
CLONING AND EXPRESSION ANALYSIS OF ALPHA AMYLASE CDNA OF PENAEUS MONODON
-
摘要: 为了研究斑节对虾α-淀粉酶基因的结构和生物学功能, 根据原实验室构建的斑节对虾(Penaeus monodon) cDNA文库得到的EST序列, 利用RACE技术获得了斑节对虾α-淀粉酶基因(PmAmy)的cDNA全长序列。该基因序列全长2465 bp, 包括2175 bp的开放阅读框, 编码724个氨基酸, 分子总量为78.9 kD, 理论等电点为4.66。PmAmy包含一个α-淀粉酶家族保守的A结构域(Thr34-Ser410)和一个C结构域(Glu420-Ala496)。PmAmy氨基酸序列与其他物种的相似性为47%—99%, 利用PmAmy构建的进化树显示斑节对虾和凡纳滨对虾(Litopenaeus vannamei)的亲缘关系最近。基因表达结果显示PmAmy在肝胰腺组织中的表达量显著高于其他组织(P<0.05)。斑节对虾PmAmy基因在卵巢发育的过程中均有表达, 表达量有所变化, 虽然没有发现显著性的差异(P=0.09)。斑节对虾PmAmy在整个生长阶段的检测中都有表达, 其中幼体发育过程中存在显著性差异, 糠虾时期PmAmy表达量显著高于无节幼体、溞状幼体和仔虾时期(P<0.05)。以上实验结果初步说明了PmAmy可能与斑节对虾的幼体发育相关。Abstract: In order to study the structure and biological function of the alpha-amylase gene in Penaeus monodon, the full-length cDNA sequence of α-Amylase from Penaeus monodon (PmAmy) was obtained by high throughput transcriptome sequencing and RACE. The PmAmy cDNA included an open reading frame of 2175 bp encoding a polypeptide of 724 amino acids, and the predicted molecular mass and isoelectric point were 78.9 kD and 4.66, respectively. The PmAmy contained a conservative A domain (Thr34-Ser410) and a C domain (Glu420-Ala496) of alpha amylase family. Homology analysis revealed that the PmAmy shared 47%—99% identity to other known amylase sequences, and the phylogenetic tree showed that the PmAmy was closely related to Litopenaeus vannamei. The expression levels of PmAmy in hepatopancreas were significantly higher than those of the other tissues (P<0.05). ThePmAmy expression was found in five ovarian development stages. The expression level in yolky stage (stageⅣ) was the highest among the five stages, and was the lowest in ovogonium stage (stageⅠ). The expression levels of PmAmy showed no significantly difference in ovary development stages. Expression of PmAmy was detected in all tested growth stages, and the expression level in mysis was significantly higher than that in nauplius, zoea and post larval (P<0.05). These results suggested thatPmAmy might be associated with larval development in P. monodon.
-
Keywords:
- Penaeus monodon /
- α-Amylase /
- Gene cloning /
- Gene expression /
- Larval development
-
鱼类早期资源指精卵结合至早期稚鱼阶段, 是鱼类个体生活史的开端[1]。通过对鱼类早期资源的研究, 可以了解鱼类的繁殖活动、产卵场位置分布和繁殖时间等早期生活史特征[1]。同时, 早期资源也是影响种群数量变化的主要原因之一[2]。然而, 捕捞、水利工程建设和江湖阻隔等人类活动造成了鱼类早期资源的急剧减少[3, 4]。例如, Sanches等[5]研究结果显示, 水利工程导致巴拉那河的鱼苗密度有显著变化。过度捕捞造成地中海沿海区域鱼类的产卵活动明显减少[6]。
长江中游是多种鱼类的产卵场和栖息地。刘明典等[7]2014—2015年在宜昌江段调查到27种鱼类卵苗。李世健等[8]在监利江段调查到51种鱼类的鱼卵和仔稚鱼。黎明政等[9]在武穴江段调查到27种鱼类的卵苗。同时, 长江中游也是重要经济鱼类四大家鱼的主要产卵场之一。20世纪60年代, 长江中游四大家鱼的产卵场有25处, 产卵量占长江总量的四分之三。其中, 宜昌江段是四大家鱼产卵规模最大的产卵场, 平均鱼卵径流量可达119.48×108粒, 占长江干流总量的9.7%[10]。20世纪80年代, 在葛洲坝修建之后, 长江中游宜昌至城陵矶江段仍有11处四大家鱼产卵场, 产卵规模为11.14×108粒, 占长江干流总规模的25.8%[11]。但是, 在三峡水库蓄水之后, 长江中游四大家鱼的繁殖活动受到了不利的影响[12]。同时, 过度捕捞, 尤其是对仔幼鱼的捕捞, 造成了鱼类资源的急剧减少[13]。此外, 三峡水库蓄水之后造成长江中游宜昌江段四大家鱼产卵时间推迟约20d, 监利江段宜昌江段四大家鱼的产卵规模减少约75%[14]。为了有效地开展长江鱼类资源的保护工作, 有必要掌握长江中游鱼类早期资源的现状及其变化规律。
本研究通过在宜都断面开展产漂流性卵鱼类早期资源调查, 了解宜昌江段产漂流性卵鱼类的繁殖情况; 结合历史资料, 分析四大家鱼及其他鱼类的繁殖规模和时间变化; 分析水文因子对产漂流性卵鱼类优势种类繁殖的影响; 为长江中游鱼类早期资源保护工作的开展提供科学依据。
1. 材料与方法
1.1 样本的采集
2017年5月12日至7月25日和2018年5月10日至7月24日, 在葛洲坝下游宜都市清江河口上约1.5—3.0 km处设置采样断面(112°26′E, 30°24′N)(图 1)。采用产漂流性卵鱼类早期资源的调查方法。采样网具包括弶网和圆锥网, 采集方法参照曹文宣等[1]和黎明政等[14]的方法。弶网用于每日采样, 记录种类和数量。圆锥网用于采集断面数据, 计算断面系数, 每年采集一次。弶网网口呈半圆形, 网口直径1 m, 网目尺寸500 μm, 网口面积0.393 m2, 网长2 m。网后部连接集苗箱, 集苗箱长40 cm, 宽和高均为30 cm, 上部敞口, 用于收集鱼卵和仔鱼。弶网采集在每日上午和下午各一次, 采集时间为7:30—9:00和16:30—18:00, 每次采集持续30—60min。同时, 测量采样点水温、透明度、流速等环境参数, 水位和流量数据来自于全国水雨情网站(http://xxfb.mwr.cn/)。圆锥网用于断面定量采样。网口呈圆形, 网口直径0.5 m, 网口面积0.196 m2, 网长2 m, 网目尺寸500 μm。圆锥网设置在断面左岸近岸点、左岸至江中心1/2处、江中心、江中心至右岸1/2处以及右岸近岸处5个采样点, 每个采样点设置表层、中层和底层3个采样水层(表层、中层和底层距水面的距离分别占水深的20%、50%和80%), 每个水层采集15min。
![]() 图 1 2017年和2018年长江中游宜都断面鱼类早期资源采样图Figure 1. Samling map of fish eggs and larvae in the Yidu section of the Yangtze River in 2017 and 2018
图 1 2017年和2018年长江中游宜都断面鱼类早期资源采样图Figure 1. Samling map of fish eggs and larvae in the Yidu section of the Yangtze River in 2017 and 20181.2 种类鉴定
鱼卵按照卵膜性质(漂流性、黏性和浮性)分类, 并依据卵径大小、胚体长等标准将大小不同的鱼卵区分培养, 培养水温维持在20—25℃, 直至孵出发育至分类特征明显的阶段, 按照形态、色素分布、肌节数目等进行种类鉴定, 具体方法参照曹文宣等[1]; 仔鱼通过显微镜观察进行种类鉴定, 之后样本用95%的乙醇保存。形态观测不能确定种类的样本用95%的乙醇固定保存在−20℃冰箱, 带回分子实验室, 通过线粒体DNA细胞色素b基因进行种类鉴定, 具体实验方法参照Xiao等[15]的方法进行。
1.3 数据处理
产卵规模计算 鱼卵和仔鱼通过断面径流量的估算方法参照易伯鲁等[10]、黎明政等[14]和母红霞等[16]的方法进行, 具体计算公式如下:
$A = M + M'\qquad\qquad\qquad\qquad\qquad\qquad$

$M = \sum\nolimits_{i = 1}^n {{T_i}} {D_i}{Q_i}C\qquad\qquad\qquad\qquad\quad$

$\begin{aligned}{M' } =& \sum\limits_{i = 1}^{n - 1} {T_{\left( {i,i + 1} \right)}'} \times C \times \left[ {\left( {{Q_i} + {Q_{i + 1}}} \right)/2} \right] \times \;\;\;\\& \left[ {\left( {{D_i} + {D_{i + 1}}} \right)/2} \right]\end{aligned}$

${D_{{i}}} = {N_i}/\left( {S\;{V_i}\;{T_i}} \right)\qquad\qquad\qquad\qquad\qquad\;\;$

$\;C = \left( {\sum\limits_{{{j}} = 1}^m {{D_j}} } \right)/\left( {m\;{D_r}} \right)\qquad\qquad\qquad\qquad$

式中, A为整个采样期内流经采集点江段面的鱼卵、仔鱼总流量; M为所有定时采集时间内鱼卵、仔鱼流量之和; M'为前后两次相邻采集之间非采集时间内计算出的鱼卵、仔鱼流量之和; i为某一特定的常规采样; i+1为第i次采样之后的一次采样; Qi为第i次采样时流经采样断面的水流量(m3/s); Di 为第i次采样时流经采样断面的仔鱼和鱼卵密度(ind./m3); C 为采样断面上的鱼卵和仔鱼的分布系数; T (i,i+1) 为第i次采样和第i+1次采样之间的时间间隔; Ni为第i次采样中采集到的鱼卵和仔鱼数量; S为网口面积(m2); Vi为第i次采样中网口的水流速(m/s); Ti为第i次采样的持续时间(s); Dj为断面采样每个采样点的鱼卵和仔鱼密度(ind./m3); j为第j个断面采样位点; m为断面采样的采样点数量; Dr为断面采样中流经靠近右岸的采样点表层的鱼卵和仔鱼密度(ind./m3)。
确定产漂流性卵鱼类优势种类 物种优势度利用Pinkas相对重要性指数(IRI, Index of relative abundance)[17]确定, 计算公式如下:
$IRI = \left( {N{\text{%}} + W{\text{%}} } \right) \times F{\text{%}} $

式中, N为某一种类鱼卵数量占采获鱼卵总数量的百分比; W为某种鱼卵生物量比例; F为某一种类出现的次数占调查总次数的百分比。由于鱼卵、鱼苗很小, 生物量难以确定, 因此只考虑数量这一因素。将IRI大于1000的种类定义为优势种。
四大家鱼产卵场位置推算 四大家鱼产卵场位置的计算公式如下:
$L = V \times T$

式中, L为四大家鱼卵、苗的漂流距离(m); V为调查江段水流速度(m/s); T为胚胎发育所需要的时间(s)。其中, 四大家鱼胚胎发育所需要的时间参照易伯鲁等[18]。
繁殖活动的水文条件分析 根据相关研究文献资料, 近些年一段断面所采集的仔鱼大多来源于三峡大坝以上江段, 鱼卵来自三峡大坝下游产卵场[14, 19]。因此,鱼类繁殖水文因子影响分析仅针对通过鱼卵鉴定的产漂流性卵鱼类的优势种类。首先, 对物种的密度及水文因子数据进行除趋势对应分析(Detrended correspondence analysis, DCA), 根据每个轴的梯度长度(Lengths of gradient, LGA)选择适合的分析方法。当LGA<3时, 采用冗余分析(Redundancy Analysis, RDA); 当LGA>4时, 采用典型对应分析(Canonical Correlation Analysis, CCA); 当4>LGA>3时, 两种分析方法皆可[20]。DCA分析结果轴梯度长度为1.8, 故本研究利用冗余分析(Redundancy Analysis,RDA)分析鱼卵密度与环境因子(水温、流量、流量日变化、透明度)之间的相关关系。
DCA和RDA分析利用Canoco 5.0[21]软件完成, 其余数据分析与作图用Excel 2010软件中完成。
2. 结果
2.1 种类组成及优势种
2017年和2018年共采集鱼卵21120粒和仔鱼2123尾。物种鉴定结果显示, 鱼卵和仔鱼共5目9科37种。其中, 鲤科(Cyprinidae) 23种, 鳅科(Cobitidae) 5种, 鲿科(Bagridae)及科(Sisoridae)各2种, 银鱼科(Salangidae)、平鳍鳅科(Homalopteridae)、鱵科(Hemiramphidae)、鮨科(Serranidae)和鰕虎鱼科(Gobiidae)各1种, 鲤科鱼类资源数量为20788粒, 占总数量的89.44%,其中鲌亚科为优势类群, 占鲤科鱼类总数量的39.45%。根据卵的性质, 漂流性卵24种, 黏性卵5种, 浮性卵1种, 沉性卵7种(表 1)。
表 1 2017年和2018年调查期间宜都断面鱼类早期资源种类组成Table 1. Species composition of eggs and larvae in the Yidu section in 2017 and 2018目Order 科Family 种Species 卵类型Type of egg 卵Eggs 苗Larvae 目Order 科Family 种Species 卵类型Type of egg 卵Eggs 苗Larvae 鲑形目Salmoniforme
银鱼科Salangidae太湖新银鱼Neosalanx taihuensis 沉性 + 圆筒吻Rhinogobio cylindricus 漂流性 + + 鲤形目Cypriniformes
鲤科Cyprinidae青鱼Mylopharyngodon piceus 漂流性 + + 蛇Saurogobio dabryi 漂流性 + + 草鱼Ctenopharyngodon idellus 漂流性 + + 光唇蛇Saurogobio gymnocheilus 漂流性 + 赤眼鳟Squaliobarbus curriculus 漂流性 + + 宜昌鳅鮀Gobiobotia filifer 漂流性 + 鳡Elopichthys bambusa 漂流性 + + 鲤Cyprinus capio 黏性 + 飘鱼Pseudolaubuca sinensis 黏性 + 鲫Carassius auratus 黏性 + 寡鳞飘鱼Pseudolaubuca engraulis 漂流性 + + 平鳍鳅科Homalopteridae 犁头鳅Lepturichthys fimbriata 漂流性 + + 贝氏䱗Hemiculter bleekeri 漂流性 + + 鳅科Cobitidae 花斑副沙鳅Parabotia fasciata 漂流性 + + 翘嘴鲌Culter alburnus 黏性 + + 武昌副沙鳅Parabotia banarescui 漂流性 + 鳊Parabramis pekinensis 漂流性 + + 紫薄鳅Leptobotia taeniops 漂流性 + 银鲴Xenocypris argentea 漂流性 + + 红唇薄鳅Leptobotia rubrilabris 漂流性 + 黄尾鲴Xenocypris davidi 黏性 + 鲇形目Siluriformes
鲿科Bagridae瓦氏黄颡鱼Pelteobagrus vachelli 沉性 + 似鳊Pseudobrama simoni 漂流性 + 光泽黄颡鱼Pelteobaggrus nitidus 沉性 + 鳙Hypophthalmichthys nobilis 漂流性 + + 科Sisoridae 中华纹胸Glyptothorax sinensis 沉性 + 鲢Hypophthalmichthys molitrix 漂流性 + + 福建纹胸Glyptothorax fukiensis 沉性 + 银Squalidus argentatus 漂流性 + + 颌针鱼目Beloniformes
鱵科Hemiramphidae间下鱵Hyporhamphus intermedius 沉性 + 铜鱼Coreius heterodon 漂流性 + + 鲈形目Perciformes
鮨科Serranidae鳜Siniperca chuatsi 浮性 + + 吻Rhinogobio typus 漂流性 + 鰕虎鱼科Gobiidae 子陵吻鰕虎鱼Rhinogobius giurinus 沉性 + 注: “+”代表可采集到Note: “+” means can be collected 2017年和2018年采集到鱼卵28种, 有青鱼(Mylopharyngodon piceus)、草鱼(Ctenopharyngodon idellus)、赤眼鳟(Squaliobarbus curriculus)、贝氏䱗(Hemiculter bleekeri)、鳙(Hypophthalmichthys nobilis)、鲢(Hypophthalmichthys molitrix)和银(Squalidus argentatus)等种类。采集到仔鱼27种, 有太湖新银鱼(Neosalanx taihuensis)、寡鳞飘鱼(Pseudolanbuca engraulis)、贝氏䱗、间下鱵(Hyporhamphus intermedius)和子陵吻鰕虎鱼(Rhinogobius giurinus)等种类。
2017年, 采集鱼卵10572粒, 仔鱼1221尾, 其中四大家鱼卵3172粒, 贝氏䱗卵2725粒, 分别占卵总数量的30.0%、25.8%; 仔鱼以贝氏䱗和寡鳞飘鱼为主, 分别占比54.3%、24.4%。2018年共采集鱼卵10548粒, 仔鱼902尾, 其中四大家鱼卵4249粒, 贝氏䱗卵2448粒, 分别占卵总数量的40.3%和23.2%; 仔鱼以子陵吻鰕虎鱼、寡鳞飘鱼和贝氏䱗为主, 分别占比22.7%、19.3%和17.7%。根据2017年和2018年采集卵和仔鱼数量分析, 宜都断面鱼卵优势种为四大家鱼(IRI=1433)和贝氏䱗(IRI=1319); 仔鱼优势种为贝氏䱗(IRI=1259)和寡鳞飘鱼(IRI=1169)。
2.2 产卵时间
2017年5月采集鱼卵以银和飘鱼为主, 占当月总卵数的27.13%和19.38%, 6月四大家鱼和贝氏䱗成为主要种类, 数量占比分别为38.89%和26.12%。在7月四大家鱼数量占比下降至18.50%, 而贝氏䱗上升至43.24%。2018年稍有不同, 四大家鱼数量百分比自5月的55.38%逐渐下降至7月的15.04%, 贝氏䱗相对比例依旧在6月达到最高, 占当月数量的42.77%, 7月保持稳定, 占当月数量的41.00%。2017年和2018年调查期间, 飘鱼和银所占数量比自5月至7月逐渐减少, 而花斑副沙鳅和鳊则相反, 自5月比例逐渐上升。2017年四大家鱼的产卵时间是从5月14日到7月2日。2018年则是从5月10日至7月5日。
仔鱼5月采集较少, 2017年5月仅采集仔鱼5尾, 分别为间下鱵、光泽黄颡鱼和光唇蛇, 2018年5月采集仔鱼16尾, 分别为太湖新银鱼、寡鳞飘鱼和贝氏䱗。仔鱼采集数量主要在6月和7月。2017年和2018年采集结果显示, 仔鱼中贝氏䱗和寡鳞飘鱼为优势种类(图 2)。
![]() 图 2 2017年和2018年宜都断面主要类群鱼卵(a)、仔鱼(b)的相对丰度Figure 2. Abundance percentage of eggs (a) and larvae (b) of dominant species in the Yidu section in 2017 and 2018
图 2 2017年和2018年宜都断面主要类群鱼卵(a)、仔鱼(b)的相对丰度Figure 2. Abundance percentage of eggs (a) and larvae (b) of dominant species in the Yidu section in 2017 and 20182.3 产卵规模
2017年, 通过宜都断面的鱼卵径流量为124.45×108粒, 鱼卵中四大家鱼径流量为36.13×108粒, 占总径流量的29.03%; 贝氏䱗径流量为19.96×108粒, 占总径流量的16.04%。2018年通过宜都断面的鱼卵径流量为101.07×108粒, 鱼卵中四大家鱼径流量为35.20×108粒, 占总径流量的34.83%; 贝氏䱗的鱼卵径流量为19.17×108粒, 占总径流量的18.97%。
2017年通过宜都断面的仔鱼径流量为16.43×108尾, 寡鳞飘鱼和贝氏䱗径流量分别为5.89×108尾和2.91×108尾, 占仔鱼总径流量的35.87%和17.72%。四大家鱼的总径流量为4.12×108尾, 占仔鱼总径流量的25.07%。2018年通过宜都断面的仔鱼径流量为8.29×108尾, 寡鳞飘鱼和贝氏䱗分别为0.97×108尾和1.22×108尾, 占仔鱼总径流量的11.72%和14.70%。四大家鱼的总径流量为0.11×108尾, 占仔鱼总径流量的1.32%。
2.4 四大家鱼的产卵场位置
通过对采集到的四大家鱼卵发育期的观测, 0.14%处于128细胞期前, 95.31%处于桑椹期至眼囊期, 4.55%处于尾芽期至心脏搏动期, 距受精1—33h。其中, 65.76%的四大家鱼卵处于原肠早期至原肠晚期, 距受精7—13h。四大家鱼仔鱼均处于鳔雏形期和鳔一室期, 距受精98—146h。根据鱼卵和仔鱼的发育期以及流速分析, 四大家鱼鱼卵来源于宜都断面2.4 km以上的江段。产卵场位置主要分布在葛洲坝下至庙咀、胭脂坝至云池江段。四大家鱼仔鱼应来源于三峡大坝以上江段(图 3)。
![]() 图 3 长江中游宜昌江段四大家鱼产卵场位置Figure 3. Spawning grounds of the four major Chinese carps in the Yichang reach in the middle Yangtze River
图 3 长江中游宜昌江段四大家鱼产卵场位置Figure 3. Spawning grounds of the four major Chinese carps in the Yichang reach in the middle Yangtze River2.5 产漂流性卵鱼类及关键环境因子
2017年最小流量在7月4日, 为6920 m3/s, 最大流量在7月11日, 为30200 m3/s。整个调查期间, 水温介于18.5—26.0℃, 5月12日达到18℃以上, 7月20日达到24℃以上。2017年出现4次产卵高峰, 分别在6月10日、6月11日、6月18日和7月10日。高峰期产卵规模总计为60.7×108粒, 占2017年总产卵规模的48.7%。
2018年6月12日流量最小, 为10200 m3/s, 最大流量在7月14日, 为43800 m3/s。整个调查期间, 水温为18.5—25.2℃, 5月10日达到18℃以上, 7月2日达到24℃以上。2018年出现2次显著的产卵高峰, 分别为5月19日和6月24日, 产卵规模共为35.9×108粒, 占2018年采集鱼卵总规模的35.5%(图 4)。
![]() 图 4 2017年(a)和2018年(b)调查期间宜都断面鱼卵日密度变化图Figure 4. Discharge and eggs density drifting through the Yidu section in 2017 (a) and 2018 (b)
图 4 2017年(a)和2018年(b)调查期间宜都断面鱼卵日密度变化图Figure 4. Discharge and eggs density drifting through the Yidu section in 2017 (a) and 2018 (b)应用RDA分析方法, 对鱼卵漂流密度与水文因子的关系进行分析。在前2个排序轴上, 物种与水文因子的相关性为0.426和0.345, 第一排序轴和第二排序轴的累积贡献率为92.83%。RDA分析结果显示, 四大家鱼卵总密度和花斑副沙鳅卵密度受流量日变化和流量影响较大; 贝氏䱗鱼卵密度受流量影响较大; 鳊鱼卵密度受水温影响较大; 银鱼卵密度主要受透明度影响(图 5)。
![]() 图 5 2017年和2018年宜都断面水文因子对漂流性卵密度的影响Figure 5. Effects of hydrological factors on the density of the drifting eggs in the Yidu section in 2017 and 2018
图 5 2017年和2018年宜都断面水文因子对漂流性卵密度的影响Figure 5. Effects of hydrological factors on the density of the drifting eggs in the Yidu section in 2017 and 20183. 讨论
3.1 长江中游鱼类繁殖现状及变化
本研究结果显示, 2017年和2018年5—7月通过长江中游宜都断面的鱼类早期资源较丰富。已有的研究结果显示, 母红霞[22]发现2009年和2010年5—7月通过宜都断面鱼卵径流量分别为57.26×108粒和64.45×108粒, 年均60.86×108粒。与2009年和2010年同比, 2017年和2018年通过宜都断面的鱼卵径流量明显增加, 年均径流量增加了85.3%。其中, 宜昌江段四大家鱼的产卵规模明显增大。2005—2012年5—7月通过宜都断面四大家鱼鱼卵径流量为(0.03—5.85)×108粒, 年均2.49×108粒[14], 而2017年和2018年四大家鱼卵年均35.67×108粒, 同比增加了13倍, 达到了20世纪80年代的水平[10, 11]。鱼卵径流量增加的主要原因可能有2个。第一, 2009年起, 原农业部在长江中游开展较大规模的四大家鱼亲鱼放流, 增加了繁殖群体数量[7, 23]。第二, 2011年三峡工程开始生态调度试验, 在一定程度上促进了鱼类的繁殖活动。徐薇等[24]的研究结果显示, 2012年三峡水库试验性生态调度期间长江中游沙市江段鱼卵总径流量和家鱼卵径流量均出现高峰, 繁殖种类明显增加。周雪等[25]的研究结果显示, 2013—2017年三峡水库生态调度造成长江中游监利江段涨水持续时间达到4d及以上, 期间四大家鱼的卵苗量较大, 占年总量的45%以上。
在三峡水库蓄水运行之后, 长江中游四大家鱼的产卵时间推迟。在三峡水库蓄水之前, 长江中游宜昌江段四大家鱼产卵时间从4月中下旬开始[10]。在蓄水后, 2005—2012年宜昌江段四大家鱼的产卵时间推迟到5月8日至5月28日, 平均推迟了21.8d, 导致产卵时间推迟的主要原因是下泄水温滞后, 到达18℃的时间推迟[14]。与2005—2012年相比, 2017年和2018年宜昌江段四大家鱼产卵时间没有明显变化, 产卵场位置也没有明显变化。因此, 本研究的结果表明, 在三峡水库蓄水运行之后, 长江中游四大家鱼产卵场的环境条件可能已经趋于稳定。如果亲本增加, 宜昌江段产卵场依然可以满足其完成繁殖活动, 产生较大规模的鱼卵和仔鱼。
本研究结果还显示, 2017年和2018年5—7月有一定规模的仔鱼通过长江中游宜都断面, 其中四大家鱼的仔鱼均来源于三峡大坝以上江段。《长江三峡水利枢纽环境影响报告书》认为, 在三峡水库蓄水之后, 长江上游涪陵以下的8个四大家鱼产卵场将消失, 产卵场将上移。根据母红霞[22]、王红丽等[26]、Li等[14]和马琴[27]在三峡水库库区的鱼类早期资源调查结果显示, 涪陵以下江段仍有产卵规模较小的四大家鱼的产卵场。2005—2012年, 通过宜都断面四大家鱼仔鱼径流量年均值为4.65×108尾[14]。2013年和2014年分别为1.52×108尾和1.27×108尾[27]。与2005—2014年相比, 2017年和2018年没有明显的变化。研究结果表明, 三峡水库库区内在一定水文条件下仍具备四大家鱼繁殖的条件, 且可以通过三峡大坝进入长江中游, 可以为长江中游鱼类资源提供一定的补充。
3.2 鱼类早期资源与环境因子的关系
本研究结果显示, 四大家鱼、贝氏䱗和花斑副沙鳅等种类的产卵规模与流量和流量日变化有着密切的关系。鱼类的繁殖活动受外界环境因子的影响[28]。已有的研究提出“最佳环境窗口”假说, 认为每个物种都有特定的环境条件需求, 以满足繁殖活动和保证产生最多的后代[29—31]。Li等[32]的研究结果显示, 四大家鱼的产卵活动需要18—24℃的水温和0.55 m/d的水位涨幅。Wang等[12]也发现, 四大家鱼的仔鱼丰度与流量增加持续时间显著相关。因此, 水温应该是四大家鱼等产漂流性卵鱼类的繁殖限制条件, 达到了一定水温鱼类才能开始繁殖活动[10, 33]。流量等则是刺激鱼类繁殖和促进繁殖活动的重要环境条件[10, 34]。
3.3 长江中游鱼类早期资源保护措施
鱼类早期资源减少导致种群补充量减少[35, 36], 是造成长江鱼类资源急剧减少的主要原因之一[1, 37]。Ma等[38]的研究结果显示, 长江中游对于通江湖泊中鱼类早期资源的补充, 维持鱼类多样性有重要意义。为了长江鱼类资源保护, 2019年农业农村部、财政部和人力资源社会保障部联合印发《长江流域重点水域禁捕和建立补偿制度实施方案》, 长江禁渔已经逐步实施, 2021年长江将全面禁渔。这一措施将消除捕捞对鱼类资源的影响, 尤其是对湖区等仔幼鱼资源的影响, 能够显著提高鱼类早期生活史阶段的存活率, 增加种群的补充量。长江鱼类的早期生活史阶段还面临着栖息地减少的问题。长江中下游的湖泊和河漫滩原本是仔幼鱼的主要栖息地。但是, 长江中下游的通江湖泊仅剩石臼湖、洞庭湖和鄱阳湖, 其余湖泊都已经被阻隔, 仔幼鱼无法进入湖泊生长, 其栖息地功能已经完全丧失。Liu等[39]的研究结果显示, 长江中下游江湖连通性减少已经严重影响了鱼类多样性。同时, 港口建设、防洪护坡和城市景观等建设破坏了河漫滩的自然环境条件, 减少了仔幼鱼的栖息地[40, 41]。因此, 为了更加有效地开展鱼类资源保护, 结合本研究结果, 还建议通过恢复江湖连通、灌江纳苗, 以及维持河漫滩自然环境条件, 增加仔幼鱼的栖息地面积; 开展重要经济鱼类亲本放流, 增加繁殖群体规模; 开展生态调度, 促进繁殖活动。
致谢:
感谢中国科学院水生生物研究所胡华明、李文静、杨萍、张文武和张晨在样本采集、数据处理上给予的帮助!
-
![]()
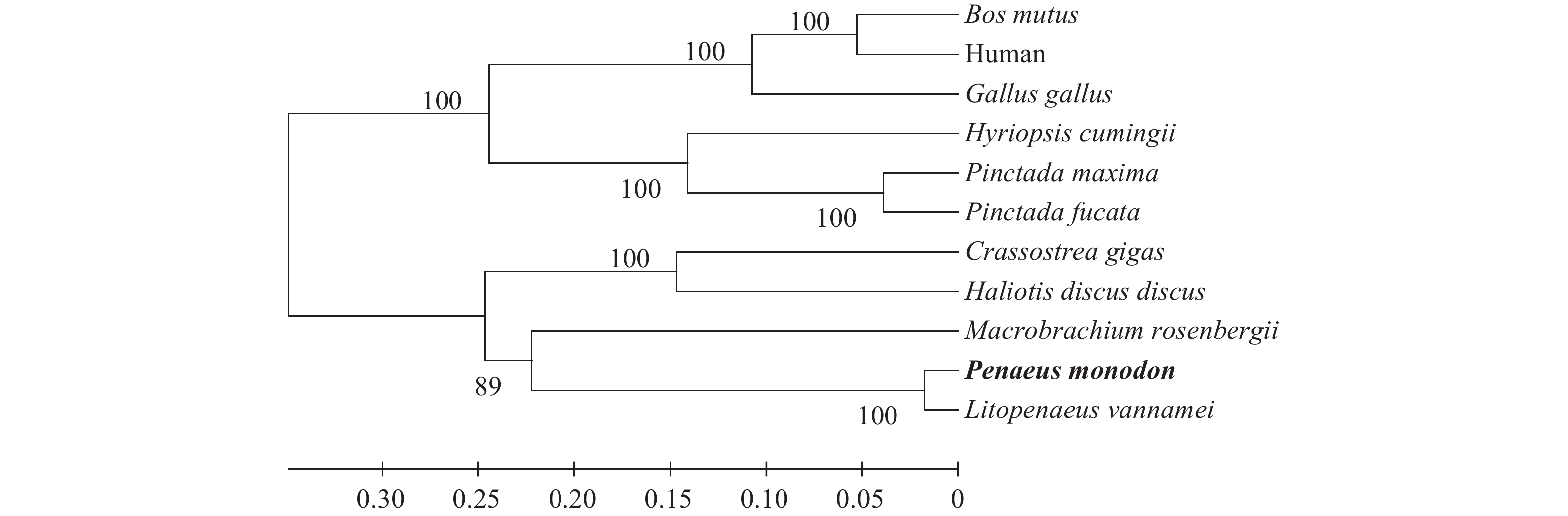
图 1 利用Clustal W程序和MEGA 5.0软件构建的PmAmy系统进化树
物种AMY序列: 所用物种序列斑节对虾KU308415; 凡纳滨对虾AIJ02079.1; 罗氏沼虾AKL71614.1; 牛XP_005898010.1; 人AAA35525.1; 鸡NP_001001528.1; 大珠母贝HM989014.1; 三角帆蚌AGW45296.1; 合浦珠母贝AGN55419.1; 长牡蛎EKC28393.1; 盘鲍ABO26611.1
Figure 1. Phylogenetic tree of the AMY amino acid sequence in different groups by program Clustal W and MEGA 5.0
The AMY sequence No. of each species: P. monodon KU308415; L. vannamei AIJ02079.1; M. rosenbergii AKL71614.1; Bos mutus XP_005898010.1; Human AAA35525.1; Gallus gallus NP_001001528.1; Pinctada maxima HM989014.1; Hyriopsis cumingii AGW45296.1; Pinctada fucata AGN55419.1; Crassostrea gigas EKC28393.1; Haliotis discus discus ABO26611.1
![]()
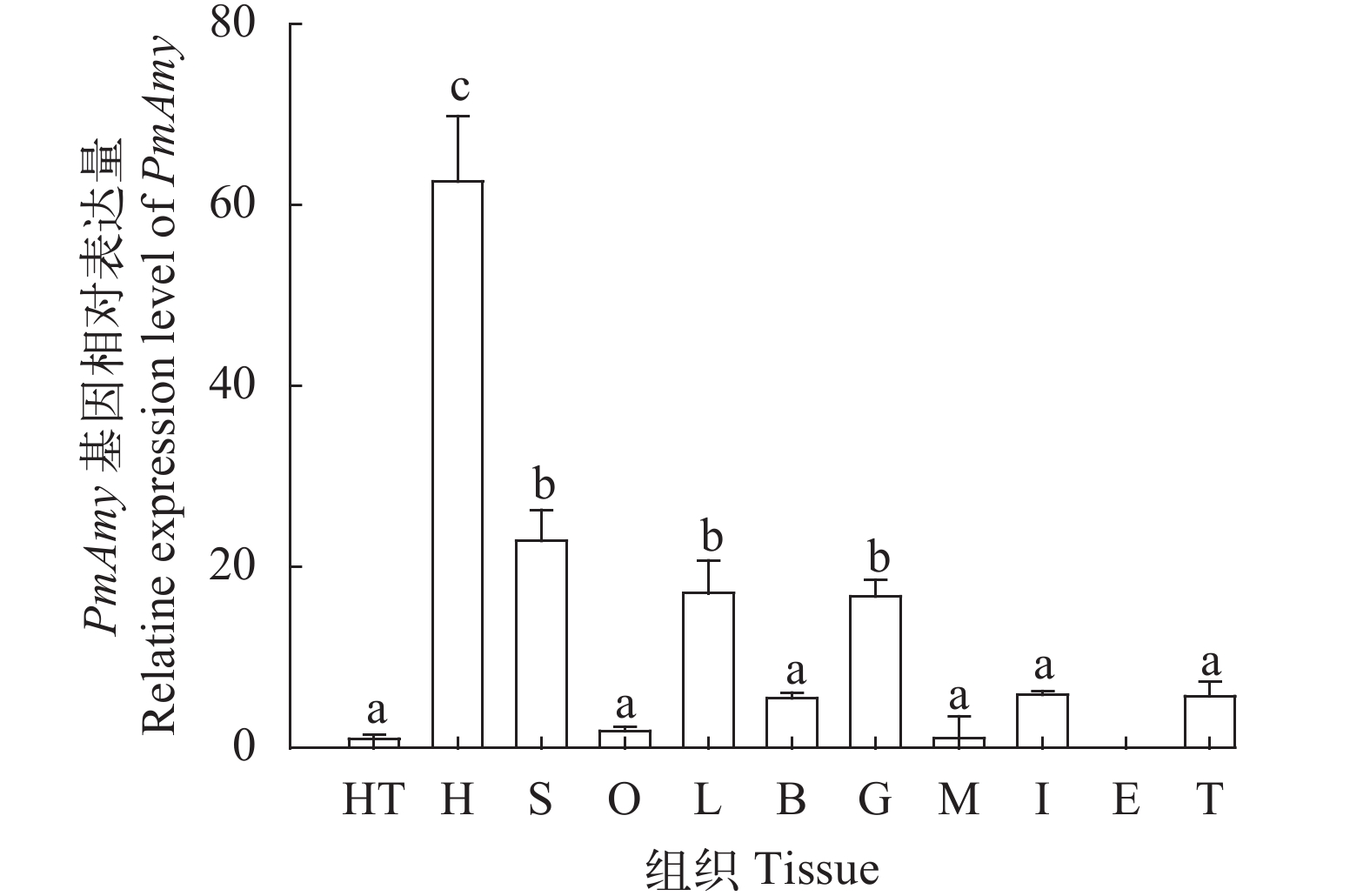
图 2 节对虾PmAmy基因在不同组织中的表达分布
垂直线表示平均值±标准差(n=3); 不同字母表示存在显著性差异(P<0.05); L. lymph淋巴; H. hepatopancreas肝胰腺; G. gill鳃; I. intestine肠; O. ovary卵巢; S. stomach胃; T. thoracic abdominal nerve胸腹神经; M. muscle肌肉; E. eyestalk眼柄; B. brain脑; HT. heart心脏
Figure 2. Distribution of PmAmy gene expression in different tissues of P. monodon
Vertical bars represent the mean±SD (n=3). Significant differences are indicated with different letters above vertical bars (P<0.05)
![]()
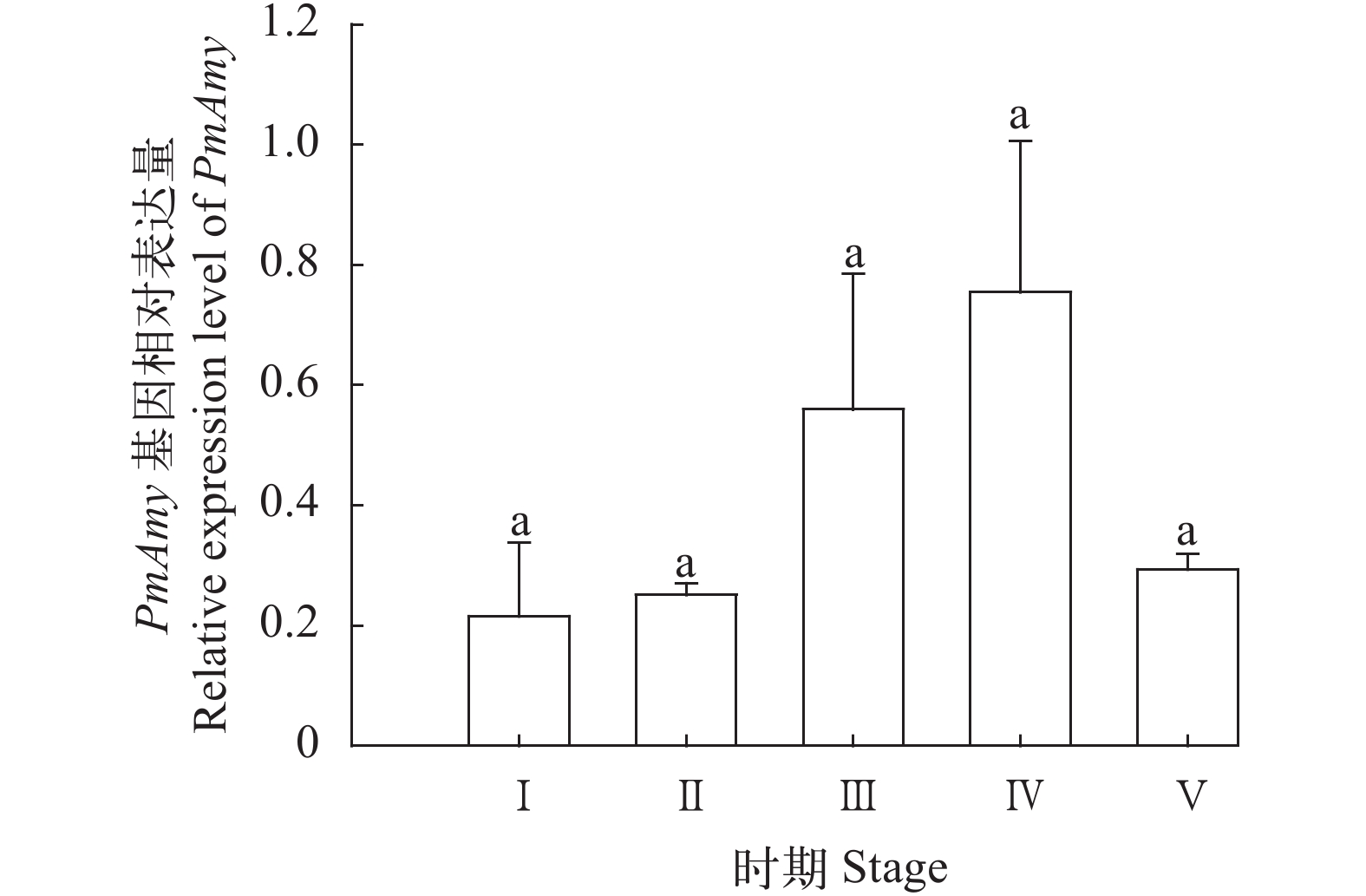
图 3 斑节对虾PmAmy基因在卵巢发育各时期的表达特征
Ⅰ期. 卵原细胞期; Ⅱ期. 核染色质期; Ⅲ期. 周边核仁期; Ⅳ期. 卵黄囊期; Ⅴ期. 成熟期
Figure 3. Relative expression level of PmAmy mRNA in different development stages of ovary
Ⅰ. ovogonium stage; Ⅱ. chromatin nucleolus stage; Ⅲ. perinucleolus stage; Ⅳ. yolky stage; Ⅴ. cortical rod stage
![]()
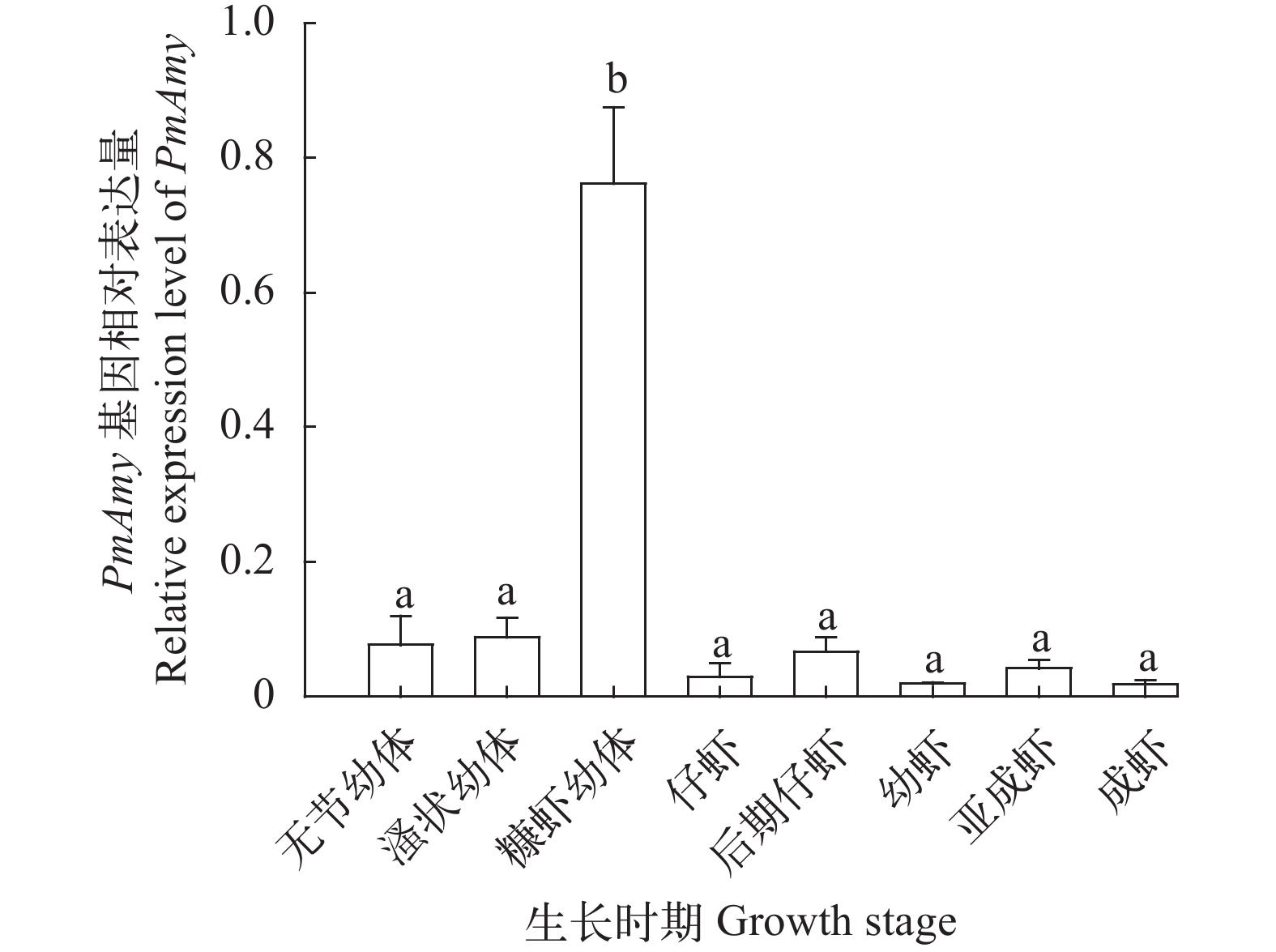
图 4 斑节对虾PmAmy基因在不同生长时期的表达
垂直线表示平均值±标准差(n=3);不同字母表示存在显著性差异(P<0.05)
Figure 4. Expression level of PmAmy in different growth stage of P. monodon
Vertical bars represent the mean±SD (n=3); Significant differences are indicated with different letters above vertical bars (P<0.05)
表 1 实验中所用引物序列
Table 1 Primers sequences used in this experiment
引物Primer 引物序列Sequence (5′—3′) 用途Application AMY-F1 ATGTCATCCTATGATTTTGGGG RACE PCR AMY-F2 AGGTCATCATCGCCGTCT AMY (RT)-F AAGGACACCACACTTACAA Real-time PCR AMY (RT)-R CCACGGAAGCCATTCAT βactin-F GGTATCCTCACCCTCAAGTA Real-time PCR βactin-R AAGAGCGAAACCTTCATAGA EF-F ATGGTTGTCAACTTTGCCCC 检测cDNA EF-R TTGACCTCCTTGATCACACC  下载: 导出CSV
下载: 导出CSV
-
[1] Kuriki T, Imanaka T. The concept of the alpha-amylase family: Structural similarity and common catalytic mechanism [J]. Journal of Bioscience and Bioengineering, 1999, 87(5): 557—565 doi: 10.1016/S1389-1723(99)80114-5
[2] Jung H, Lyons R E, Hurwood D A, et al. Genes and growth performance in crustacean species: a review of relevant genomic studies in crustaceans and other taxa [J]. Reviews in Aquaculture, 2013, 5(2): 77—110 doi: 10.1111/raq.2013.5.issue-2
[3] 阮国良. 草鱼、鳡和翘嘴鲌胰蛋白酶和淀粉酶的早期发育. 硕士学位论文, 华中农业大学. 2010 http://cdmd.cnki.com.cn/Article/CDMD-10504-2010271840.htm Ruan G L. Early development of trypsins and amylases of grass carp Ctenopharyngodon idellus, yellowcheck carp Elopichthys bambusa and topmouth culter Culter alburnus [D]. Thesis for Master of Science, Huazhong Agricultural University. 2010
阮国良. 草鱼、鳡和翘嘴鲌胰蛋白酶和淀粉酶的早期发育. 硕士学位论文, 华中农业大学. 2010 http://cdmd.cnki.com.cn/Article/CDMD-10504-2010271840.htm[4] 陈亮, 梁旭方, 王琳, 等. 鳜鱼胰蛋白酶和淀粉酶与胃蛋白酶原基因的克隆与序列分析. 中国生物化学与分子生物学报, 2009, 25(12): 1115—1123 Chen L, Liang X F, Wang L, et al. Molecular cloning and sequences analysis of trypsin, amylase and pepsinogen genes from Chinese perch (Siniperca chuatsi) [J]. Chinese Journal of Biochemistry and Molecular Biology, 2009, 25(12): 1115—1123
陈亮, 梁旭方, 王琳, 等. 鳜鱼胰蛋白酶和淀粉酶与胃蛋白酶原基因的克隆与序列分析. 中国生物化学与分子生物学报, 2009, 25(12): 1115—1123[5] Boer P H, Hickey D A. The alpha-amylase gene in Drosophila melanogaster: nucleotide sequence, gene structure and expression motifs [J]. Nucleic Acids Research, 1986, 14(21): 8399—8411 doi: 10.1093/nar/14.21.8399
[6] 朱书礼, 李新辉, 杨计平, 等. 草鱼α-淀粉酶基因5′侧翼序列克隆与分析. 南方水产科学. 2014, 10(5): 87—93 Zhu S L, Li X H, Yang J P, et al. Cloning and analysis of 5' flanking sequence of α-amylase gene from Ctenopharyngodon idellus [J]. South China Fisheries Science, 2014, 10(5): 87—93
朱书礼, 李新辉, 杨计平, 等. 草鱼α-淀粉酶基因5′侧翼序列克隆与分析. 南方水产科学. 2014, 10(5): 87—93[7] Sellos D, Moal J, Degremont L, et al. Structure of amylase genes in populations of pacific cupped oyster (Crassostrea gigas): Tissue expression and allelic polymorphism [J]. Marine Biotechnology, 2003, 5(4): 360—372 doi: 10.1007/s10126-002-0089-7
[8] 李璐. 合浦珠母贝α-淀粉酶基因结构、SNP筛选、生长关联与生态响应研究. 硕士学位论文, 上海海洋大学. 2013 http://cdmd.cnki.com.cn/Article/CDMD-10264-1014013536.htm Li L. Genomic structure, SNP screening, growth association and ecological response of α-amylase gene in the pearl oyster Pinctada fucata [D]. Thesis for Master of Science, Shanghai Ocean University. 2013
李璐. 合浦珠母贝α-淀粉酶基因结构、SNP筛选、生长关联与生态响应研究. 硕士学位论文, 上海海洋大学. 2013 http://cdmd.cnki.com.cn/Article/CDMD-10264-1014013536.htm[9] Vanwormhoudt A, Bourreau G, Lemoullac G. Amylase polymorphism in crustacea decapoda-electrophoretic and immunological studies [J]. Biochemical Systematics and Ecology, 1995, 23(2): 139—149 doi: 10.1016/0305-1978(94)00090-4
[10] Takii K, Shimeno S, Kedeka M. Changes in digestive enzyme activities in eel after feeding [J]. Bulletin of the Japanese Society of Scientific Fisheries, 1985, 51(12): 2027—2031 doi: 10.2331/suisan.51.2027
[11] Munilla-Morán R, Stark J R. Metabolism in marine flatfish VI. Effect of nutritional state on digestion in turbot, Scophthulmus maximus (L.) [J]. Comparative Biochemistry and Physiology Part B, 1990, 95(3): 625—634 doi: 10.1016/0305-0491(90)90032-O
[12] Kawai S, Lkeda S. Studies on digestive enzymes of fishes: I. Carbohydrases in digestive organs of several fishes [J]. Nsugaf, 1971, 37(4): 333—337 doi: 10.2331/suisan.37.333
[13] Huvet A, Jeffroy F, Fabioux C, et al. Association among growth, food consumption-related traits and amylase gene polymorphism in the Pacific oyster Crassostrea gigas [J]. Animal Genetics. 2008, 39(6): 662—665 doi: 10.1111/age.2008.39.issue-6
[14] Prudence M, Moal J, Boudry P, et al. An amylase gene polymorphism is associated with growth differences in the Pacific cupped oyster Crassostrea gigas [J]. Animal Genetics, 2006, 37(4): 348—351 doi: 10.1111/age.2006.37.issue-4
[15] Glenn KL, Grapes L, Suwanasopee T, et al. SNP analysis of AMY2 and CTSL genes in Litopenaeus vannamei and Penaeus monodon shrimp [J]. Animal Genetics, 2005, 36(3): 235—236 doi: 10.1111/age.2005.36.issue-3
[16] 辛静静, 刘小林, 李喜莲, 等. 凡纳滨对虾α-淀粉酶基因PCR-RFLP多态性与生长性状的相关分析. 海洋学报(中文版), 2011, 33(3): 124—130 Xin J J, Liu X L, Li X L, et al. PCR-RFLP polymorphism of alpha-amylase gene and its association with growth traits of Litopenaeus vannamei [J]. Acta Oceanologica Sinica, 2011, 33(3): 124—130
辛静静, 刘小林, 李喜莲, 等. 凡纳滨对虾α-淀粉酶基因PCR-RFLP多态性与生长性状的相关分析. 海洋学报(中文版), 2011, 33(3): 124—130[17] 黄建华, 马之明, 周发林, 等. 南海北部野生斑节对虾卵巢解剖结构及组织学的研究. 南方水产, 2005, 1(3): 49—53 Huang J H, Ma Z M, Zhou F L, et al. Study on anatomy structure and histology of the ovary of wild Penaeus monodon (Fabricius) from the north of South China Sea [J]. South China Fisheries Science, 2005, 1(3): 49—53
黄建华, 马之明, 周发林, 等. 南海北部野生斑节对虾卵巢解剖结构及组织学的研究. 南方水产, 2005, 1(3): 49—53[18] 黄建华, 周发林, 马之明, 等. 南海北部斑节对虾卵巢发育的形态及组织学观察. 热带海洋学报, 2006, 25(3): 47—52 Huang J H, Zhou F L, Ma Z M, et al. Morphological and histological observation on ovary development of Penaeus monodon from northern South China Sea [J]. Journal of Tropical Oceanography, 2006, 25(3): 47—52
黄建华, 周发林, 马之明, 等. 南海北部斑节对虾卵巢发育的形态及组织学观察. 热带海洋学报, 2006, 25(3): 47—52[19] 江世贵, 杨丛海, 周发林, 等. 斑节对虾种虾繁育技术. 北京: 海洋出版社. 2013, 80—86 Jiang S G, Yang C H, Zhou F L, et al. Breeding Technology of Penaeus monodon [M]. Beijing: Ocean Press. 2013, 80—86
江世贵, 杨丛海, 周发林, 等. 斑节对虾种虾繁育技术. 北京: 海洋出版社. 2013, 80—86[20] 李运东, 周发林, 黄建华, 等. 斑节对虾组织蛋白酶L基因的克隆及其表达分析. 南方水产科学, 2016, 12(3): 58—66 Li Y D, Zhou F L, Huang J H, et al. Cloning and expression analysis of Cathepsin L cDNA of Penaeus monodon [J]. South China Fisheries Science, 2016, 12(3): 58—66
李运东, 周发林, 黄建华, 等. 斑节对虾组织蛋白酶L基因的克隆及其表达分析. 南方水产科学, 2016, 12(3): 58—66[21] 郑丽明, 周发林, 杨其彬, 等. 斑节对虾金属硫蛋白基因cDNA克隆与表达分析. 水生生物学报, 2011, 35(6): 913—919 http://ssswxb.ihb.ac.cn/CN/abstract/abstract1992.shtml Zheng L M, Zhou F L, Yang Q B, et al. Molecular cloning and expression analysis of metallothionein from Penaeus monodon [J]. Acta Hydrobiologica Sinica, 2011, 35(6): 913—919
郑丽明, 周发林, 杨其彬, 等. 斑节对虾金属硫蛋白基因cDNA克隆与表达分析. 水生生物学报, 2011, 35(6): 913—919 http://ssswxb.ihb.ac.cn/CN/abstract/abstract1992.shtml[22] 唐小红, 樊佳佳, 白俊杰. 草鱼α-淀粉酶基因组织表达特征和早期发育的表达谱. 海洋渔业, 2015, 37(1): 31—37 http://kns.cnki.net/KCMS/detail/detail.aspx?filename=htyy201501005&dbname=CJFD&dbcode=CJFQ Expression analysis of α-amylase gene in various tissues and early development of Ctenopharyngodon idellus [J]. Marine Fisheries, 2015, 37(1): 31—37
唐小红, 樊佳佳, 白俊杰. 草鱼α-淀粉酶基因组织表达特征和早期发育的表达谱. 海洋渔业, 2015, 37(1): 31—37 http://kns.cnki.net/KCMS/detail/detail.aspx?filename=htyy201501005&dbname=CJFD&dbcode=CJFQ[23] 吴婷婷, 朱晓鸣. 鳜鱼、青鱼、草鱼、鲤、鲫、鲢消化酶活性的研究. 中国水产科学, 1994, 1(2): 10—17 Wu T T, Zhu X M. Studies on the activity of digestive enzymes in mandarin fish, black carp, grass carp, common carp, crucian carp and silver carp [J]. Journal of Fishery Sciences of China, 1994, 1(2): 10—17
吴婷婷, 朱晓鸣. 鳜鱼、青鱼、草鱼、鲤、鲫、鲢消化酶活性的研究. 中国水产科学, 1994, 1(2): 10—17[24] 彭涛. 克氏原螯虾α-淀粉酶基因的克隆与功能分析. 安徽农业大学, 2015 http://cdmd.cnki.com.cn/Article/CDMD-10364-1016064157.htm Peng Tao. Cloning and functional analysis of a-amylase gene in Procambarus clarkia [D]. Anhui Agricultural University, 2015
彭涛. 克氏原螯虾α-淀粉酶基因的克隆与功能分析. 安徽农业大学, 2015 http://cdmd.cnki.com.cn/Article/CDMD-10364-1016064157.htm[25] Cuvier-Peres A, Kestemont P. Development of some digestive enzymes in Eurasian perch larvae Perca fluviatilis [J]. Fish Physiology and Biochemistry, 2001, 24(4): 279—285 doi: 10.1023/A:1015033300526
[26] Darias M J, Murray H M, Gallant J W, et al. Characterization of a partial alpha-amylase clone from red porgy (Pagrus pagrus): Expression during larval development [J]. Comparative Biochemistry and Physiology B-Biochemistry & Molecular Biology, 2006, 143(2): 209—218
[27] Kim B G, Divakaran S, Brown C L, et al. Comparative digestive enzyme ontogeny in two marine larval fishes: Pacific threadfin (Polydactylus sexfilis) and bluefin trevally (Caranx melampygus) [J]. Fish Physiology and Biochemistry, 2001, 24(3): 225—241 doi: 10.1023/A:1014054431627
[28] Kurokawa T, Suzuki T, Ohta H, et al. Expression of pancreatic enzyme genes during the early larval stage of Japanese eel Anguilla japonica [J]. Fisheries Science, 2002, 68(4): 736—744 doi: 10.1046/j.1444-2906.2002.00487.x
[29] 潘鲁青, 刘泓宇, 肖国强. 甲壳动物幼体消化酶研究进展. 中国水产科学, 2006, 13(3): 492—501 Pan LQ, Liu H Y, Xiao G Q. A review on digestive enzyme of crustacean larvae [J]. Journal of Fishery Sciences of China, 2006, 13(3): 492—501
潘鲁青, 刘泓宇, 肖国强. 甲壳动物幼体消化酶研究进展. 中国水产科学, 2006, 13(3): 492—501[30] 田华梅, 王群, 赵云龙, 等. 中华绒螯蟹胚胎发育过程中的消化酶活力及氨基酸组成. 中国水产科学, 2003, 10(5): 404—408 Tian H M, Wang Q, Zhao Y L, et al. Digestive enzyme activities and amino acids composition during embryonic development of Eriocheir sinensis [J]. Journal of Fishery Sciences of China, 2003, 10(5): 404—408
田华梅, 王群, 赵云龙, 等. 中华绒螯蟹胚胎发育过程中的消化酶活力及氨基酸组成. 中国水产科学, 2003, 10(5): 404—408[31] 刘立鹤, 陈立侨, 周永奎, 等. 甲壳动物消化酶的研究. 饲料工业, 2006, 27(18): 56—62 doi: 10.3969/j.issn.1001-991X.2006.18.018 Liu L H, Chen L Q, Zhou Y K, et al. Studies on the digestive enzymes of the chitosan [J]. Feed Industry, 2006, 27(18): 56—62
刘立鹤, 陈立侨, 周永奎, 等. 甲壳动物消化酶的研究. 饲料工业, 2006, 27(18): 56—62 doi: 10.3969/j.issn.1001-991X.2006.18.018[32] Wei J, Zhang X, Yu Y, et al. Comparative transcriptomic characterization of the early development in pacific white shrimp Litopenaeus vannamei [J]. Plos One, 2014, 9(9): e106201 doi: 10.1371/journal.pone.0106201
[33] Wei J, Zhang X, Yu Y, et al. RNA-seq reveals the dynamic and diverse features of digestive enzymes during early development of pacific white shrimp Litopenaeus vannamei [J]. Comparative Biochemistry & Physiology Part D Genomics & Proteomics, 2014, 11(5): 37—44.
-
期刊类型引用(20)
1. 米玮洁,张舜,胡俊,高少波,胡菊香. 黄河内蒙古河段水文情势变化及生态效应初探. 水生态学杂志. 2024(01): 42-49 .  百度学术
百度学术
2. 班学君,樊博,刘瀚,俞立雄,林俊强,夏继红,张迪. 鱼类产卵行为与生态水文指标响应关系研究——以长江四大家鱼为例. 水生态学杂志. 2024(01): 67-74 . 百度学术
3. 李莎,姜伟,李博,党莹超,苏巍,戴凌全. 长江葛洲坝坝下江段鱼类群落结构及多样性. 水生态学杂志. 2024(03): 121-130 . 百度学术
4. 张云昌,丁洋,赵进勇. 河流伦理建构中的八个理论和实践问题探讨. 中国水利. 2024(16): 7-11 . 百度学术
5. 易燃,陶江平,杨志,刘宏高,徐薇,曹俊,张鹏. 改进的栖息地法求解多种产漂流性卵鱼类生态流量方法研究. 中国农村水利水电. 2024(09): 45-51+59 . 百度学术
6. 蒋祥龙,黎明政,杨少荣,林鹏程,常涛,王春伶,张晨,高欣. 鄱阳湖鱼类多样性的时空变化特征研究. 水生生物学报. 2023(03): 376-388 . 本站查看
7. 周月娟,高雷,段辛斌,姜伟,黄安阳,王珂,陈大庆,汪登强. 基于耳石微结构的长江监利断面草鱼和鲢仔鱼群体日龄结构研究. 长江流域资源与环境. 2023(02): 324-330 . 百度学术
8. 徐薇,金瑶,陈桂亚,董纯,易燃,赵娜,陶江平. 三峡水库十年生态调度(2011—2020年)期间下游沙市江段产漂流性卵鱼类自然繁殖变化. 湖泊科学. 2023(05): 1729-1741 . 百度学术
9. 高雷,郭国忠,杨浩,陈大庆,王珂,段辛斌. 长江中游洪湖江段四大家鱼早期资源及生态调度影响. 中国水产科学. 2023(12): 1556-1565 . 百度学术
10. 高欣,赵亚辉,田菲,王晓爱,黎明政,林鹏程,常涛,俞丹,刘焕章. 中国内陆水体鱼类多样性监测专项网的监测和研究进展. 生物多样性. 2023(12): 199-210 . 百度学术
11. 霍堂斌,宋聃,刘伟,唐富江,王继隆,张澜澜,李喆,赵荣伟. 松花江下游富锦江段鱼类早期资源状况. 中国水产科学. 2022(01): 91-101 . 百度学术
12. 彭云鑫,方弟安,徐东坡,丁隆强,黎加胜,任鹏,何晓辉,罗宇婷. 长江安庆段草鱼仔稚鱼资源时空动态特征与水文因子的相关性. 中国水产科学. 2022(02): 314-324 . 百度学术
13. 蒋祥龙,黎明政,杨少荣,林鹏程,王春伶,张晨,高欣. 鄱阳湖鱼类集合群落结构特征及其时间变化研究. 长江流域资源与环境. 2022(03): 588-601 . 百度学术
14. 李鹏程,荣义峰,杜浩,王普渊,刘文成,刁亚芹. 基于深度学习的青海湖裸鲤产卵场遥感识别方法. 中国水产科学. 2022(03): 398-407 . 百度学术
15. 王小豪,方弟安,孙海博,何苗,薛向平. 长江如皋江段仔稚鱼资源现状调查分析. 南方农业学报. 2022(06): 1742-1751 . 百度学术
16. 向浩,林枫,伍遇普,田波,蔡志宇,吴益平,黄盼君,刘千凡,周志国. 长江中游宜昌段浮游动物群落结构特征与环境因子相关性分析. 黑龙江水产. 2022(05): 3-12 . 百度学术
17. 汤正阳,蒋爱萍,林俊强,任玉峰,张迪,刘新波,杨靖,赵庆绪. 长江流域水工程生态环境保护研究进展. 水利水电技术(中英文). 2022(S2): 297-309 . 百度学术
18. 祁涛,班璇,杜浩,郭文献,杜耘,龙奥运,南璐艺,石小涛,郑楚洋. 长江中游鱼类敏感水文期内水文情势对其资源量的影响评估. 长江流域资源与环境. 2022(12): 2621-2632 . 百度学术
19. 胡春明,董熙,尤立,李曜. 红花尔基水利枢纽对下游河道水文情势影响研究. 干旱区资源与环境. 2021(06): 89-95 . 百度学术
20. 王慧,张建新,田翊霏,陈君,耿硕,杨元昊,王立新. 湑水河国家级水产种质资源保护区鱼类资源调查与评价. 家畜生态学报. 2021(11): 54-59 . 百度学术
其他类型引用(6)





计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 26