
EFFECTS OF DIETARY VITAMIN A ON GROWTH PERFORMANCE AND CALCIUM AND PHOSPHORUS CONTENTS IN BONE OF SUBADULT GRASS CARP (CTENOPHARYNGODON IDELLUS)
-
摘要: 实验旨在探究养成期草鱼对饲料中维生素A的需求及维生素A对养成期草鱼的骨骼中钙磷含量的影响。选取养成期(254.6±0.5)g草鱼420尾,随机分为7组,每组3个重复,每个重复20尾。分别用维生素A含量为89、732、1129、2378、4688、7218和9802 IU/kg的等氮等能实验饲料喂养84d。实验结果表明:(1)饲料中添加4688 IU/kg的维生素A显著提高了养成期草鱼的特定生长率(P < 0.05),并显著降低了饲料系数(P < 0.05),但饲料维生素A含量对鱼体摄食率、成活率以及肝脏指数没有显著影响(P>0.05)。(2)饲料添加维生素A对鱼体粗蛋白、粗脂肪、灰分没有显著影响(P>0.05),但是9802 IU/kg组的鱼体水分显著低于其他实验组(P < 0.05)。(3)饲料中4688 IU/kg的维生素A显著提高血清中超氧化物歧化酶的活性(P < 0.05),但饲料维生素A水平对血清丙二醛、钙、磷含量以及碱性磷酸酶活性没有显著影响(P>0.05)。(4)饲料添加4688和7218 IU/kg的维生素A能显著提高草鱼头骨和脊椎骨中灰分以及钙、磷含量(P < 0.05)。在实验条件下,以特定生长率、饲料系数以及血清SOD活性为评价指标,通过折线分析得出养成期草鱼适宜维生素A的需求量分别为3184和3077 IU/kg。Abstract: To determine the optimum dietary requirement of vitamin A (VA) of subadult grass carp (Ctenopharyngodon idellus), seven isoenergetic and isonitrogenous semi-purified diets were formulated to contain VA level at 89, 732, 1129, 2378, 4688, 7218 and 9802 IU/kg diet respectively, for a 12-week growth trial that was conducted in 21 net-cages located in an earth pond. The results indicated that dietary VA level significantly regulated the specific growth rate (SGR) and feed conversion ratio (FCR) of subadult grass carp (P < 0.05) with the highest SGR and the lowest FCR both in 4688 IU/kg group. Feeding rate, survival rate and hepatosomatic index were not affected by dietary VA level (P>0.05). Dietary VA level showed significant (P < 0.05) effect on serum SOD activity with the highest SOD values in 4688 IU/kg group. Serum ALP activity, MDA, calcium (Ca) and phosphorus (P) contents were not significantly (P>0.05) affected by dietary VA level. Body moisture content in 9802 IU/kg group was significantly lower. Body lipid, protein and ash contents were not affected by dietary VA level (P>0.05). The ash contents either in skull or in vertebra were significantly affected (P < 0.05) by dietary VA level. The higher Ca and P contents in skull and vertebra were both observed in 4688 and 7218 IU/kg groups (P < 0.05). Broken-line analysis for the SGR of fish suggested that the optimal dietary VA content for subadult grass carp was 3184 IU/kg diet.
-
Keywords:
- Subadult Grass carp /
- Vitamin A /
- Growth performance /
- Biochemical analysis /
- Bone homeostasis
-
纤毛虫原生动物具有二型核, 小核基因通常被内部删除序列(Internal eliminated sequences, IESs) 分隔成多个大核命运序列(Macronucleus destined sequences, MDSs) 片段。在接合生殖过程中, 通过小核IESs的删除和MDSs的重组连接、端粒序列的生成和基因的高度扩增等过程形成具功能的大核基因[1-5]。染色体重排广泛存在于3种纤毛虫类群中: Armophorea、旋唇纲Spirotrichea、层咽纲Phyllo-pharyngea[6], 许多小核基因MDSs的排列顺序会发生变化, 形成乱序基因。其中在旋唇纲腹毛类纤毛虫中该现象尤为明显, 例如尖毛虫(Oxytricha) 和棘尾虫(Stylonychia) 中分别有96%和98%的小核基因组序列在小核发育成大核的过程中被删除[7]。
在腹毛类纤毛虫中, 有3种乱序基因为人们熟知, 分别为肌动蛋白Ⅰ型(Actin Ⅰ)[1]、端粒结合蛋白(α-TBP)[8]及DNA聚合酶α (DNA polymerase α)[9]。目前已报道10个物种12个种群的小核Actin Ⅰ基因序列[4, 1, 10-14]、6个物种的小核α-TBP基因[8, 15, 16]、7个物种的小核DNApol α基因[9, 17-20]。Hogan等[12]推测了腹毛类纤毛虫小核Actin Ⅰ基因的进化路线, 从共同的原始祖先到3个MDSs的Urostyla grandis, 进化为拥有4个MDSs的Engelmanniella mobilis, MDS2发生倒置, 再进化为拥有8个MDSs的Stylony-chia lemnae, 然后分化为两个分支, 一支为3个尖毛虫类群, 一支为S. pustulata和Oxytricha sp. (Misty)。之后, Chen等[21]在成功扩增2种尾柱目纤毛虫大小核Actin Ⅰ基因的基础上, 将腹毛类小核Actin Ⅰ进化路线进一步修正, 提出腹毛类不同目之间可能存在不同的进化路线。Wong等[16]推测了α-TBP基因可能的进化路线, 认为祖先类群为非乱序物种(如Holosticha sp.), 历经乱序排列的Uroleptus sp.、Pa-raurostyla weissei, 进化为中间MDS发生易位的O. trifallax, 再经历两个MDSs发生融合(如O. nova/S. myti-lus)。DNApol α的进化路线尚未有人给出。综上, 已报道的腹毛类3种乱序基因进化路线仅为较为粗略的大致框架, 细节部分依然亟待修订和补充。
本研究在湛江湖光红树林水域中采集得到一个腹毛类尖毛虫物种Oxytricha sp. (ZJ)。通过测定O. sp. (ZJ) 大、小核的Actin Ⅰ、α-TBP和DNApol α基因序列, 并对尖毛虫属内现有物种3个乱序的小核基因结构进行比较, 探讨尖毛虫属基因乱序模式, 增加人们对相关基因进化的认识。
1. 材料与方法
1.1 样品与DNA提取
Oxytricha sp. (ZJ) 于2010年11月采自湛江湖光红树林水域(东经109°40′-110°35′, 北纬20°14′-21°35′), 镜检后分离[22]。于5‰人工海水中投入米粒滋生细菌, 进行单克隆培养。至大量繁殖阶段, 筛绢过滤除去大的杂质后, 5000 r/min, 3min离心富集虫体。DNA提取按照伊珍珍等[23]改良的Qiagen Dneasy Blood & Tissue Kit试剂盒使用方法操作。电泳检测完整性, 微量核酸测量仪检测其浓度。
1.2 PCR扩增与测定
大核Actin Ⅰ基因序列的获得 利用引物Actin800F和Actin800R1[18](表 1) 获得约800 bp的大核Actin Ⅰ基因片段。PCR反应总体系为50 μL, 包括去离子水36 μL, 10×Ex buffer 5 μL, dNTP mix (2.5 mmol/L) 5 μL, 模板2 μL, 正向反向引物(25 mmol/L) 各0.8 μL, Ex Taq (5 U/μL) 0.4 μL。PCR扩增程序如下: 95℃预变性5min; 95℃变性30s, 52℃退火30s, 72℃延伸1min, 扩增35个循环; 72℃终延伸10min。经1.0%琼脂糖凝胶电泳检测后, 切胶, 用Universal DNA Purification Kit试剂盒回收纯化, 购自天根生化科技(北京) 有限公司, 连接至pMD18-T载体(TaKaRa, 日本), 转化大肠杆菌DH5α感受态细胞, 挑选5个阳性克隆后, 送美吉公司双向测序, 测序引物为M13F-47与M13R-48。
表 1 大、小核Actin Ⅰ基因所用引物名称及序列Table 1. The name and sequence of primers for macro-and micronuclear Actin Ⅰ gene引物名称Name 引物序列Sequence (5′-3′) Actin800F AACTGGGAYGAYATGGARAAGAT Actin800R1 ATCCACATKSHGGCGAAGGT J-F (1203) GGTTATTGCCAGCCCAGACAGAA J-M3F1 TTTCTCAGGTGAGGATGCTCC J-M3F2 TCCCATCAATCGTCGGTAGACCC AP12 GTAATACGACTCTATAGGGCACGCGTGGTCGACGGCCCGGGCTGGTCCCCAAAACCCCAAAACCCCAAAA AP1 GTAATACGACTCACTATAGGGC AP2 ACTATAGGGCACGCGTGGT J-F (673) TGTGATGCTGGTGATGGTGTTAC J-(133RC) CCCTACTAATCTCATTCTTGGCAT J-M9F TGCCCTATCAAAGTATTTCCTA J-F (42) CCATCAGTAAATTTGACATTACTAT J-M8R CTCATTCAACCTGTCCGTAAC J-I1F CATTATGGATTTATGTTTATTGGTG J-I2F CCCAAGTCAATCAACAAAAGTATC J-R (1550) TTGAAACTAATCTGAATAAAAATG 基于上述获得的序列设计引物J-F (1203)、J-M3F1、J-M3F2 (表 1)。利用Chang等[19]建立的端粒抑制PCR (TSP-PCR) 扩增。分别以AP12 (表 1)/Actin800R1、Actin800F/AP12、J-M3F1/AP12为上下游引物进行第一轮PCR, 扩增条件为: 94℃ 2min, 72℃ 4min, 重复7个循环, 再94℃ 2min, 67℃ 4min, 重复32个循环, 然后67℃ 4min。再分别以AP1 (表 1)/Actin800R1、Actin800F/AP1、J-M3F1/AP1为上下游引物做第二轮巢式PCR, AP2 (表 1)/Actin800R1、J-F (1203)/AP2、J-M3F2/AP2为上下游引物做第三轮巢式PCR。扩增条件为: 94℃ 25s, 72℃ 4min, 重复5个循环, 再94℃ 25s, 67℃ 4min, 重复20个循环, 然后67℃ 4min。连续3轮PCR, 获得基因左端序列片段AP2/Actin800R1和右端序列J-F (1203)/AP2以及J-M3F2/AP2。扩增产物经切胶回收、转化、克隆测序, 拼接得到大核基因序列全长, 并提交至GenBank, 登录号为KX602204。
小核Actin Ⅰ基因序列的获得 根据已报道Oxytricha属的小核Actin Ⅰ基因乱序结构, 在推测出的乱序区MDS3/4和MDS1上分别设计上、下游引物, J-F (673)(表 2) 和J-(133RC)(表 2), 使用巢式PCR和降落PCR扩增(95℃预变性5min, 95℃变性30s, 67℃退火30s, 之后退火温度依次降低1℃, 72℃延伸2min, 扩增15个循环; 95℃变性30s, 52℃ 30s, 72℃延伸2min, 扩增30个循环; 72℃终延伸10min)。得到目的条带。切胶回收、转化、克隆测序, 获得小核部分基因片段。
表 2 大、小核α-TBP基因所用引物名称及序列Table 2. The name and sequence of primers for macro-and micronuclear α-TBP gene引物名称Name 引物序列Sequence (5′-3′) αTBP-R GTGTCCTTRATGARRTAGAAWCC J-R2 TTGAAGAAGTTAGCACCGAGACCGT J-R1 GTCGGAGGGTTCAATCTTGGTTACG J-M5F1 TCTACAACAGTTCATGGGCTCTC J-M5F2 CACTTGAGGAAATCAGTGGTGCTG J-M2R TGAGTCACCAGTTCCCTTTTAGC J-M1F1 TTGAGATATGTGGCTGGATTTG J-M1F2 GGCTGGATTTGAAAATACGAGTAG J-I4R GTAGGTAAATTAAAGATTTGGC J-M9F TTAAGGATGGCTCAGGTCAAGT J-M8F1 TTGTTGAGCCCACTTTCTGATG J-M8F2 CGAAAACTTGGAGGATCTTAGCA J-M11F GGTGAGGTAGTAAGAATCAGATC J-M10R GTGGGAATTTAAGTTTAAGAGC J-I9F GAGAAGTCTATAAGTAAAAGTGCC J-R (2133) GATTATAAAATGATTAAAGATCATG J-I13R2 CTTATCATAAACATTATTCAC 在已获得的小核基因序列的基础上设计多条引物J-M9F、J-F (42)、J-M8R、J-I1F、J-I2F、J-R (1550)(表 1)。再分别以J-M9F/J-M8R、J-M3F1/J-(133RC)、J-I1F/J-M8R为上下游引物进行首轮PCR, J-F (42)/J-M8R、J-M3F2/J-R (1550)、J-I2F/J-M8R进行巢式PCR, 扩增得到片段J-F (42)/J-M8R、J-M3F2/J-R (1550)、J-I2F/J-M8R。覆盖小核基因全长。拼接获得小核基因序列全长(图 1), 并提交至GenBank, 登录号为KX602205。
![]() 图 1 Oxytricha sp. (ZJ) 大、小核Actin Ⅰ基因结构、引物位置及获得的PCR产物示意图灰色方块代表MDSs, 黑色方块代表IESs; 其中MDS2倒置; 白色方块代表未得到的基因区域, 下同Figure 1. Schematic illustration of macro-and micronuclear structures, PCR primer locations and products obtained in Oxytricha sp. (ZJ)Actin Ⅰ geneMDSs are represented in grey, IESs are represented in black; MDS2 is inverted; white box represents unavailable gene regim; the same applies below
图 1 Oxytricha sp. (ZJ) 大、小核Actin Ⅰ基因结构、引物位置及获得的PCR产物示意图灰色方块代表MDSs, 黑色方块代表IESs; 其中MDS2倒置; 白色方块代表未得到的基因区域, 下同Figure 1. Schematic illustration of macro-and micronuclear structures, PCR primer locations and products obtained in Oxytricha sp. (ZJ)Actin Ⅰ geneMDSs are represented in grey, IESs are represented in black; MDS2 is inverted; white box represents unavailable gene regim; the same applies below大、小核α-TBP基因序列的获得 根据上文描述的方法测定Oxytricha sp. (ZJ) 的α-TBP基因的完整大核序列。大核扩增所用引物为αTBP-F、αTBP-R、J-R2、J-R1、J-M5F1、J-M5F2 (表 2)。小核所用引物为J-M2R、J-M1F1、J-M1F2、J-I4R、J-M9F、J-M8F1、J-M8F2、J-M11F、J-M10R、J-I9F、J-R (2133)、J-I13R2 (表 2、图 2), 并提交至Gen-Bank, 大核基因登录号为KX602208, 小核基因为KX602209。
![]() 图 2 Oxytricha sp. (ZJ) 大、小核α-TBP基因结构、引物位置及获得的PCR产物示意图Figure 2. Schematic illustration of macro-and micronuclear structures, PCR primer locations and products obtained in Oxytricha sp. (ZJ)α-TBP gene
图 2 Oxytricha sp. (ZJ) 大、小核α-TBP基因结构、引物位置及获得的PCR产物示意图Figure 2. Schematic illustration of macro-and micronuclear structures, PCR primer locations and products obtained in Oxytricha sp. (ZJ)α-TBP gene大、小核DNApol α基因序列的获得 基于上文描述的方式获得Oxytricha sp. (ZJ) 的DNApol α基因完整的大核序列。引物为Pol α-F、Pol α-R、J-R2 (DPα)、J-R1 (DPα)、J-F3 (DPα)、J-F4 (DPα) (表 3), 得到DNApol α基因完整的大核序列。
表 3 大、小核DNApol α基因所用引物名称及序列Table 3. The name and sequence of primers for macro-and micronuclear DNApol α gene引物名称Name 引物序列Sequence (5′-3′) Pol α-F CTAYTGGATHGATGCWCATG Pol α-R CACCAVTCTCTWCKNACCATATC J-R2(DPα) CTAGTAACTGGCTTGCATCTCC J-R1(DPα) AGTGCCTCTAGCTTTGCCTTTC J-F3(DPα) CGATGGAGTGTTTAAGAGTTTAC J-F4(DPα) ATCACCAGATGCTAAAGTTGTGC J-M32F CGTTTCTGAACTAAATCTCT J-M1R1 CCTCTCTTCCTGACTATGTTGTA J-M1R2 TTTAGTTCCATCTTTACGCCCAG J-L1F TCCACTCATCATTTTTTTACTGC J-R5R CTAGTAACTGGCTTGCATCTCC J-R6R AATGTGTTTCCAGTGACWGTGCT J-R9R TCTTTTCTCTGTCTGAGTTGCTA J-5000R CATGCTCACTCTCTTAATACCTC J-5100R GCTAAACCCCATGTAGAAAAGTTG 因在DNApol α基因小核序列扩增时, 扩增结果均为大、小核基因序列的嵌合体, 故只获得部分序列。所用引物为J-M32F、J-M1R1、J-M1R2。J-L1F、J-R5R、J-R6R、J-R9R、J-5000R、J-5100R (表 3、图 3), 并提交至GenBank, 大核基因登录号为KX602206, 小核基因为KX602207。
![]() 图 3 Oxytricha sp. (ZJ) 大、小核DNApol α基因结构、引物位置及获得的PCR产物示意图Figure 3. Schematic illustration of macro-and micronuclear structures, PCR primer locations and products obtained in Oxytricha sp. (ZJ) DNApol α gene
图 3 Oxytricha sp. (ZJ) 大、小核DNApol α基因结构、引物位置及获得的PCR产物示意图Figure 3. Schematic illustration of macro-and micronuclear structures, PCR primer locations and products obtained in Oxytricha sp. (ZJ) DNApol α gene大核Actin Ⅰ基因系统树的构建 因为已完成大核α-TBP基因和DNApol α基因测序的尖毛虫物种数目太少(仅3种), 故而只依据Actin Ⅰ基因序列构建系统树。除本工作测得序列(序列号为: KX-602204) 外, 另外7条序列源于Genbank数据库(序列号为: U63567、AF508049、AF508050、AY044839、U18940、U63566、M22480)。使用Bioedit7.1.9.0[24]内置Clustal进行多序列比对, 序列末端截平, 并手动调整删除空格或比对模糊序列。本工作使用Mr-Modeltest 2[25]进行核酸模型筛选GTR+G+I, 使用MrBayes3.1.2[26]构建了贝叶斯树(Bayesian infe-rence, BI)。用马尔科夫链的蒙特卡洛方法(MCMC) 设置为4条链, 运行1000000代, 每100代进行抽样, 以后验概率来表示各分支的可信性。TreeView v5.1.2[27]、MEGA7[28]等软件用于树形图的美化。
2. 结果
2.1 Oxytricha sp. (ZJ) 的大、小核Actin Ⅰ基因结构特征
Oxytricha sp. (ZJ) 的大核Actin Ⅰ基因全长为1546 bp (含端粒序列)。含有编码375个氨基酸的开放阅读框(ORF), 其AT含量为53.81%。5′和3′非编码区的序列长度分别为198 bp (含20 bp端粒序列) 和226 bp (含20 bp端粒序列), AT含量分别为73.71%和62.87%。5个阳性菌斑测序累计检测到59个碱基变异位点, 其中ORF含有38个碱基变异位点, 涉及12个氨基酸的差异, 未引起移码突变或无义突变。剩余的碱基变异位点, 有5个位于5′非编码区, 16个位于3′非编码区。推测多克隆间序列差异是由于基因重复现象引起[29]。
Oxytricha sp. (ZJ) 的小核Actin Ⅰ基因序列长度为1775 bp。通过与大核序列比对确定该小核基因由8个IESs和9个MDSs组成。IESs序列长度为11-79 bp, MDSs序列长度为19-583 bp。MDS/IES交接处有5-13 bp的指针重复序列。8个IESs均为富含AT的序列(68.00%-100.00%)(表 4)。MDSs按照3-4-6-5-7-9-2-1-8的顺序排列, 其中MDS2为倒置排列(图 1)。
表 4 Oxytricha sp. (ZJ) 小核Actin Ⅰ基因的MDSs、IESs及指针序列Table 4. Characteristics of MDSs, IESs, and pointer sequences in Oxytricha sp. (ZJ)Actin Ⅰ geneMDSn MDSn的长度Length of MDSn MDSn和(n+1) 间的指针序列Pointer sequence between MDSn and (n+1) MDSi和MDSj间IES的长度Length of IESs between MDSi and MDSj IESs AT含量IESs AT content (%) 1 196 ACAACAAACTTGC 3-4 79 79.75 2 35 ? …GGCTGG 4-6 30 73.33 3 338 TCTCT 6-5 68 83.82 4 583 CCAAGTCAATCAA 5-7 30 80.00 5 19 TATTRCCAGC 7-9 11 100.00 6 139 CTGAGCAATCA 9-2 21 90.48 7 87 TTAACWATTTR 2-1 26 80.77 8 ? AGGTGG…? 1-8 25 68.00 9 99 6-8 注: MDSs和IESs的长度都包括了一拷贝的指针序列; 不同克隆间指针序列的差异用兼并碱基表示; 问号代表数值或序列无法确定
Note: Length of MDSs include pointers on both ends; IESs exclude pointer sequences; Use degenerate symbols reflects differences between clones; Question marks indicate undetermined information基于尖毛虫属内8个物种的大核Actin Ⅰ基因序列建的贝叶斯树(图 4a) 中, Oxytricha trifallax、O. fallax与O. granulifera以最高置信值聚为一支, 其余5个物种O. longa、O. sp. (Misty)、O. nova、O. sp. (Aspen)、O. sp. (ZJ) 聚为一支。
![]() 图 4 基于Actin Ⅰ基因序列构建的贝叶斯树(a) 与Actin Ⅰ基因结构进化路线图(b)Figure 4. Baysian trees based on the sequences of Actin Ⅰ gene (a), and evolutionary route map of Actin Ⅰ gene structure (b)
图 4 基于Actin Ⅰ基因序列构建的贝叶斯树(a) 与Actin Ⅰ基因结构进化路线图(b)Figure 4. Baysian trees based on the sequences of Actin Ⅰ gene (a), and evolutionary route map of Actin Ⅰ gene structure (b)2.2 Oxytricha sp. (ZJ) 的大、小核α-TBP基因结构特征
Oxytricha sp. (ZJ) 的大核α-TBP基因全长为2140 bp。含有编码495个氨基酸的ORF, 其AT含量为53.56%。5′和3′非编码区的序列长度分别为170 bp (含20 bp端粒序列) 和438 bp (含20 bp端粒序列), AT含量分别为74.83%和66.50%。通过与前人数据[22]比较, 找到一个长度为53 bp、富含AT (77.36%) 的内含子序列。5个阳性克隆测序累计检测到107个碱基变异位点, 其中ORF含有49个碱基变异位点, 涉及19个氨基酸的差异, 未引起移码突变或无义突变。剩余的碱基变异位点, 有2个位于5′非编码区, 53个位于3′非编码区, 3个位于内含子。
Oxytricha sp. (ZJ) 小核α-TBP基因序列长度为2562 bp。通过与大核序列比对确定该小核基因由13个IESs和14个MDSs组成。IESs序列长度为16-71 bp, MDSs序列长度为13-731 bp。在MDS/IES交接处有5-11 bp的指针重复序列。13个IESs中, 除MDS1/MDS3间的AT含量较低外(52.38%), 其他12个IESs均为富含AT序列(62.50%-87.32%)(表 5)。MDSs按照1-3-5-7-9-11-2-4-6-8-10-12-13-14的顺序排列, 标号为奇数的MDSs与标号为偶数的MDSs各自簇集在一起(图 2)。
表 5 Oxytricha sp. (ZJ) 小核α-TBP基因的MDSsIESs及指针序列Table 5. Characteristics of MDSs, IESs, and pointer sequences in Oxytricha sp. (ZJ)α-TBP geneMDSn MDSn的长度Length of MDSn MDSn和(n+1) 间的指针序列Pointer sequence between MDSn and (n+1) MDSi和MDSj间IES的长度Length of IESs between MDSi and MDSj IESs AT含量IESs AT content (%) 1 223 GGTGCAC 1-3 21 52.38 2 282 GTCTYGTA 3-5 33 72.73 3 35 ATTCACA 5-7 29 68.97 4 22 ATTAGAGT 7-9 42 78.57 5 158 TCGCWCATTC 9-11 16 62.50 6 27 AGAAGTC 11-2 50 86.00 7 13 TGCCACT 2-4 20 65.00 8 173 TCAAGCTTAA 4-6 45 73.33 9 21 AGTTTT 6-8 19 68.42 10 48 CTGGTGA 8-10 30 76.67 11 26 ATCAGCTACTT 10-12 25 76.00 12 28 AAAGT 12-13 34 85.29 13 731 TAAGTRAT 13-14 71 87.32 14 312 注: MDSs和IESs的长度都包括了一拷贝的指针序列; 不同克隆间指针序列的差异用兼并碱基表示
Note: Length of MDSs include pointers on both ends; IESs exclude pointer sequences; Use degenerate symbols reflects differences between clones2.3 Oxytricha sp. (ZJ) 的大、小核DNApol α基因结构特征
Oxytricha sp. (ZJ) 大核DNApol α基因全长为5118 bp。含有编码1534个氨基酸的ORF, 其AT含量约为62.15%。含两个长度分别为96 bp (intron-1) 和49 bp (intron-2) 的内含子, AT含量分别为71.88%和69.39%。5′和3′非编码区的序列长度分别为158 bp (含20 bp端粒序列) 和216 bp (含20 bp端粒序列), AT含量分别为71.85%和74.09%。5个阳性克隆测序累计检测到205个碱基变异位点, 其中ORF含有184个碱基变异位点, 涉及39个氨基酸的差异, 未引起移码突变或无义突变。剩余的碱基变异位点, 有5个位于5′非编码区, 5个位于3′非编码区, 8个位于intron-1, 3个位于intron-2。
已获得Oxytricha sp. (ZJ) 的小核DNApol α基因片段长度为2068 bp。包括15个富含AT序列(62.50%-90.91%) 的IESs, 14个包含两端指针序列的完整MDSs (5-7-9-11-13-15-17-19-21-23-25-27-29-30)(表 6), 以及2个MDSs (2和32) 的部分区域。并由此确定了另13个MDSs (1-6-8-10-12-14-16-18-20-22-24-26-28) 的位置(不包含指针序列), 指针序列只能在相连的3个MDSs (29-30-31) 间被确定出来(图 3)。
表 6 Oxytricha sp. (ZJ) 小核DNApol α基因片段MDSs, IESs及指针序列Table 6. Characteristics of MDSs, IESs, and pointer sequences in Oxytricha sp. (ZJ) DNApol α gene segmentMDSn MDSn的长度Length of MDSn MDSn和(n+1) 间的指针序列Pointer sequence between MDSn and (n+1) MDSi和MDSj间IES的长度Length of IESs between MDSi and MDSj IESs AT含量IESs AT content (%) 2 ? ? 2-5 24 62.50 5 39 ? 5-7 22 77.27 7 85 ? 7-9 11 90.91 9 70 ? 9-11 17 82.35 11 39 ? 11-13 20 80.00 13 55 ? 13-15 26 84.62 15 32 ? 15-17 5 80.00 17 101 ? 17-19 42 78.57 19 86 ? 19-21 17 88.24 21 100 ? 21-23 14 85.71 23 82 ? 23-25 31 83.87 25 40 ? 25-27 14 85.71 27 97 ? 27-29 50 74.00 29 418 AACAT 29-30 43 79.09 30 160 TGAAGAA 30-31 85 82.35 31 ? 注: MDSs的长度包括了两端的指针序列; IESs的长度不含指针序列; 问号代表数值无法确定
Note: Length of MDSs include pointers on both ends; IESs exclude pointer sequences; Question marks indicate undetermined information3. 讨论
3.1 尖毛虫属小核Actin Ⅰ基因的乱序模式
Actin Ⅰ基因是腹毛类纤毛虫中最早发现的乱序基因[4]。目前尖毛虫属已知小核Actin Ⅰ基因结构的物种共包括5个:Oxytricha trifallax、O. nova、O. fallax、O. sp. (Aspen)、O. sp. (Misty)。根据Hogan等[12]推测的腹毛类小核Actin Ⅰ基因结构进化路线, 尖毛虫属内小核Actin Ⅰ基因乱序模式分为两种情况:O. sp. (Misty) 与Stylonychia pustulata拥有较为相似的模式(模式1);O. trifallax、O. nova、O. sp. (Aspen) 模式(模式2) 相近。而之后报道的O. fallax与我们新测序的O. sp. (ZJ) 模式可归入模式2。
尖毛虫属内小核Actin Ⅰ基因乱序模式2所涉及的5个物种可以分为2种情况: (1)Oxytricha fal-lax与O. trifallax两个种群Actin Ⅰ的小核基因乱序模式均为3-4-6-5-7-9-10-2-1-8 (模式2.1)。DuBois和Prescott等[12]将三者视为O. trifallax-O. fallax群组, 代表了一个单一种。(2)O. sp. (ZJ) 小核Actin Ⅰ基因由8个IESs和9个MDSs组成, 同属内的O. nova在组成数目上完全相同, 且2个物种的MDSs排列顺序均为3-4-6-5-7-9-2-1-8 (模式2.2)[1], 比模式2.1少了长的IES6与非常短的MDS10。O. sp. (Aspen)[11]仅有部分小核Actin Ⅰ基因完成测序, Hogan等[12]推测其乱序模式与O. nova (模式2.2) 相似, 鉴于数据的不完整性, 在此我们不做详细讨论。
对比基于大核Actin Ⅰ基因序列构建的系统树(图 4a) 与小核Actin Ⅰ基因结构进化路线(图 4b) 可以看出, 二者存在一定的相似之处, 但却不完全相同。在图 4a中作为姐妹枝出现的Oxytricha fallax与O. trifallax的小核Actin Ⅰ乱序模式均为2.1, 拥有10个MDSs (图 4b)。但是, 在大核Actin Ⅰ基因系统树中(图 4a), 同属于小核Actin Ⅰ乱序模式2.2 (图 4b) 的3个物种O. nova、O. sp. (Aspen)、O. sp. (ZJ) 并未聚在一起。这表明, 大小核Actin Ⅰ基因进化路线并不完全一致。
模式2.1、2.2之间, 在IESs的长度方面均具有较大差异。在模式2.1中, Oxytricha trifallax的IESs长度为19-276 bp[10]; 而在模式2.2中, O. sp. (ZJ) 和O. nova的IESs长度分别是11-79 bp (本研究)、11-109 bp[4]。这符合前人的推测:由于IESs的快速进化与易变性, 在进化过程中其长度也发生了改变, 而小核Actin Ⅰ基因进化过程中, IESs的长度比其序列更为保守[15]。凭IESs的序列很难推断进化的图式。
在模式2.1中, Oxytricha trifallax的指针序列长度为2-14 bp[11]; 而在模式2.2中, O. nova和O. sp. (ZJ) 的指针序列长度范围较为接近, 分别为5-13 bp[4]和4-13 bp (表 4)。但相同乱序模式的物种指针序列并未发现具有更多序列相似性。
3.2 尖毛虫属小核α-TBP基因的乱序模式
目前尖毛虫属已知小核α-TBP基因结构的物种共包括2个:Oxytricha trifallax、O. nova。我们的研究表明, O. sp. (ZJ) 小核α-TBP基因由13个IESs和14个MDSs组成, 与同属的O. nova一致[8], 说明在这两个种分化过程中该小核基因没有发生新的MDS片段的重组或易位。与之相对比, O. trifallax多了位于非乱序区的3个MDSs和3个IESs[15](图 2)。猜测这两种不同的模式是由于IES的插入/丢失造成。O. sp. (ZJ) 与O. nova的MDSs排列模式为1-3-5-7-9-11-2-4-6-8-10-12-13-14[8]; 而O. trifallax排列模式为1-3-5-7-10-12-2-4-6-8-9-11-13-14-15-16-17[15]。O. sp. (ZJ)、O. nova中的MDS8、MDS13-14分别与O. trifallax中的MDS8-9、MDS14-17同源(图 6)。
![]() 图 5 Oxytricha sp. (ZJ)、O. nova和O. trifallax大、小核Actin Ⅰ基因结构模式图(参照DuBois等, 1995[10])梯形代表端粒, 黑色圆点代表起始密码子与终止密码子Figure 5. Schematic illustration of macro-and micronuclear Actin Ⅰ gene structures of Oxytricha sp. (ZJ), O. nova and O. trifallax (reference DuBois, et al., 1995[10])Ladder represent telomeres, black dots mark the pointer of start and stop codons
图 5 Oxytricha sp. (ZJ)、O. nova和O. trifallax大、小核Actin Ⅰ基因结构模式图(参照DuBois等, 1995[10])梯形代表端粒, 黑色圆点代表起始密码子与终止密码子Figure 5. Schematic illustration of macro-and micronuclear Actin Ⅰ gene structures of Oxytricha sp. (ZJ), O. nova and O. trifallax (reference DuBois, et al., 1995[10])Ladder represent telomeres, black dots mark the pointer of start and stop codons![]()
Oxytricha sp. (ZJ)、O. nova的IESs数目均为13个, 二者的长度差别不大, 分别为16-71 bp、11-55 bp; 而O. trifallax的IESs数目为16个, 长度为5-108 bp[15]。O. sp. (ZJ) 和O. nova中的IESs10, 11, 12与O. trifallax中的IESs11, 12, 13相对应。与小核Actin Ⅰ的结果相似, O. sp. (ZJ) 与O. nova具有同样α-TBP基因乱序模式, 且IESs的长度较为一致, O. sp. (ZJ)、O. nova、O. trifallax的IES平均长度分别为33、26和35 bp。O. sp. (ZJ)(16 bp) 和O. nova (8 bp) 最短的IES是IES5, 而O. trifallax最短的为IES9 (5 bp)[8, 15]。由此可见, IESs的长度、数目具有物种特异性。
模式2.1的Oxytricha trifallax指针序列长度为7-17 bp[15], 模式2.2的O. nova和O. sp. (ZJ) 的指针序列长度分别为为3-19 bp[8]、5-11 bp (表 5), 长度范围差异大。相同乱序模式的物种指针序列并未发现具有更多序列相似性。
3.3 尖毛虫属小核DNApol α基因的乱序模式
与小核Actin Ⅰ和α-TBP基因相比, DNApol α基因乱序程度更为复杂, 基因序列更长, 乱序模式为非随机排列[9]。目前, 尖毛虫属已知小核DNApol α基因结构的物种共包括2个:Oxytricha trifallax、O. nova。我们的研究未能获取O. sp. (ZJ) 的小核DNApol α基因全长。根据扩增出的O. sp. (ZJ) 小核DNApol α基因部分序列, 我们确定出14个完整MDSs (排列模式为5-7-9-11-13-15-17-19-21-23-25-27-29-30), 并推测出未扩增成功的16个MDSs (MDSs1-2、偶数MDSs6-28、MDSs31-32) 的位置, 同时推测MDS3与MDS4可能丢失。O. trifallax包含至少51个MDSs和50个IESs[17], O. nova包含至少45个MDSs和44个IESs[9]。通过比对分析(图 7), 我们发现O. sp. (ZJ) 含有3个非乱序排列的MDSs, 分别是MDS29、MDS30、MDS31, 而在O. trifallax和O. nova中分别是9个和7个非乱序排列的MDSs[17]。
![]()
模式2.1的Oxytricha trifallax指针序列长度为2-15 bp[17], 模式2.2的O. nova指针序列长度为2-16 bp[9]。本研究中未获得O. sp. (ZJ) 的DNApol α基因序列全长, 很多指针序列无法推测(表 6), 故而本研究未对该基因不同物种间指针序列进行对比。
我们确定出的Oxytricha sp. (ZJ) 小核DNApol α基因的14个完整MDSs位于倒置区域。O. sp. (ZJ) 的MDS7对应O. nova的MDS6, 与O. trifallax的MDS7-8-9同源, 并且MDS8在O. trifallax的小核基因主体部分中是缺失的[17]。据此合理推断是: (1) 此区域由3个MDSs组成, 且中间MDS的缺失代表了祖先种的基因结构特征。O. trifallax从祖先种分离时保留了该结构特征。而在O. sp. (ZJ) 和O. nova分化前的进化过程中将3个MDSs融合成了一个MDS。(2) 为第一种可能的反向进程, 即祖先种在此区域为一个MDS结构, 而在O. trifallax从祖先种分化时, 独立的发生了一次重组将中间的部分转离了小核基因的主体部分, 类似移位现象, 使得一个MDS变成了3个MDSs。之前Wong等[16]的研究表明, 在种系分离自共同祖先后, 独立发生编码区域的重新划分和融合。
因Oxytricha sp. (ZJ) 的MDS7与O. trifallax的MDS7-8-9同源, 且O. trifallax在此处的IES与O. sp. (ZJ) 的MDS7对应区域的序列完全一致(图 8), 表明在O. sp. (ZJ) 的进化过程中可能将该IES同化成了MDS的一部分, 即发生IES向MDS的转换, 伴随此种转换, 使得原先3个MDSs融合成了一个MDS, 而原先位于中间的MDS被丢弃。这个转换过程呈现出腹毛类纤毛虫小核基因在进化过程中方式的多样性。这支持了上述第一种解释, 即在O. sp. (ZJ) 和O. nova分化前的进化过程中将3个MDSs融合成了一个MDS。Duharcourt等[30]在Paramecium中与Chalker与Yao[31]在Tetrahymena中都发现了旧大核的IESs在新大核中的保留现象。而Mollenbeck等[19]通过比较Stylonychia lemnae两种群的小核Actin Ⅰ基因发现, 发现IES向MDS转变的迹象, 但这种转变发生在非编码区, 未对基因的开放阅读框造成影响。本实验则发现编码区内IES向MDS的转变, 使得本应删除的序列成为基因组永久保留的一部分。
![]() 图 8 Oxytricha sp. (ZJ) 进化过程中IES向MDS的转变Figure 8. IES to MDS transformation during Oxytricha sp. (ZJ) evolution
图 8 Oxytricha sp. (ZJ) 进化过程中IES向MDS的转变Figure 8. IES to MDS transformation during Oxytricha sp. (ZJ) evolution -
![]()
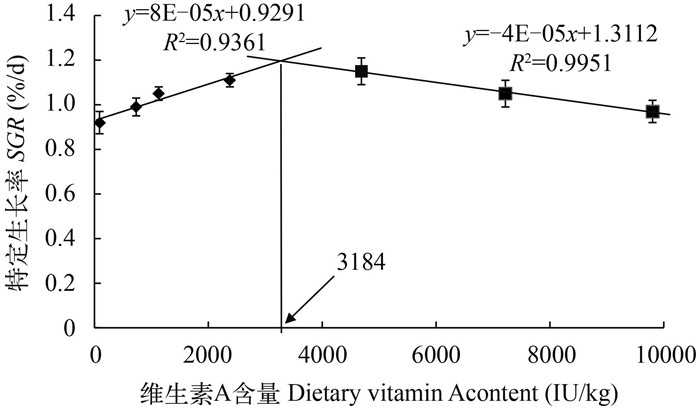
图 1 维生素A对养成期草鱼特定生长率的影响
以特定生长率进行折线模型回归得出养成期草鱼对维生素A的适宜需求量为3184 IU/kg饲料
Figure 1. The effect of dietary VA level on SGR of subadult grass carp fed experimental diets
VA requirement derived from SGR by the broken-line model is 3184 IU/kg diet
![]()
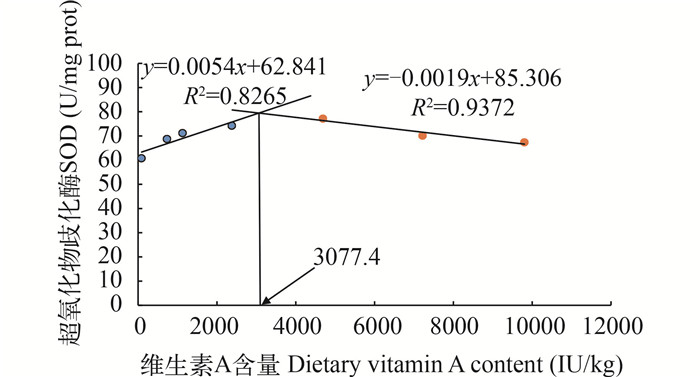
图 2 维生素A对养成期草鱼血清超氧化物歧化酶活性的影响
以SOD为判断标准, 通过折线模型得出养成期草鱼对维生素A的适宜需求量为3077.4 IU/kg饲料
Figure 2. Effect of dietary VA level on SOD activity of subadult grass carp fed experimental diets
VA requirement derived from SOD activity by the broken-line model is 3077.4 IU/kg diet
表 1 基础饲料配方和基本组成
Table 1 Ingredients and chemical composition of the basal diet for grass carp
原料Ingredient % 脱脂鱼粉Defatted fish meal 5.00 酪蛋白Casein 24.66 明胶Gelatin 9.00 α-淀粉α-starch 24.00 玉米淀粉Corn starch 21.16 亚麻油Linseed oil 1.47 大豆油Soybean oil 1.38 棕榈油Palm oil 1.20 不含维生素A维生素预混料Vitamin A free vitamin premixa 1.00 矿物质预混料Mineral premixb 2.00 维生素A的预混料Vitamin A premixc 1.00 磷酸二氢钙Ca (H2PO4)2 2.30 氯化胆碱Choline chloride (50%) 1.00 乙氧基喹啉Ethoxy quinoline (30%) 0.05 纤维素Cellulose 5.00 DL-蛋氨酸DL-Met (99%) 0.18 总计Total 100.00 营养成分Proximate composition 粗脂肪Crude fat 4.23 粗灰分Crude ash 4.13 水分Moisture 9.52 粗蛋白Crude protein 32.81 总能Gross energy (KJ/g) 19.16 注: a.每千克不含维生素A维生素预混料含:维生素D3 (500000 IU/g), 0.480 g; DL-a生育酚醋酸酯(50%), 20.00 g; 维生素K3(23%), 0.22 g; 维生素B12(1%), 0.1 g; D-生物素(2%), 5 g; 叶酸(96%), 0.52 g; 盐酸硫胺素(90%), 0.13 g; 维生素C醋酸酯(99%), 7.16 g; 烟酸(99%), 2.58 g; 肌醇(99%), 52.33 g; D-泛酸钙pantothenate (98%), 3.07 g; 核黄素(80%), 0.99 g; 吡哆醇(81%), 0.75 g; 玉米淀粉, 906.67 g; b.每千克矿物质元素预混料: MgSO4·7H2O (9.86% Mg), 350.91 g; FeSO4·7H2O (19.7% Fe), 38.07 g; CuSO4·5H2O (25.00% Cu), 0.60 g; ZnSO4·7H2O (22.5% Zn), 6.67 g; MnSO4·H2O (31.8% Mn), 2.04 g; KI (3.8% I), 1.45 g; Na2SeO3 (1% Se), 2.50 g; corn starch, 597.76 g; c.含维生素A预混料根据饲料试验设计的维生素A含量来添加, 同时调整玉米淀粉的含量
Note: a. Per kilogram of vitamin A free vitamin premix containing: vitamin D3 (500000 IU/g), 0.480 g; α-tocopherol acetate (50%), 20.00 g; vitamin K3 (23%), 0.22 g; vitamin B12 (1%), 0.1 g; D-biotin (2%), 5 g; folic acid (96%), 0.52 g; thiamin (90%), 0.13 g; L-ascorbic acid (99%), 7.16 g; niacin (99%), 2.58 g; inositol (99%), 52.33 g; D-calcium pantothenate (98%), 3.07 g; riboflavin (80%), 0.99 g; pyridoxine (81%), 0.75 g; corn starch, 906.67 g; b. Per kilogram of mineral premix containing: MgSO4·7H2O (9.86% Mg), 350.91 g; FeSO4·7H2O (19.7% Fe), 38.07 g; CuSO4·5H2O (25.00% Cu), 0.60 g; ZnSO4·7H2O (22.5% Zn), 6.67 g; MnSO4·H2O (31.8% Mn), 2.04 g; KI (3.8% I), 1.45 g; Na2SeO3 (1% Se), 2.50g; corn starch, 597.76 g; c. Vitamin A premixes were compounded by adding vitamin A acetate according to the designed vitamin A content in the diet to replace the equal quantity of cornstarch in the control diet 下载: 导出CSV
下载: 导出CSV
表 2 饲料中不同维生素A含量对养成期草鱼生长、摄食和存活率活性的影响
Table 2 Effects of dietary vitamin A concentrations on growth performance, feeding and survival rate of subadult grass carp
饲料维生素A含量
Dietary VA level (IU/kg)初均重
IW (g)特定生长率
SGR (%/d)饲料系数
FCR摄食率
FR (%BW/d)存活率
Survival (%)肝体比
HSI (%)脏体比
VSI (%)89 261.67±14.46 0.92±0.05a 2.26±0.01c 2.62±0.12 90.0±2.9 2.25±0.13 6.92±0.21a 732 260.47±15.71 0.99±0.04abc 2.07±0.12bc 2.52±0.16 91.7±1.7 2.35±0.21 7.63±0.08ab 1129 246.63±7.11 1.05±0.03abc 1.99±0.05ab 2.60±0.06 95.0±2.9 2.21±0.26 7.60±0.34ab 2378 242.90±15.83 1.11±0.03bc 1.88±0.07ab 2.57±0.20 90.0±2.9 2.28±0.25 7.89±0.60ab 4688 252.50±8.04 1.15±0.06c 1.66±0.09a 2.35±0.06 93.3±1.7 2.30±0.33 8.03±0.28ab 7218 258.33±11.21 1.05±0.06abc 1.91±0.11ab 2.48±0.07 93.3±1.7 2.06±0.05 8.05±0.43ab 9802 259.73±7.37 0.97±0.05ab 2.12±0.08bc 2.55±0.03 90.0±2.9 2.02±0.18 8.66±0.24b 注:表中数值(平均值±标准误)为3个重复的平均值; 数值后不同的上标字母表示差异显著(P < 0.05);下同
Note: Values (mean±SE) are the means of triplicate. Values in each column with different superscripts showed significant difference (P < 0.05); the same applies below
下载: 导出CSV
表 3 饲料中不同维生素A含量对养成期草鱼血清SOD活性、MDA、ALP、Ca、P含量的影响
Table 3 Effects of dietary vitamin A concentrations on serum biochemistry of subadult grass carp
饲料维生素A含量
Dietary VA level (IU/kg)超氧化物歧化酶
SOD (U/mg prot)碱性磷酸酶
ALP (U/L)丙二醛
MDA (mmol/L)钙
Ca (mmol/L)磷
P (mmol/L)89 60.77±4.67a 97.80±2.40 9.79±0.48 2.82±0.12 2.72±0.08 732 68.70±1.26abc 73.43±32.47 9.69±1.49 2.28±0.02 2.08±0.35 1129 71.14±2.65bc 84.70±10.28 10.71±0.64 2.88±0.01 2.20±0.32 2378 74.20±1.12bc 106.63±4.43 10.59±0.83 2.76±0.17 2.39±0.17 4688 77.10±2.72c 118.10±24.70 9.95±0.80 2.75±0.12 3.36±1.13 7218 70.08±1.49bc 104.80±20.35 9.41±1.38 2.79±0.01 3.08±0.06 9802 67.33±2.03ab 121.56±10.40 9.35±2.18 2.89±0.11 2.41±0.13
下载: 导出CSV
表 4 饲料中不同维生素A含量对养成期草鱼全鱼常规成分含量的影响
Table 4 Effect of dietary vitamin A concentrations on whole body proximate composition of subadult grass carp
饲料维生素A含量
Dietary VA level (IU/kg)水分
Moisture (%)粗蛋白
Crude protein (%)粗脂肪
Crude lipid (%)灰分
Crude ash (%)89 71.46±1.11ab 16.55±0.17 8.85±1.06 3.62±0.14 732 73.04±0.98b 16.76±0.51 7.55±0.52 3.76±0.06 1129 73.31±0.98b 16.35±0.31 7.82±0.42 3.91±0.18 2378 72.26±0.67ab 16.41±0.55 8.18±0.38 3.44±0.16 4688 71.76±1.35ab 16.48±0.93 7.89±0.06 3.77±0.42 7218 71.87±0.56ab 16.46±0.45 8.47±0.41 3.74±0.12 9802 69.23±0.29a 16.80±0.33 8.66±0.30 3.92±0.16
下载: 导出CSV
表 5 维生素A对养成期草鱼头骨脊椎骨灰分、钙、磷含量以及钙磷含量比值的影响
Table 5 Effects of dietary vitamin A concentrations on Ca, P and ash content and Ca/P in skull and vertebra of subadult grass carp (g/kg in dry matter)
饲料维生素A含量
Dietary VA level (IU/kg)头骨Skull 脊椎骨Vertebra 钙Ca 磷P 钙/磷Ca/P 灰分Ash 钙Ca 磷P 钙/磷Ca/P 灰分Ash 89 205.3±0.4ab 98.4±0.5ab 2.10±0.01a 518.7±2.9b 197.3±11.1a 98.0±3.2a 2.12±0.01 456.8±14.3a 732 207.2±5.0ab 98.6±2.7ab 2.13±0.02ab 517.4±11.8b 211.5±4.7ab 101.4±2.2ab 2.14±0.01 466.8±10.7ab 1129 199.9±1.9a 95.0±0.9a 2.13±0.01ab 514.7±5.9b 212.2±4.9ab 101.2±2.4ab 2.16±0.01 480.6±11.4ab 2378 214.5±4.7bc 104.8±1.5c 2.11±0.01ab 538.5±4.5c 215.0±7.9ab 102.5±4.1ab 2.13±0.01 484.2±20.7ab 4688 224.4±2.9c 110.2±0.2d 2.15±0.02b 543.7±4.5c 225.5±0.7b 108.3±1.0b 2.16±0.02 504.0±2.1b 7218 223.5±2.3c 107.4±1.8cd 2.13±0.01ab 540.3±3.5c 225.7±0.2b 107.7±0.9b 2.13±0.02 505.3±3.5b 9802 208.9±4.4ab 102.2±3.6bc 2.11±0.01ab 485.5±9.1a 212.9±8.1ab 100.5±4.4ab 2.12±0.02 458.4±16.3a
下载: 导出CSV
-
[1] Halver J E. The Vitamins. In Fish Nutrition[M]. USA California:Academic Press. 2002, 61-141
[2] Moren M, Opstad I, Berntssen M H G, et al. An optimum level of vitamin A supplements for Atlantic halibut (Hippoglossus hippoglossus L.) juveniles[J]. Aquaculture, 2004, 235(1):587-599 https://www.researchgate.net/publication/223891956_An_optimum_level_of_Vitamin_A_supplements_for_Atlantic_halibut_Hippoglossus_hippoglossus_L_juveniles
[3] Hemre G I, Deng D F, Wilson R P, et al. Vitamin A metabolism and early biological responses in juvenile sunshine bass (Morone chrysops×M. saxatilis) fed graded levels of vitamin A[J]. Aquaculture, 2004, 235(1):645-658
[4] 蒋明.草鱼幼鱼对维生素A, D和K需要量的研究.硕士学位论文.华中农业大学, 武汉. 2007 Jiang M. Studies on requirements of dietary Vitamin A, D and K for grass carp (Ctenopharyngodon idellus) fingerling[D]. Huazhong Agricultural University, Wuhan. 2007
蒋明.草鱼幼鱼对维生素A, D和K需要量的研究.硕士学位论文.华中农业大学, 武汉. 2007[5] Takeuchi T, Dedi J, Haga Y, et al. Effect of vitamin A compounds on bone deformity in larval Japanese flounder (Paralichthys olivaceus)[J]. Aquaculture, 1998, 169(3):155-165 http://www.academia.edu/6719092/Effect_of_vitamin_A_compounds_on_bone_deformity_in_larval_Japanese_flounder_Paralichthys_olivaceus
[6] Shaik Mohamed J, Sivaram V, Roy T S C, et al. Dietary vitamin A requirement of juvenile greasy grouper (Epinephelus tauvina)[J]. Aquaculture, 2003, 219(1):693-701 https://www.researchgate.net/publication/222086391_Dietary_vitamin_A_requirement_of_juvenile_greasy_grouper_Epinephelus_tauvina
[7] Hernandez L H H, Teshima S I, Ishikawa M, et al. Die-tary vitamin A requirements of juvenile Japanese flounder Paralichthys olivaceus[J]. Aquaculture Nutrition, 2005, 11(1):3-9 doi: 10.1111/anu.2005.11.issue-1
[8] National Research Council (NRC):Nutrient Requirements of Fish and Shrimp[M]. National Academy of Science, Washington, DC
[9] Cuesta A, Ortuño J, Rodriguez A, et al. Changes in some innate defence parameters of seabream (Sparus aurata L.) induced by retinol acetate[J]. Fish & Shellfish Immunology, 2002, 13(4):279-291 https://www.researchgate.net/publication/11025612_Changes_in_some_innate_defence_parameters_of_seabream_Sparus_aurata_L_induced_by_retinol_acetate?_sg=l9GQ946V-36PMR44_RUMeqGS9U2yQzsc7O1L59not2B46BlLrUVSNzccRnu5xQV2Tqpl_3o3vbMZxsF-U4u3gw
[10] Hu C J, Chen S M, Pan C H, et al. Effects of dietary vi-tamin A or β-carotene concentrations on growth of juve-nile hybrid tilapia, Oreochromis niloticus×O. aureus[J]. Aquaculture, 2006, 253(1):602-607 https://www.researchgate.net/publication/250043680_Vitamin_A_in_diets_for_Nile_tilapia
[11] 张璐.鲈鱼和大黄鱼几种维生素的营养生理研究和蛋白源开发.博士研究生学位论文.中国海洋大学, 青岛. 2006 Zhang L. The nutritional physiology of several vitamins and the potential of several protein sources as replacement of fish meal for Japanese seabass (Lateolabrax japonicus C.) and large yellow croaker (Pseudosciaena crocea R)[D]. Ocean University of China, Qingdao. 2006
张璐.鲈鱼和大黄鱼几种维生素的营养生理研究和蛋白源开发.博士研究生学位论文.中国海洋大学, 青岛. 2006[12] 魏玉婷.大菱鲆(Scophthalmus maximus)幼鱼对饲料中蛋氨酸, 精氨酸, 维生素A及维生素E需求量的研究.硕士学位论文.中国海洋大学, 2010 Wei Y T. Studies on requirements of dietary methionine, arginine, vitamin A, vitamin E for turbot (Scophthalmus maximus)[D]. Ocean University of China, Qingdao. 2006
魏玉婷.大菱鲆(Scophthalmus maximus)幼鱼对饲料中蛋氨酸, 精氨酸, 维生素A及维生素E需求量的研究.硕士学位论文.中国海洋大学, 2010[13] Chen C, Zhu W, Wu F, et al. Quantifying the dietary potassium requirement of subadult grass carp (Ctenopharyngodon idellus)[J]. Aquaculture Nutrition, 2015, DOI: 10.1111/anu.12279
[14] 李彬, 梁旭方, 刘立维, 等.饲料蛋白水平对大规格草鱼生长, 饲料利用和氮代谢相关酶活性的影响.水生生物学报, 2014, 38(2):233-240 http://ssswxb.ihb.ac.cn/CN/abstract/abstract2291.shtml Li B, Liang X F, Liu L W, et al. Effects of dietary protein levels on growth, feed utilization and the enzymes acti-vity on nitrogen metabolism of grass carp (Ctenopharyngodon idellus)[J]. Acta Hydrobiologica Sinica, 2014, 38(2):233-240
李彬, 梁旭方, 刘立维, 等.饲料蛋白水平对大规格草鱼生长, 饲料利用和氮代谢相关酶活性的影响.水生生物学报, 2014, 38(2):233-240 http://ssswxb.ihb.ac.cn/CN/abstract/abstract2291.shtml[15] Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J]. Analytical Biochemistry, 1976, 72(1):248-254 https://www.researchgate.net/publication/281498068_A_Rapid_and_Sensitive_Method_for_Quantitation_of_Microgram_Quantities_of_Protein_Utilizing_the_Principle_of_Protein-Dye_Binding
[16] Otsuki T, Shimizu K, Iemitsu M, et al. Chlorella intake attenuates reduced salivary SIgA secretion in kendo training camp participants[J]. Nutrition Journal, 2012, 11(1):103-110 doi: 10.1186/1475-2891-11-103
[17] Wen H, Yan A S, Gao Q, et al. Dietary vitamin A requirement of juvenile Amur sturgeon (Acipenser schrenckii)[J]. Journal of Applied Ichthyology, 2008, 24(5):534-538 doi: 10.1111/jai.2008.24.issue-5
[18] Villeneuve L A N, Gisbert E, Moriceau J, et al. Intake of high levels of vitamin A and polyunsaturated fatty acids during different developmental periods modifies the expression of morphogenesis genes in European sea bass (Dicentrarchus labrax)[J]. British Journal of Nutrition, 2006, 95(4):677-687 doi: 10.1079/BJN20051668
[19] Rudnicki M, Silveira M M, Pereira T V, et al. Protective effects of Passiflora alata extract pretreatment on carbon tetrachloride induced oxidative damage in rats[J]. Food and Chemical Toxicology, 2007, 45(4):656-661 doi: 10.1016/j.fct.2006.10.022
[20] Bannister J V, Bannister W H, Rotilio G. Aspects of the structure, function, and applications of superoxide dismutase[J]. CRC Critical Reviews in Biochemistry, 1987, 22(2):111-180 doi: 10.3109/10409238709083738
[21] Muñoz M, Cedeño R, Rodrıguez J, et al. Measurement of reactive oxygen intermediate production in haemocytes of the penaeid shrimp, Penaeus vannamei[J]. Aquaculture, 2000, 191(1):89-107 http://www.docin.com/p-1285077306.html
[22] D'Aquino M, Dunster C, Willson R L. Vitamin A and glutathione-mediated free radical damage:competing reactions with polyunsaturated fatty acids and vitamin C[J]. Biochemical and Biophysical Research Communications, 1989, 161(3):1199-1203 doi: 10.1016/0006-291X(89)91369-7
[23] Beattie J H, Avenell A. Trace element nutrition and bone metabolism[J]. Nutrition Research Reviews, 1992, 5(1):167-188 doi: 10.1079/NRR19920013
[24] Sakamoto S, Yone Y. A principal source of deposited li-pid in phosphorus deficient red sea bream (Chrysophrys major)[J]. Bulletin of the Japanese Society of Scientific Fisheries (Japan), 1980, 46:1227-1230 doi: 10.2331/suisan.46.1227
[25] Retina Society Terminology Committee. The classification of retinal detachment with proliferative vitreoretinopathy[J]. Ophthalmology, 1983, 90(2):121-125 doi: 10.1016/S0161-6420(83)34588-7
[26] Conaway H H, Pirhayati A, Persson E, et al. Retinoids stimulate periosteal bone resorption by enhancing the protein RANKL, a response inhibited by monomeric glucocorticoid receptor[J]. Journal of Biological Chemistry, 2011, 286(36):31425-31436 doi: 10.1074/jbc.M111.247734
[27] Martinez G M, Baron M P, Bolker J A. Skeletal and pigmentation defects following retinoic acid exposure in larval summer flounder, Paralichthys dentatus[J]. Journal of the World Aquaculture Society, 2007, 38(3):353-366 doi: 10.1111/jwas.2007.38.issue-3
[28] Negm R K, Cobcroft J M, Brown M R, et al. The effects of dietary vitamin A in rotifers on the performance and skeletal abnormality of striped trumpeter (Latris lineata) larvae and post larvae[J]. Aquaculture, 2013, 404:105-115 https://www.researchgate.net/publication/256917410_The_effects_of_dietary_vitamin_A_in_rotifers_on_the_performance_and_skeletal_abnormality_of_striped_trumpeter_Latris_lineata_larvae_and_post_larvae
[29] Lall S P, Lewis-McCrea L M. Role of nutrients in skele-tal metabolism and pathology in fish-an overview[J]. Aquaculture, 2007, 267(1):3-19 https://www.researchgate.net/profile/Santosh_Lall/publication/223160282_Role_of_nutrients_in_skeletal_metabolism_and_pathology_in_fish_-_An_overview/links/558aac9508ae48b7b56d6d16.pdf
[30] Samarut E, Fraher D, Laudet V, et al. ZebRA:An overview of retinoic acid signaling during zebrafish development[J]. Biochimica et Biophysica Acta (BBA)-Gene Re-gulatory Mechanisms, 2015, 1849(2):73-83 doi: 10.1016/j.bbagrm.2014.05.030
-
期刊类型引用(1)
1. 何燕,尹家奇,生欣. 艾美游仆虫大核基因组与转录组测序及结构特征. 水生生物学报. 2021(05): 1014-1023 .  本站查看
本站查看
其他类型引用(0)








计量
- 文章访问数: 1574
- HTML全文浏览量: 439
- PDF下载量: 23
- 被引次数: 1