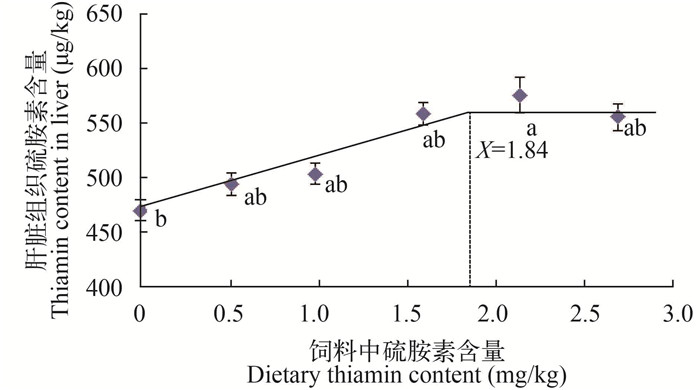
肝脏硫胺素沉积量随着饲料中硫胺素含量的增加而上升, 当硫胺素含量为2.13 mg/kg时达到最大值, 显著高于0添加组(P < 0.05), 相比其余各组差异性不显著(P > 0.05)。以饲料中硫胺素添加量为横坐标, 肝脏硫胺素沉积量为纵坐标, 进行双折线回归分析, 得出Y=563.01-52.87(X-1.84)(R2=0.931), 故团头鲂幼鱼对硫胺素的适宜需求量为1.84 mg/kg (图 2)。
![]() 图 2 饲料中硫胺素与团头鲂幼鱼肝脏硫胺素含量之间的关系Figure 2. The relationship between dietary thiamin levels and liver thiamin contents of juvenileMegalobrama amblycephala
图 2 饲料中硫胺素与团头鲂幼鱼肝脏硫胺素含量之间的关系Figure 2. The relationship between dietary thiamin levels and liver thiamin contents of juvenileMegalobrama amblycephala2.3 饲料中硫胺素添加量对团头鲂幼鱼血浆生化指标的影响
由表 3可知, 随着饲料中硫胺素含量的升高, 血浆葡萄糖水平先下降后上升, 在1.59 mg/kg饲料组时达到最小值, 其次是2.13 mg/kg组, 两组间差异性不显著(P > 0.05), 相比其他各组差异显著(P < 0.05)。0和0.51 mg/kg组血浆中丙酮酸含量较高, 且0组显著高于0.98-2.68 mg/kg各组(P < 0.05), 但0.98-2.68 mg/kg这几组之间差异性不显著(P > 0.05)。血浆中乳酸含量各组间差异性不显著(P > 0.05)。
表 3 不同硫胺素含量对团头鲂幼鱼血浆生化指标的影响Table 3. Effects of dietary thiamin levels on plasma biochemical parameters of blunt snout bream实测值Measured value (mg/kg) 葡萄糖GLU (mmol/L) 丙酮酸Pyruvate (mmol/gprot) 乳酸Lactate (mmol/gprot) 0 4.37±0.55a 0.16±0.01a 5.24±0.85 0.51 4.23±0.21a 0.16±0.00ab 5.80±0.19 0.98 4.15±0.55a 0.15±0.00b 5.78±0.60 1.59 3.92±0.57b 0.15±0.01b 5.59±0.47 2.13 4.07±0.23b 0.15±0.01b 5.93±0.22 2.68 4.15±0.41a 0.15±0.01b 5.44±0.46 3. 讨论
3.1 饲料硫胺素添加量对团头鲂幼鱼生长指标和肝脏沉积量的影响
以增重率为评价指标, 通过折线回归模型分析, 得到团头鲂幼鱼适宜硫胺素需求量为1.48 mg/kg。这与草鱼(1.16-1.19 mg/kg)[9]和大菱鲆(0.6-2.6 mg/kg)[4]的需求量接近, 高于鲤鱼(0.5 mg/kg)[2]、建鲤(1.02 mg/kg)[7]、虹鳟(1.0 mg/kg)[3]等的需求量, 但低于军曹鱼(6.80-8.56 mg/kg)[8]、奥尼罗罗非鱼(3.5 mg/kg)[6]的需求量。有研究表明, 鱼类的硫胺素需求量与饲料中的糖水平有关[21], 当日粮糖水平较高时, 鱼类硫胺素的需求量会相应升高。团头鲂作为淡水草食性鱼类, 对饲料中糖类的利用率较高[15], 饲料糖水平在淡水鱼类中处于较高水平, 这可能是团头鲂幼鱼对硫胺素的需求量在各种鱼中处于较高水平的原因。此外, 这种需求量的差异还可能与鱼的种类、生长阶段、食性、试验日粮的营养水平、养殖条件和评价指标等有关。在本次实验中, 特定生长率和增重率在不同硫胺素添加水平表现出了相同的变化趋势, 硫胺素0添加的对照组生长缓慢, 特定生长率显著低于添加适量硫胺素的实验组, 这和赵智勇[9]的研究相似, 出现这种情况可能是因为对照组缺乏硫胺素导致团头鲂幼鱼体内糖代谢受阻, 不能正常氧化供能和合成脂肪, 加快了体内蛋白的供能消耗, 从而表现出生长缓慢。另外, 适宜水平的硫胺素添加能够降低团头鲂幼鱼的饲料系数, 可能是因为硫胺素使得鱼体能够有效地利用饲料中的糖类, 而糖分解的产物能够使鱼体更好地利用饲料中脂类和蛋白质[9], 从而提升饲料的利用率, 降低饲料系数。
肝脏硫胺素沉积量也通常作为确定水产动物硫胺素需求量的评价指标[9, 10], 本试验结果表明随着饲料中硫胺素添加量的升高, 肝脏组织中的硫胺素沉积量先升高后基本稳定, 试验结果与赵智勇[9]和任春等[10]一致。通过双折线回归分析得出的团头鲂幼鱼对硫胺素需求量为1.84 mg/kg, 高于通过增重率得出的结果, 这与前人在维生素需求量中的研究相似[7, 19], 有研究表明, 鱼类由外界摄取的维生素大部分用在稳定该维生素在体内状态的平衡[22], 而为了达到鱼体内部该种维生素的稳定, 必须能摄取到比鱼类达到最大生长所需要的更多的维生素的浓度才能用于维持内部的稳定[23], 因此利用肝脏硫胺素沉积量来评价鱼类的适宜硫胺素需要量比以增重率为评价的需求量偏大。
3.2 饲料硫胺素含量对团头鲂幼鱼成活率的影响
在本试验中, 从第四周开始, 0硫胺素组试验鱼开始出现厌食, 游动缓慢, 体色加深, 死亡率增加等症状, 这可能揭示硫胺素是团头鲂幼鱼生长所必需的营养物质, 缺乏硫胺素对团头鲂幼鱼摄食、活动、体色均产生严重影响。硫胺素在鱼体内被合成焦磷酸硫胺素, 作为辅酶促进糖类分解和糖的异生作用, 在葡萄糖代谢过程中发挥重要作用, 当饲料中缺乏硫胺素时可引起体内糖代谢受阻, 导致氧化供能和脂肪合成功能异常, 鱼体分解蛋白质供能, 这可能是本试验中缺乏硫胺素组团头鲂幼鱼生长缓慢的原因之一。相关研究表明, 硫胺素缺乏使斑点叉尾鲙[3]、虹鳟[5]和建鲤[7]等出现生长缓慢、厌食、死亡率升高等情况, 和本试验结果相吻合。
3.3 饲料硫胺素添加量对团头鲂幼鱼血液生化的影响
本试验考察了试验鱼血浆葡萄糖、丙酮酸和乳酸水平, 发现缺乏硫胺素组血浆丙酮酸含量较高, 而适宜的硫胺素水平组血浆丙酮酸含量低, 这与Morito等[3]、Lim等[6]和任春等[10]得出的结果一致。本实验可能揭示了硫胺素对丙酮酸的影响, 当饲料中硫胺素不足时, 可能造成团头鲂幼鱼体内丙酮酸氧化脱羧酶的合成量降低, 使得丙酮酸无法正常转化为乙酰辅酶A, 导致三羧酸循环障碍, 糖的有氧氧化异常[24], 故血浆丙酮酸含量升高。在本实验中硫胺素缺乏组团头鲂幼鱼血糖水平显著高于其他组, 这与赵智勇[9]的研究结果相似, 可能原因是硫胺素缺乏造成的三羧酸循环障碍抑制了血糖的有氧氧化过程, 进而导致血糖堆积。
综上所述, 以团头鲂幼鱼的增重率和肝脏硫胺素沉积量为评价指标, 进行双折线回归分析, 饲料中硫胺素适宜添加量分别为1.48和1.84 mg/kg, 添加硫胺素1.59-2.13 mg/kg能够提高团头鲂幼鱼的成活率。
-
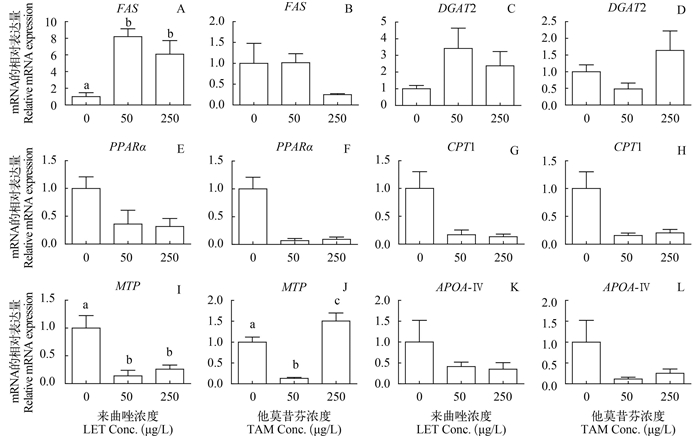
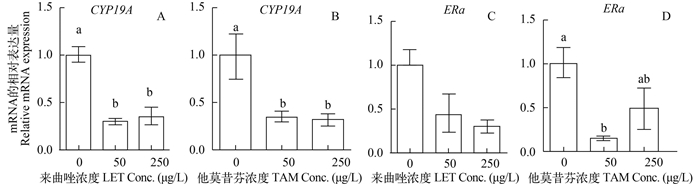
![]()
图 1 LET与TAM对雌性斑马鱼暴露, 肝脏CYP19A与ERα基因相对表达量
图中各组之间标注不同字母表示差异显著, P < 0.05;下同
Figure 1. Effect of LET and TAM on hepatic genes related to estrogen synthesis and estrogen receptor of female zebrafish
Values in groups with different letters are significantly different, P < 0.05; the same applies below
![]()
图 2 LET与TAM对雌性斑马鱼暴露, 肝脏相关脂肪代谢基因的相对表达量
Figure 2. Effect of LET and TAM on hepatic genes related to lipid metabolism of female zebrafish
表 1 用于实时荧光定量PCR分析的引物序列
Table 1 Sequences of real time PCR primers
引物名称
Primer引物序列Sequense (5′-3′) 引物来源
Source of primerβ-actin F: TCTGGTGATGGTGTGACCCA 刘晓丽等[18] R: GGTGAAGCTGTAGCCACGCT CYP19A F: CTGAAAGGGCTCAGGACAA Shi, 等[19] R: TGGTCGATGGTGTCTGATG ERα F: TCAGCGGTTCCAGTCTTC 刘晓丽等[18] R: CTGCTGGTTGTGGGTGTA FAS F: GGAGCAGGCTGCCTCTGTGC 吴俊琳[20] R: TTGCGGCCTGTCCCACTCCT DGAT2 F: ACGCATAACCTGCTTCCC 本研究 R: TCCTGTGGCTTCTGTCCC PPARα F: CTGCGGGACATCTCTCAGTC 吴俊琳[20] R: ACCGTAAACACCTGACGACG CPT1 F: GCATTGACCTTCAGCTCAGC 吴俊琳[20] R:CTGCCAACACCAGCACGAAC MTP F: AAGATCAGAAGCCTCTATGC 本研究 R: TCAACCAGACATTTCCCT APOA-Ⅳ F:AGCTGCACAATCCTCGTCTGC GM Her, 等[21] R:TGCAGGAGCCTGTGTTTGCCT  下载: 导出CSV
下载: 导出CSV
表 2 斑马鱼肝脏、内脏、肌肉中的脂肪含量
Table 2 Effects of LET and TAM on the triglyceride content in liver, viscera and muscle of female zebrafish (%)
组别
Group肝脏脂肪含量
Lipids of liver内脏脂肪含量
Lipids of
visceral肌肉脂肪含量
Lipids of
muscle对照组 3.45±0.49a 7.08±1.03a 4.26±0.75 低浓度LET 4.61±0.33b 11.3±2.64b 5.33±0.99 高浓度LET 3.75±0.75a 6.84±1.96a 4.52±0.85 低浓度TAM 3.69±0.73a 8.14±1.11a 3.78±1.29 高浓度TAM 1.52±0.04c 5.00±1.61a 3.45±1.02 注:数据表示为平均值±标准差, 同一列标注不同字母表示差异显著, P < 0.05
Note: Valuesare the mean±standard errors. Values in same row with different letters are significantly different, P < 0.05
下载: 导出CSV
-
[1] 杜震宇.养殖鱼类脂肪肝成因及相关思考.水产学报, 2014, 38(9):1628-1638 http://www.cnki.com.cn/Article/CJFDTOTAL-SCKX201409053.htm DU Z Y. Causes of fatty liver in farmed fish:a review and new perspectives[J]. Journal of Fisheries of China, 2014, 38(9):1628-1638
杜震宇.养殖鱼类脂肪肝成因及相关思考.水产学报, 2014, 38(9):1628-1638 http://www.cnki.com.cn/Article/CJFDTOTAL-SCKX201409053.htm[2] Pierron F, Baudrimont M, Bossy A, et al. Impairment of lipid storage by cadmium in the European eel (Anguilla anguilla)[J]. Aquatic Toxicology, 2007, 81(3):304-311 doi: 10.1016/j.aquatox.2006.12.014
[3] Riu A, Mccollum C W, Pinto C L, et al. Halogenated bisphenol-A analogs act as obesogens in zebrafish larvae (Danio rerio)[J]. Toxicological Sciences an Official Journal of the Society of Toxicology, 2014, 139(1):48-58 doi: 10.1093/toxsci/kfu036
[4] Ley C J, Lees B, Stevenson J C. Sex-and menopause-associated changes in body-fat distribution[J]. American Journal of Clinical Nutrition, 1992, 55(5):950-954
[5] Lemieux S, Prud'Homme D, Bouchard C, et al. Sex differences in the relation of visceral adipose tissue accumulation to total body fatness[J]. American Journal of Clini-cal Nutrition, 1993, 58(4):463-467 https://www.researchgate.net/profile/Simone_Lemieux/publication/14818808_Sex_differences_in_relation_of_visceral_adipose_tissue_accumulation_to_total_body_fatness/links/54b92d810cf24e50e93c2d61.pdf
[6] Gambacciani M, Ciaponi M, Cappagli B, et al. Body weight, body fat distribution, and hormonal replacement therapy in early postmenopausal women[J]. Journal of Clinical Endocrinology & Metabolism, 1997, 82(2):414-417 http://d.scholar.cnki.net/detail/SJPD2059_U/SJPD12102900978773
[7] Haarbo J, Marslew U, Gotfredsen A, et al. Postmenopau-sal hormone replacement therapy prevents central distribution of body fat after menopause[J]. Metabolism-clini-cal & Experimental, 1991, 40(12):1323-1326 https://www.researchgate.net/publication/21195135_Postmenopausal_hormone_replacement_therapy_prevent_central_distribution_of_body_fat_after_menopause
[8] Jones M E, Thorburn A W, Britt K L, et al. Aromatase-deficient (ArKO) mice have a phenotype of increased adiposity[J]. Proceedings of the National Academy of Sciences of the United States of America, 2000, 97(23):12735-12740 doi: 10.1073/pnas.97.23.12735
[9] Jones M E E, Thorburn A W, Britt K L, et al. Aromatase-deficient (ArKO) mice accumulate excess adipose tissue[J]. Journal of Steroid Biochemistry & Molecular Biology, 2001, 79(s1-5):3-9 https://www.researchgate.net/publication/11511588_Aromatase-deficient_ArKO_mice_accumulate_excess_adipose_tissue
[10] Heine P A, Taylor J A, Iwamoto G A, et al. Increased adipose tissue in male and female estrogen receptor-alpha knockout mice[J]. Proceedings of the National Academy of Sciences of the United States of America, 2000, 97(23):12729-12734 doi: 10.1073/pnas.97.23.12729
[11] Ohlsson C, Hellberg N, Parini P, et al. Obesity and disturbed lipoprotein profile in estrogen receptor-α-deficient male mice[J]. Biochemical & Biophysical Research Communications, 2000, 278(3):640-645 https://www.researchgate.net/profile/Bo_Angelin/publication/12231872_Obesity_and_Disturbed_Lipoprotein_Profile_in_Estrogen_Receptor-a-Deficient_Male_Mice/links/57d8077208ae0c0081ede8ba.pdf?origin=publication_detail
[12] Sun L, Zha J, Spear P A, et al. Tamoxifen effects on the early life stages and reproduction of Japanese medaka (Oryzias latipes)[J]. Environmental Toxicology & Pharmacology, 2007, 24(1):23-29 https://www.researchgate.net/profile/Zijian_Wang3/publication/51516110_Tamoxifen_effects_on_the_early_life_stages_and_reproduction_of_Japanese_medaka_(Oryzias_latipes)/links/55b1cb5e08ae092e964ff5eb.pdf?inViewer=true&pdfJsDownload=true&disableCoverPage=true&origin=publication_detail
[13] Sun L, Zha J, Spear P A, et al. Toxicity of the aromatase inhibitor letrozole to Japanese medaka (Oryzias latipes) eggs, larvae and breeding adults[J]. Comparative Biochemistry & Physiology Part C Toxicology & Pharmacology, 2007, 145(4):533-541 https://www.researchgate.net/publication/6423480_Toxicity_of_the_aromatase_inhibitor_letrozole_to_Japanese_medaka_Oryzias_latipes_eggs_larvae_and_breeding_adults
[14] Sun L, Shao X, Jian C, et al. Transcriptional responses in the brain, liver and gonad of Japanese ricefish (Oryzias latipes) exposed to two anti-estrogens[J]. Comparative Biochemistry & Physiology Part C Toxicology & Pharmacology, 2011, 153(4):392-401 https://www.researchgate.net/publication/49798840_Transcriptional_responsein_the_brain_liver_and_gonad_of_Japanese_ricefish_Oryzia_latipes_exposedto_two_anti-estrogens?_sg=2oG3FzYmMZ22wJFoeBoRequn1tI3EzWCpQnbOyZoOuR-24PwPTuAAooM4zpmUFul-G533AaIt6Uu0TTzQxHt7kGIHHkxJ8YIhE2Zh_yZA0Y
[15] Percec V, Clough R S, Grigoras M, et al. Reductive dehalogenation versus substitution in the polyetherification of 4, 4'-dihalodiphenyl sulfones with bisphenolates[J]. Macromolecules, 1993, 26(14):3650-3662 doi: 10.1021/ma00066a026
[16] Schrader T J, Langlois I, Soper K, et al. Mutagenicity of bisphenol A (4, 4'-isopropylidenediphenol) in vitro:effects of nitrosylation[J]. Teratogenesis Carcinogenesis & Mutagenesis, 2002, 22(6):425-441 https://www.researchgate.net/publication/11068480_Mutagenicity_of_bisphenol_A_44%27-isopropylidenediphenol_in_vitro_Effects_of_nitrosylation?_sg=dG7KGUEkJW9wf98gZ7r9dCm2qy-8TTCp-dxa78C4u3pT98Uf8T-SzOl_9l9K3zqeFuCkxYNMgdJGrUAnN6oXsA
[17] Luo L F, Yang L Q, Wu D S, et al. Effects of bisphenol A on OCT4 and SOX2 genes expression in mouse embryo-nic stem cells[J]. Chinese Journal of Preventive Medicine, 2013, 47(2):164-169
[18] 刘晓丽, 汪奇, 贾林芝, 等.壬基酚对斑马鱼精巢组织及性激素合成酶基因表达的影响.环境科学学报, 2011, 31(11):2523-2529. http://www.cnki.com.cn/Article/CJFDTOTAL-HJXX201111030.htm Liu X L, Wang Q, Jia L Z, et al. Alterations in testicular histology and the mRNAs of enzymes responsible for sex steroid synthesis in the zebrafish (Danio rerio) exposed to nonyphenol[J]. Acta Scientiae Circumstantiae, 2011, 31(11):2523-2529
刘晓丽, 汪奇, 贾林芝, 等.壬基酚对斑马鱼精巢组织及性激素合成酶基因表达的影响.环境科学学报, 2011, 31(11):2523-2529. http://www.cnki.com.cn/Article/CJFDTOTAL-HJXX201111030.htm[19] Shi X, Du Y, Lam P K S, et al. Developmental toxicity and alteration of gene expression in zebrafish embryos exposed to PFOS[J]. Toxicology & Applied Pharmacology, 2008, 230(1):23-32 https://www.researchgate.net/publication/5445994_Developmental_toxicity_and_alteration_of_gene_expression_in_zebrafish_embryos_exposed_to_PFOS
[20] 吴俊琳.鱼类脂肪组织核磁共振成像研究及斑马鱼(Danio rerio)脂代谢中的性别差异.硕士学位论文.华东师范大学, 上海. 2015 Wu J L. Evaluation of the distribution of fatty tissues in fish using magnetic resonance imaging (MRI) and effects of gender on lipid metabolism in zebrafish, Danio rerio[D]. Thesis for Master of Science. East China Normal University. 2015
吴俊琳.鱼类脂肪组织核磁共振成像研究及斑马鱼(Danio rerio)脂代谢中的性别差异.硕士学位论文.华东师范大学, 上海. 2015[21] Her G M, Hsu C C, Hong J R, et al. Overexpression of gankyrin induces liver steatosis in zebrafish (Danio rerio)[J]. Biochimica Et Biophysica Acta, 2011, 1811(9):536-548 doi: 10.1016/j.bbalip.2011.06.011
[22] Cowley S M, Hoare S, Mosselman S, et al. Estrogen receptors α and β form heterodimers on DNA[J]. Journal of Biological Chemistry, 1997, 272(32):19858-19862 doi: 10.1074/jbc.272.32.19858
[23] Das S K, Taylor J A, Korach K S, et al. Estrogenic responses in estrogen receptor-alpha deficient mice reveal a distinct estrogen signaling pathway[J]. Proceedings of the National Academy of Sciences, 1997, 94(24):12786-12791 doi: 10.1073/pnas.94.24.12786
[24] Paul C. Begovac †, Wallace R A. Stages of oocyte deve-lopment in the pipefish, Syngnathus scovelli[J]. Journal of Morphology, 2005, 197(3):353-369 https://www.researchgate.net/publication/229730983_Stages_of_oocyte_development_in_the_pipefish_Syngnathus_scovelli
-
期刊类型引用(1)
1. 徐超,刘文斌,李远友,谢帝芝,丁志荣,李向飞. 苯磷硫胺对饲喂高糖饲料团头鲂血浆生化指标及肝糖脂代谢的影响. 中国水产科学. 2021(02): 186-194 .  百度学术
百度学术
其他类型引用(3)
计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 4