
SPECIES DELIMITATION AND PHYLOGENETIC RELATIONSHIPS OFPSEUDOGOBIO FISHES IN THE SUBFAMILY GOBIONINAE
-
摘要: 逗亚科似逗属鱼类的物种界定保持争议, 系统发育关系尚待解决。研究取样似逗属鱼类所有种, 使用核基因多位点序列重建似逗属鱼类的系统发育关系, 运用分子的物种界定方法并结合形态特征分析厘定我国似逗属鱼类的分类。贝叶斯系统发育树结果表明: 桂林似逗与平江似逗是单系种; 似逗与扁嘴似逗是多系种, 前者包括五个谱系A至E, 后者包括两个谱系A与B。POFAD距离分析和Structurama分析的结果表明似逗和扁嘴似逗的每个谱系是独立遗传种群, BP & P分析结果强烈支持它们是不同种。*Beast物种树结果揭示: 扁嘴似逗谱系B位于似逗属鱼类的基部位置; 似逗谱系A与B是姊妹群关系, 似逗谱系C是扁嘴似逗谱系A的姊妹群, 它们一起与桂林似逗形成姊妹群关系; 似逗谱系D与E是姊妹群关系, 它们一起是平江似逗的姊妹群。结合形态证据, 对我国似逗属鱼类分类厘定如下: 限定严格意义似逗包括似逗谱系A+B; 恢复长吻似逗(Pseudogobio longirostris Mori, 1934)给予似逗谱系C分类名; 似逗谱系D与E是隐存种, 桂林似逗与平江似逗是有效种。Abstract: Species delimitation and phylogenetic relationships of the genus Pseudogobio fishes in the subfamily Gobio-ninae remain debated and unresolved. This study used molecular species delimitation methods and morphological analyses to reconstruct phylogenetic relationships of Pseudogobio fishes based on nuclear loci and the classification of this genus in China. The results of Bayesian tree showed that Pseudogobio guilinensis and P. banggiangensis were monophyletic specie, and that P. vaillanti and P. escocinus were polyphyletic species that the former included five Clades A-E and the latter was comprised of two Clades A and B. The analyses of POFAD and Structurama showed that each clade of P. vaillanti and P. escocinus were independent genetic populations, and BP & P analysis strongly supported them as different species. The results of *Beast species tree revealed that P. esocinus Clade B occupied a basal position of Pseudogobio fishes; Clades A and B of P. vaillanti were the sister taxon relationship, and P. vaillanti Clade C was the sister taxon of P. esocinus Clade A, and all of them together were the sister group of P. guilinensis; Clades D and E of P. vaillanti were the sister taxon relationship, which were the sister group of P. banggiangensis. A revised classification of Pseudogobio fishes in China by molecular and morphological evidence as follows: P. vaillanti sensu stricto was limited to P. vaillanti Clades A and B; A resurrection of P. longirostris Mori, 1934 was assigned to P. vaillanti Clade C; Clades D and E of P. vaillanti were cryptic species, and P. guilinensis and P. banggiangensis were valid species, respectively.
-
Keywords:
- Cyprinidae /
- Gobioninae /
- Pseudogobio /
- Species delimitation /
- Phylogeny
-
似逗属(Pseudogobio)鱼类(鲤形目Cypriniformes: 鲤科Cyprinidae, 逗亚科Gobioninae)是东亚地区特有的淡水小型鱼类, 主要栖息于溪流生境[1,2]。它们主要以碎屑、藻类、水草与底栖动物为饵料[2,3]。该属鱼类包括4种[4]: 扁嘴似逗[Pseudogobio esocinus (Temminck & Schlegel, 1846)], 模式地日本琵琶湖, 分布于日本本岛;似逗[Pseudogobio vail-lanti (Sauvage, 1878)] 模式地江西鄱阳湖流域, 广泛分布于我国大陆沿海各水系; 桂林似逗(Pseudogobio guilinensis Yao & Yang, 1977), 模式地广西桂林, 分布于我国珠江流域; 平江似逗(Pseudogobio banggiangensis Nguyen & Ngo, 2001), 模式地越南平江河(流入我国称为水口河, 属于珠江流域西江水系左江), 分布于我国珠江流域。
似逗属鱼类的物种分类一直存在争议。Bănă-rescu & Nalbant[1]认为似逗属鱼类包括扁嘴似逗1种含2亚种, 即指名亚种(Pseudogobio esocinus esoci-nus)与似逗亚种(P. esocinus vaillanti)。罗云林等[5]对我国的似逗属鱼类进行了分类厘订, 认为我国似逗属鱼类包括1种含3亚种: 似逗指名亚种(Pseudogobio vaillanti vaillanti)、桂林似逗(P. vaillanti guilinensis)与长吻似逗(P. vaillanti longirostris Mori, 1934)。乐佩琦[2]认为长吻似逗亚种是似逗指名亚种的同物异名, 并把似逗指名亚种与桂林似逗都提升为种。杨秀平等[6]利用形态可量性状使用主成分分析对我国似逗属鱼类进行了形态比较, 认为桂林似逗为似逗的同物异名, 我国似逗属鱼类仅似逗单种。
基于形态特征与分子证据重建的逗亚科鱼类系统发育关系研究[7—12]均支持似逗属鱼类是一单系群, 但是似逗属鱼类种间的系统发育关系并未解析清楚。夏曦中等[13]利用线粒体细胞色素b基因, 对我国辽河、长江、钱塘江、珠江与海南岛的广义似逗进行了谱系地理研究, 结果表明广义似逗具有明显的空间遗传结构, 能分为5大谱系, 其中两个谱系分布于长江与钱塘江, 另外三个谱系各自分布于辽河、珠江与海南岛, 谱系间遗传距离达到6%—10%。基于鱼类线粒体基因的种间平均遗传距离一般为8%左右[14—16], 夏曦中等[13]的研究结果暗示我国似逗属鱼类可能不止似逗单种。Tomi-naga等[17]使用线粒体细胞色素b基因对广义扁嘴似逗进行了初步的谱系地理研究, 结果显示广义扁嘴似逗是多系群, 包括两大谱系: 一个谱系是日本本岛特有的, 另一个谱系嵌套于似逗内部。Tomina-ga等[18]进一步加强取样使用线粒体细胞色素b基因对广义扁嘴似逗进行了详细的谱系地理研究, 结果表明广义扁嘴似逗能分为三大谱系A、B与C。其中, 谱系A与分布于我国北方的似逗形成姊妹群关系; 谱系B与谱系C是姊妹群, 它们一起与分布于我国南方的似逗形成姊妹群关系。
综上所述, 似逗属鱼类的物种分类保持争议, 系统发育关系尚待进一步解决。清晰的物种界定是准确理解生物多样性空间格局形成过程的基础。最近的研究[19—21]提倡分子证据基础的物种界定方法并结合形态证据进行物种界定。因此, 本研究取样似逗属鱼类所有物种, 选择5个核基因位点作为分子标记, 使用系统发育技术重建似逗属鱼类的系统发育关系, 运用分子证据基础的物种界定方法并结合形态证据进行分类厘定, 为将来以该属鱼类作为研究对象, 理解东亚地区淡水鱼类多样性空间格局形成过程奠定基础。
1. 材料与方法
1.1 物种取样和分子标记
本研究共取样似逗属鱼类4种69尾(表 1)。其中广义似逗58尾, 来自长江、钱塘江、瓯江、灵江、闽江、韩江、鸭绿江、大洋河与辽河水系; 平江似逗1尾, 来自珠江水系; 桂林似逗4尾, 来自珠江水系; 广义扁嘴似逗6尾, 来自日本琵琶湖与云出川水系。依据Tang等[11]重建的逗亚科鱼类分子系统发育关系, 选取长蛇逗与棒花鱼作为外类群。分子标记选择5个单拷贝核基因: 外胚层神经皮层蛋白1基因(ENC1)、错配修复蛋白MutL基因3(MLH3)、错配修复蛋白MutS基因6(MSH6)、核位点重组激活基因1(RAG1)和核糖体蛋白S7基因第一内含子(rpS7Intron 1)。
表 1 种名、采样地点、个体编号及GenBank登录号Table 1. Species, sampling sites, code and GenBank accession number of all individuals采样地点Sampling locality 编号Code GenBank登录号GenBank accession number MLH3 MSH6 ENC1 RAG1 rpS7 Intron 1 广义似逗Pseudogobio vaillanti sensu lato 安徽省潜山县 (长江下游) PV-A-QS01 KX078872 KX078974 KX079148 KX078923 KX079002 安徽省潜山县 (长江下游) PV-A-QS02 KX079298 KX079221 KX079147 KX079062 KX078990 安徽省潜山县 (长江下游) PV-A-QS03 KX079297 KX079226 KX079146 KX079063 KX079000 江西省靖安县 (长江下游) PV-A-YC KX079313 KX079222 KX079152 KX079071 KX078991 江西省奉新县 (长江下游) PV-A-FX KX079312 KX079225 KX079167 KX079053 KX078992 江西省余江县 (长江下游) PV-A-YJ KX079295 KX079219 KX079150 KX079072 KX078999 江西省安福县 (长江下游) PV-A-AF KX079303 KX079212 KX079171 KX079049 KX079013 江西省新余市 (长江下游) PV-A-XX KU715610* KX079217 KX079145 KX079074 KX079005 江西省永丰县 (长江下游) PV-A-YF KX079296 KX079224 KX079151 KX079059 KX079006 江西省永新县 (长江下游) PV-A-YX KX079294 KX079223 KX079149 KX079073 KX079001 江西省遂川县 (长江下游) PV-A-JA KX079301 KX079220 KX079165 KX079055 KX079004 江西省崇仁县 (长江下游) PV-A-CR KX079302 KX079207 KX079170 KX079050 KX079007 江西省广昌县 (长江下游) PV-A-GC KX079311 KX079206 KX079166 KX079054 KX078993 浙江省桐庐县 (钱塘江) PV-A-TL01 KX079310 KX079210 KX079159 KX079064 KX078989 浙江省桐庐县 (钱塘江) PV-A-TL02 KX079292 KX079211 KX079158 KX079065 KX078987 浙江省永嘉县 (瓯江) PV-A-YoJ KX079304 KX079214 KX079155 KX079068 KX078998 浙江省仙居县 (灵江) PV-A-XJ01 KX079299 KX079208 KX079154 KX079069 KX079011 浙江省仙居县 (灵江) PV-A-XJ02 KX079309 KX079213 KX079153 KX079070 KX079012 福建省松溪县 (闽江) PV-A-SoX01 KX079291 KX079205 KX079161 KX079060 KX079008 福建省松溪县 (闽江) PV-A-SoX02 KX079293 KX079218 KX079160 KX079061 KX078994 福建省建宁县 (闽江) PV-A-JN01 KX079306 KX079209 KX079164 KX079056 KX078988 福建省建宁县 (闽江) PV-A-JN02 KX079300 KX079216 KX079163 KX079057 KX078997 福建省长汀县 (韩江) PV-A-CT01 KX079308 KX079230 KX079169 KX079051 KX079009 福建省长汀县 (韩江) PV-A-CT02 KX079307 KX079229 KX079168 KX079052 KX079010 福建省上杭县 (韩江) PV-A-ShH KX079305 KX079215 KX079162 KX079058 KX079003 福建省武平县 (韩江) PV-A-WP01 KX079290 KX079228 KX079157 KX079066 KX078995 福建省武平县 (韩江) PV-A-WP02 KX079289 KX079227 KX079156 KX079067 KX078996 陕西省勉县 (长江中游) PV-B-MX KX079271 KX079202 KX079122 KX079092 KX079023 河南省新野县 (长江中游) PV-B-XiY01 KX079270 KX079195 KX079123 KX079099 KX079025 河南省新野县 (长江中游) PV-B-XiY02 KX079273 KX079197 KX079124 KX079090 KX079026 1.2 DNA提取、PCR扩增与产物测序
从95%酒精保存的肌肉组织中采用高盐法提取基因组DNA[22]。PCR过程使用引物[23,24]见表 2, 各基因PCR扩增反应的条件如下: 94℃预变性5min; 94℃变性50s, 52℃退火50s, 72℃延伸60s; 循环40次; 72℃最后延伸8min。PCR产物纯化后在ABI 3730 DNA测序仪中使用扩增引物进行测序。
1.3 序列处理与贝叶斯树的构建
使用DAMBE ver. 5.6.7软件[25]进行蛋白质编码基因的序列对位, 使用MAFFT ver. 7.22软件[26]进行非蛋白质编码基因对位。序列对位完成后手工校正并排除具有插入与缺失的区域。使用MEGA ver. 6.06软件[27]计算基因的分子特征。蛋白质编码基因分区策略按密码子进行, 非蛋白质编码基因不分区。使用jModeltest ver. 2.1.7软件[28], 依据赤池信息准则选择每个分区的最适碱基替换模型。在Mrbayes ver. 3.2.2软件[29]下使用核基因多位点联合序列构建贝叶斯系统发育树, 主要步骤如下: 基于马尔可夫链蒙特卡罗进行5000万代运算, 取样频率为1000, 保证分离频率平均标准偏差小于0.01。运行完成后舍弃前1/4的取样树, 根据剩余的3/4取样树构建50%的多数原则一致树, 并计算每个节点的贝叶斯后验概率值。
1.4 物种界定
在POFAD ver. 1.0.7软件[30]中采用标准化矩阵计算基于核基因多位点的个体之间Genpofad距离, 并在SplitsTree4 ver. 4.14.2软件[31]中构建邻接网络图(简称POFAD距离分析)。在Structurama 2.0软件[32]中采用Dirichlet 过程进行种群数最适值的测试, 运算过程中将其设为随机变量并服从形状参数为0.1, 尺度参数为10的伽马分布; 共运行5000万代马尔可夫链蒙特卡罗, 每1000代取样一次, 舍弃前1/4的样本; 最终获得具有最高后验概率的种群数(简称Structurama分析)。依据贝叶斯系统发育树、POFAD距离分析以及Structurama分析的结果, 设定先验物种。在BP & P 3.0软件[33,34]中验证种的成立与否。采用无指导树物种界定方法计算指定物种划分方法的后验概率分布, 按theta和tau的先验分布运行3组独立分析: (1) thetaprior=1, 10, tauprior=1, 10; (2) thetaprior=2, 2000, tauprior=2, 2000; (3) thetaprior=1, 10, tauprior=2, 2000。共运行250万代, 每5代取样一次, 舍弃前1/5的样本(简称BP & P分析)。
1.5 *BEAST物种树的构建
在*BEAST ver. 2.3.0软件[35]中构建物种树, 主要步骤包括: 选择对数正态分布的松散钟模型作为分子钟模型, 树的先验模型设定为Yule过程; 运行4个独立重复, MCMC代数设置为20000万代, 取样频率为1000; 舍弃前1/4的样本后, 在Log Combiner ver. 2.3.0软件[35]中合并每个独立重复的log文档和trees文档; 在Tracer ver. 1.5软件[36]中估算合并后的log文档参数, 保证每个参数的有效取样大小大于200; 在Tree Annotator ver. 2.3.0软件[35]中获得最大谱系置信树。
表 2 分子标记扩增使用的引物Table 2. Primers for the amplification of molecular markers基因Gene 正向引物Forward primer (5′-3′) 反向引物Reverse primer (5′-3′) 来源Source ENC1 AGGCCGGTGACATGCTGGA GGAGGTGYTTCCAGGTGCTGA 本研究 MLH3 TAGACATAGAAGCCTGCAA TGGTTCRAGGCATATGTCAT 本研究 CGTGAGGGYCATTACAA CCGCCATGTTRGTGACATC MSH6 CAGACAGCCCTGTRAAAC GAGTACCCATGCTRTGGAT 本研究 AACCTTGAGATCCTTCAGAA GAAGAGTCTCCTCATGCAGTC RAG1 CAGTAYCAYAAGATGTACCG TTGTGAGCYTCCATRAACTT Kim & Bang[23] rpS7 intron 1 GCCTCTTCCCAGGCCGTC AACTCATCTGGCTTTTCGCC Chow & Hazama[24] 1.6 外部形态特征的计数与测量
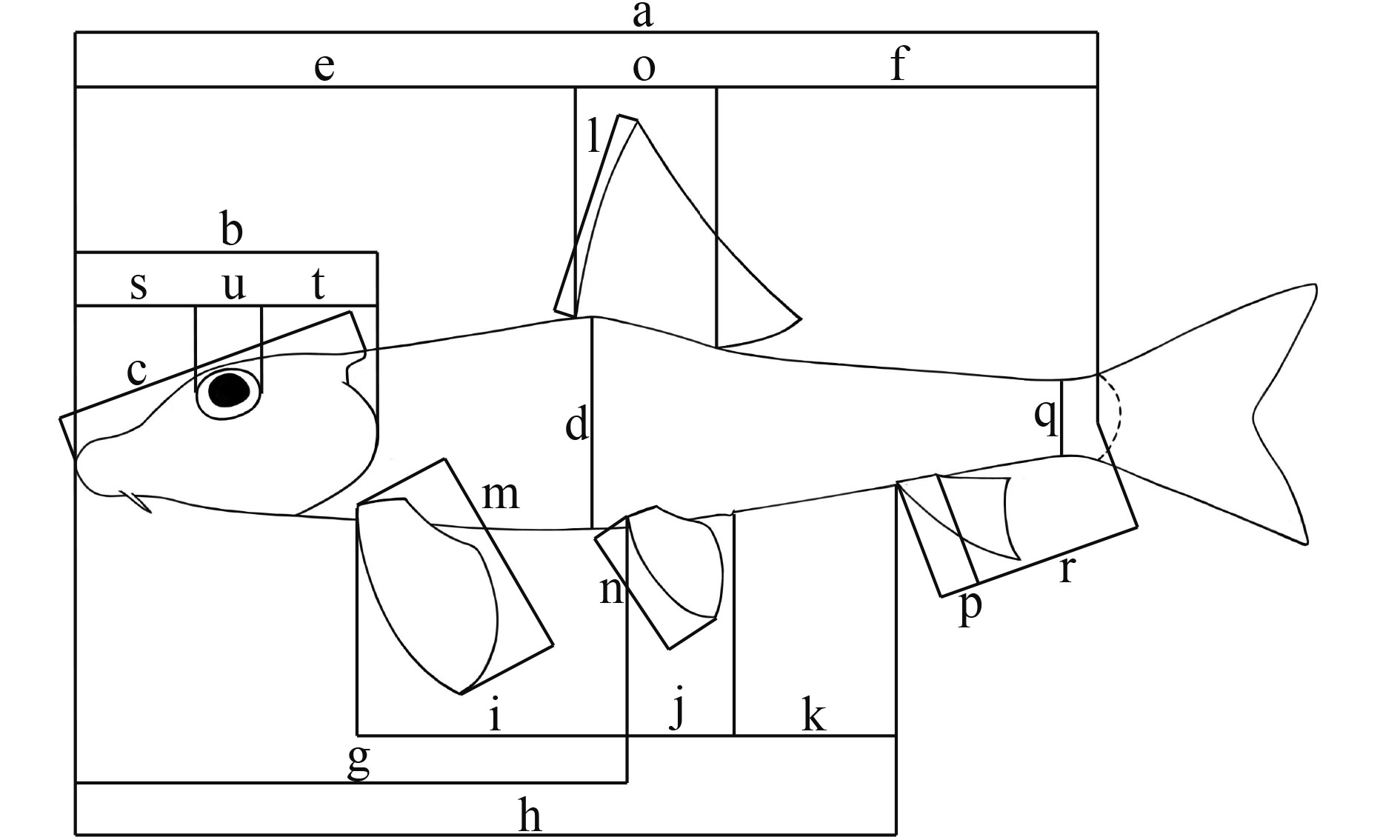
统一采用鱼体背部俯视方向左侧进行计数与测量, 可数性状依据陈宜瑜[37], 可量性状依据Kottelat和Freyhof[38](图 1)。在Past ver. 1.90[39]软件下完成可量性状数据的主成分分析。为消除异速生长的影响, 可量性状数据进行对数转换并使用Burnaby 方法进行校正[40,41]。
![]() 图 1 可量性状示意图(参考Kottelat & Jorg[38])a. 体长; b. 头长; c. 背侧头长; d. 体高; e. 背鳍前长; f. 背鳍后长; g. 腹鳍前长; h. 臀鳍前长; i. 胸腹鳍起点间距离; j. 腹鳍肛门起点间距离; k. 肛门臀鳍起点间距离; l. 背鳍长; m. 胸鳍长; n. 腹鳍长; o. 背鳍基长; p. 臀鳍基长; q. 尾柄高; r. 尾柄长; s. 吻长; t. 眼后头长; u. 眼径; v. 眼间距(图中未显示)Figure 1. The diagram of morphometric characters (referring to Kottelat & Jorg[38])a. Standard length (SL); b. Head length (HL); c. Dorsal head length; d. Body depth; e. Pre-dorsal length; f. Post-dorsal length; g. Pre-pelvic length; h. Pre-anal length; i. Distance between pectoral- and pelvic-fin origins; j. Distance between pelvic-fin and anus origins; k. Distance between anus and anal-fin origins; l. Length of dorsal fin; m. Length of pectoral fin; n. Length of pelvic fin; o. Length of dorsal-fin base; p. Length of anal-fin base; q. Depth of caudal peduncle; r. Length of caudal peduncle; s. Snout length; t. Postorbital length; u. Eye diameter; v. Eye diameter (not shown in this figure)
图 1 可量性状示意图(参考Kottelat & Jorg[38])a. 体长; b. 头长; c. 背侧头长; d. 体高; e. 背鳍前长; f. 背鳍后长; g. 腹鳍前长; h. 臀鳍前长; i. 胸腹鳍起点间距离; j. 腹鳍肛门起点间距离; k. 肛门臀鳍起点间距离; l. 背鳍长; m. 胸鳍长; n. 腹鳍长; o. 背鳍基长; p. 臀鳍基长; q. 尾柄高; r. 尾柄长; s. 吻长; t. 眼后头长; u. 眼径; v. 眼间距(图中未显示)Figure 1. The diagram of morphometric characters (referring to Kottelat & Jorg[38])a. Standard length (SL); b. Head length (HL); c. Dorsal head length; d. Body depth; e. Pre-dorsal length; f. Post-dorsal length; g. Pre-pelvic length; h. Pre-anal length; i. Distance between pectoral- and pelvic-fin origins; j. Distance between pelvic-fin and anus origins; k. Distance between anus and anal-fin origins; l. Length of dorsal fin; m. Length of pectoral fin; n. Length of pelvic fin; o. Length of dorsal-fin base; p. Length of anal-fin base; q. Depth of caudal peduncle; r. Length of caudal peduncle; s. Snout length; t. Postorbital length; u. Eye diameter; v. Eye diameter (not shown in this figure)2. 结果
2.1 序列特征
本研究使用的5个单拷贝核基因一起序列总长7582 bp, 变异位点数749, 简约信息位点数400。其中, ENC1序列长度为1290 bp, 变异位点数101, 简约信息位点数58;MLH3序列长度为2223 bp, 变异位点数230, 简约信息位点数115;MSH6序列长度为1992 bp, 变异位点数171, 简约信息位点数105;RAG1序列长度为1449 bp, 变异位点数88, 简约信息位点数47;rpS7 intron 1序列长度为628 bp, 变异位点数159, 简约信息位点数75。所有个体的GenBank登录号见表 1。
2.2 物种界定
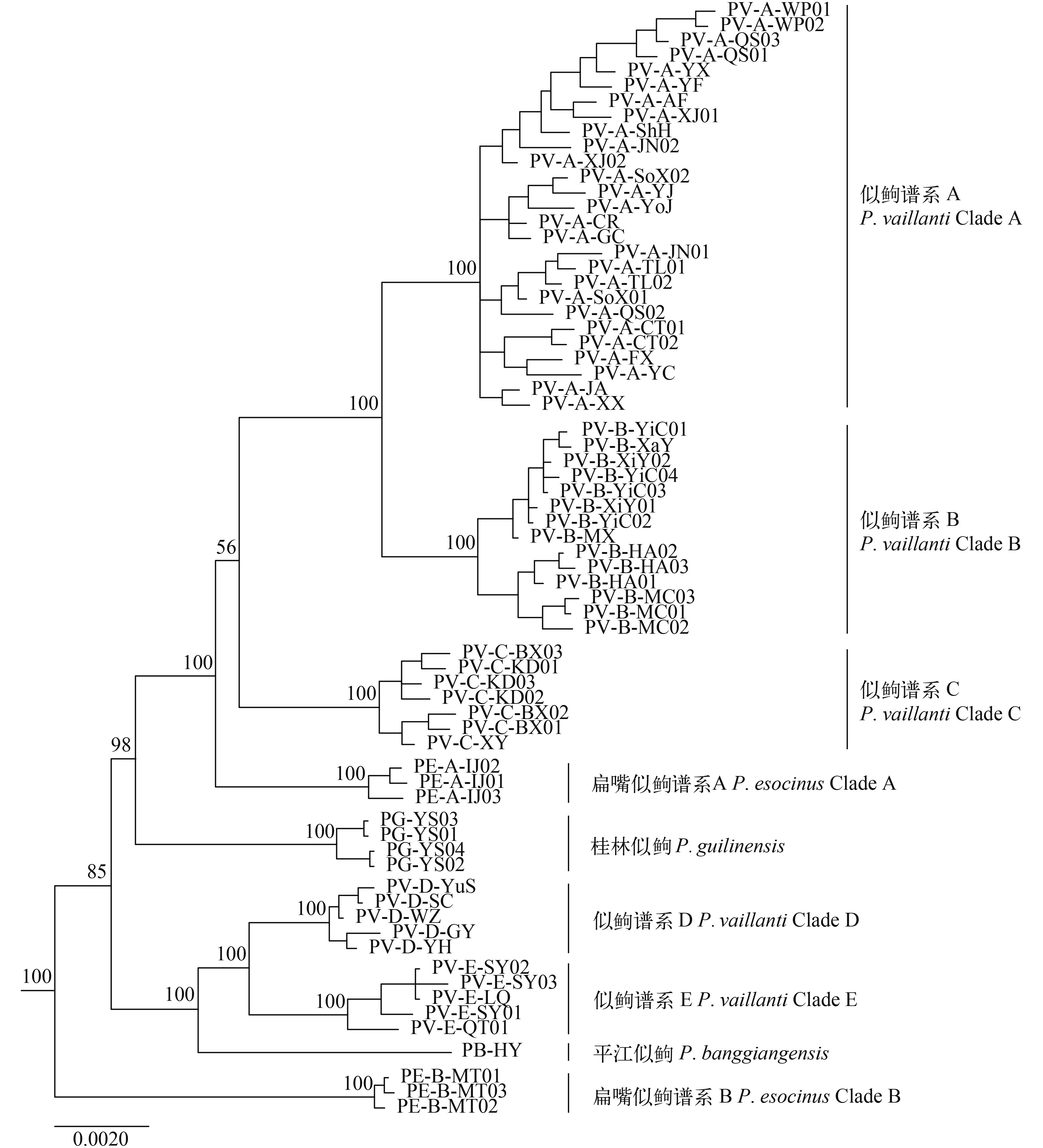
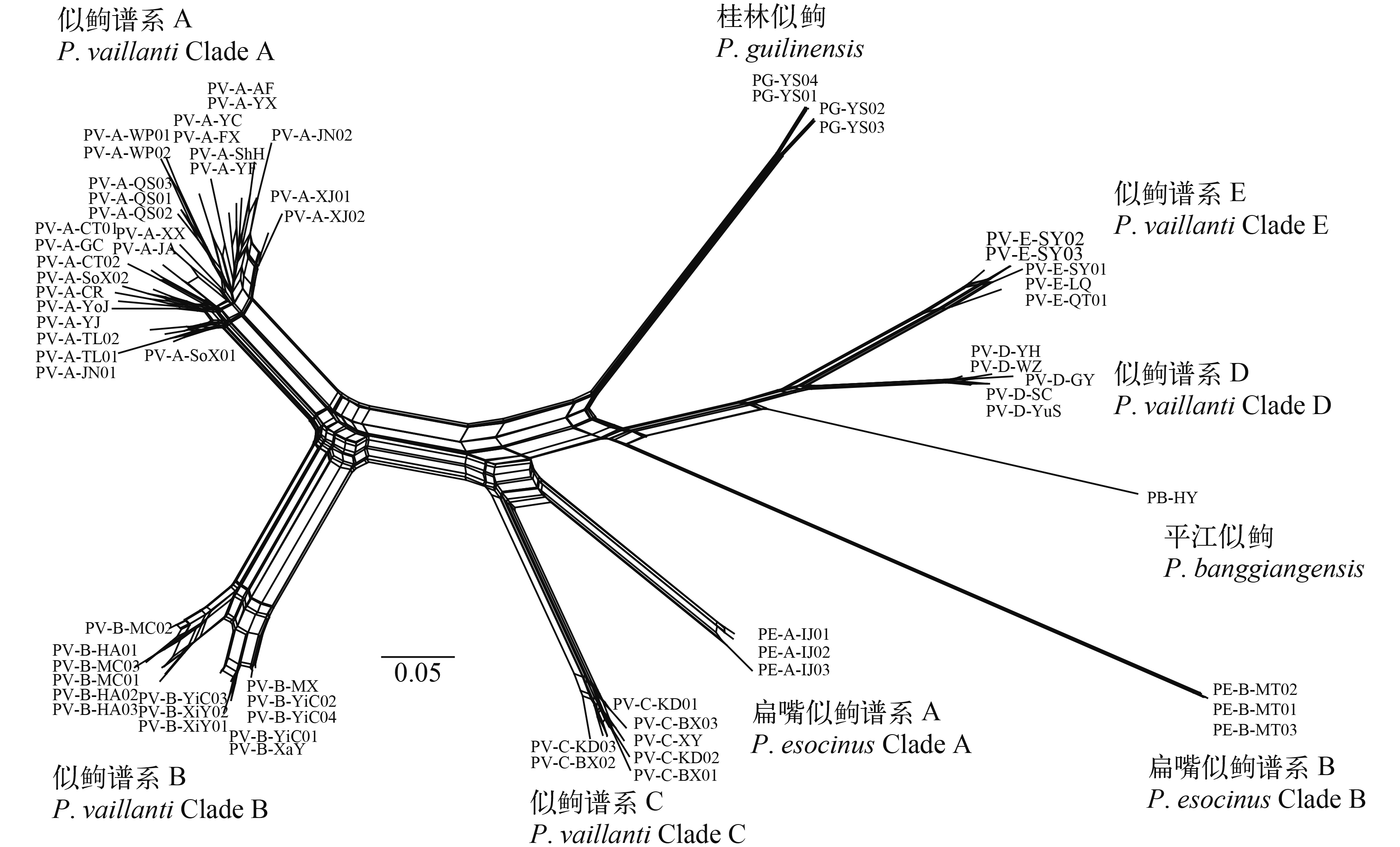
基于核基因多位点联合序列重建的贝叶斯系统发育树(图 2)揭示: 广义似逗是多系, 包括五个单系的谱系A至E; 桂林似逗与平江似逗都是单系; 广义扁嘴似逗是多系, 包括两个单系的谱系A和B。POFAD距离分析(图 3)支持桂林似逗、平江似逗、广义似逗谱系A至E与广义扁嘴似逗谱系A与B的每个谱系是独立遗传种群。Structurama分析结果与POFAD距离分析结果一致。BP & P分析结果强烈支持桂林似逗、平江似逗、广义似逗谱系A至E以及广义扁嘴似逗谱系A与B的每个谱系是不同种(贝叶斯后验概率100%)。
![]() 图 2 基于核基因多位点联合序列重建的似逗属鱼类贝叶斯系统发育树节点上方为贝叶斯后验概率, 外类群没有显示Figure 2. Bayesian tree of Pseudogobio fishes based on concatenated sequences of nuclear lociAbove nodes are Bayesian posterior probabilities, and outgroup taxa are not shown
图 2 基于核基因多位点联合序列重建的似逗属鱼类贝叶斯系统发育树节点上方为贝叶斯后验概率, 外类群没有显示Figure 2. Bayesian tree of Pseudogobio fishes based on concatenated sequences of nuclear lociAbove nodes are Bayesian posterior probabilities, and outgroup taxa are not shown![]()
2.3 系统发育关系
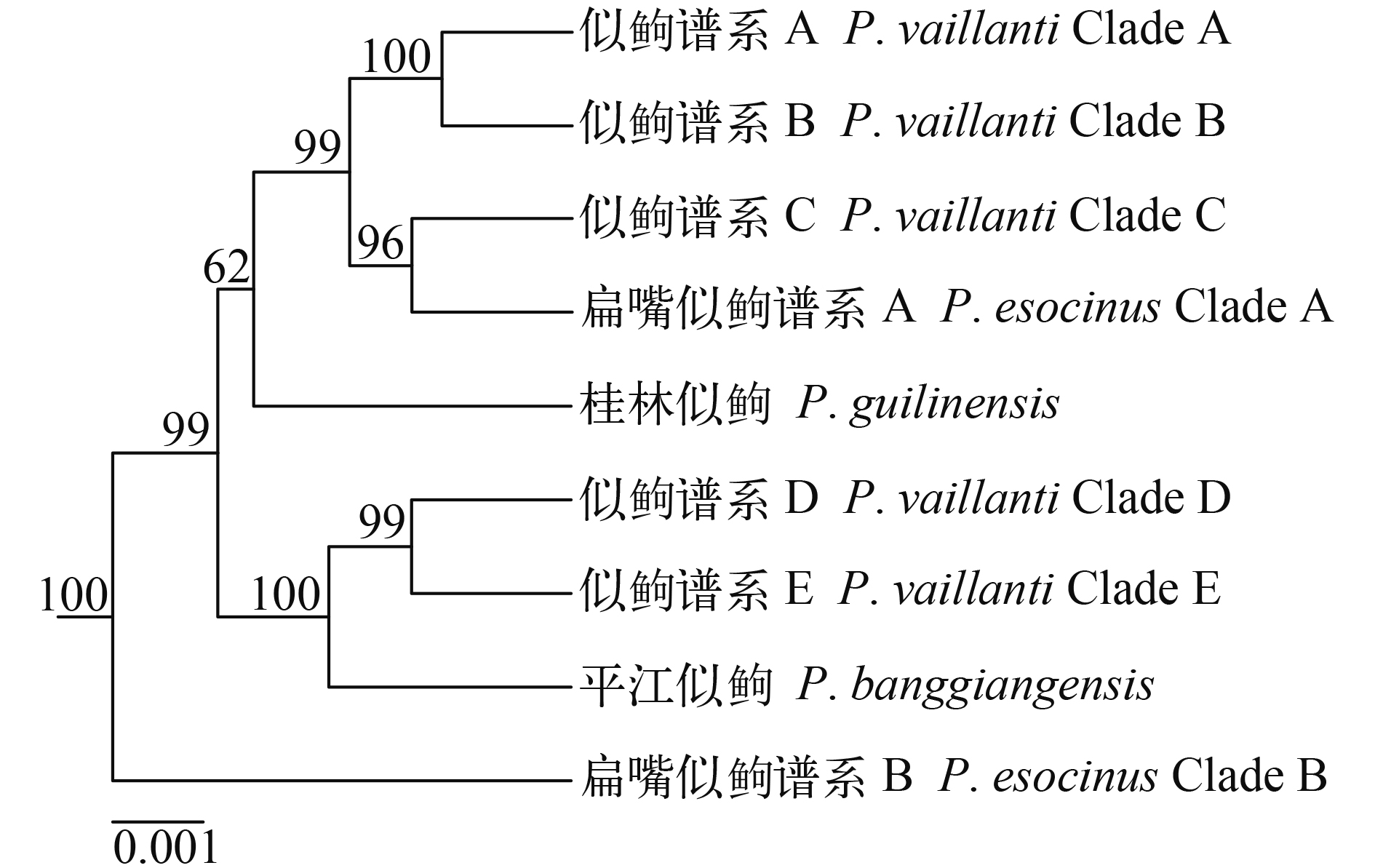
基于核基因多位点联合序列重建的贝叶斯系统发育树(图 2)与*BEAST物种树(图 4)树形基本一致。最近的鱼类分子系统发育关系研究[42—44]暗示*BEAST物种树比基于核基因多位点联合序列重建的贝叶斯系统发育树更能准确地反映系统发育关系。因此, 下面仅描述*BEAST物种树揭示的结果。
![]()
*BEAST物种树(图 4)揭示的似逗属鱼类系统发育关系为: (1)扁嘴似逗谱系B位于似逗属鱼类的基部位置, 是所有其他似逗属鱼类的姊妹群(贝叶斯后验概率100%); (2)似逗谱系A与B是姊妹群关系, 似逗谱系C是扁嘴似逗谱系A的姊妹群; (3)似逗谱系A+B与似逗谱系C+扁嘴似逗谱系A形成姊妹群关系(贝叶斯后验概率96%—100%), 它们一起是桂林似逗的姊妹群(贝叶斯后验概率62%); (4)似逗谱系D与E是姊妹群关系, 它们一起是平江似逗的姊妹群(贝叶斯后验概率99%—100%); (5)平江似逗+似逗谱系D与E是桂林似逗+扁嘴似逗谱系A+似逗谱系A至C的姊妹群(贝叶斯后验概率99%)。
2.4 形态特征
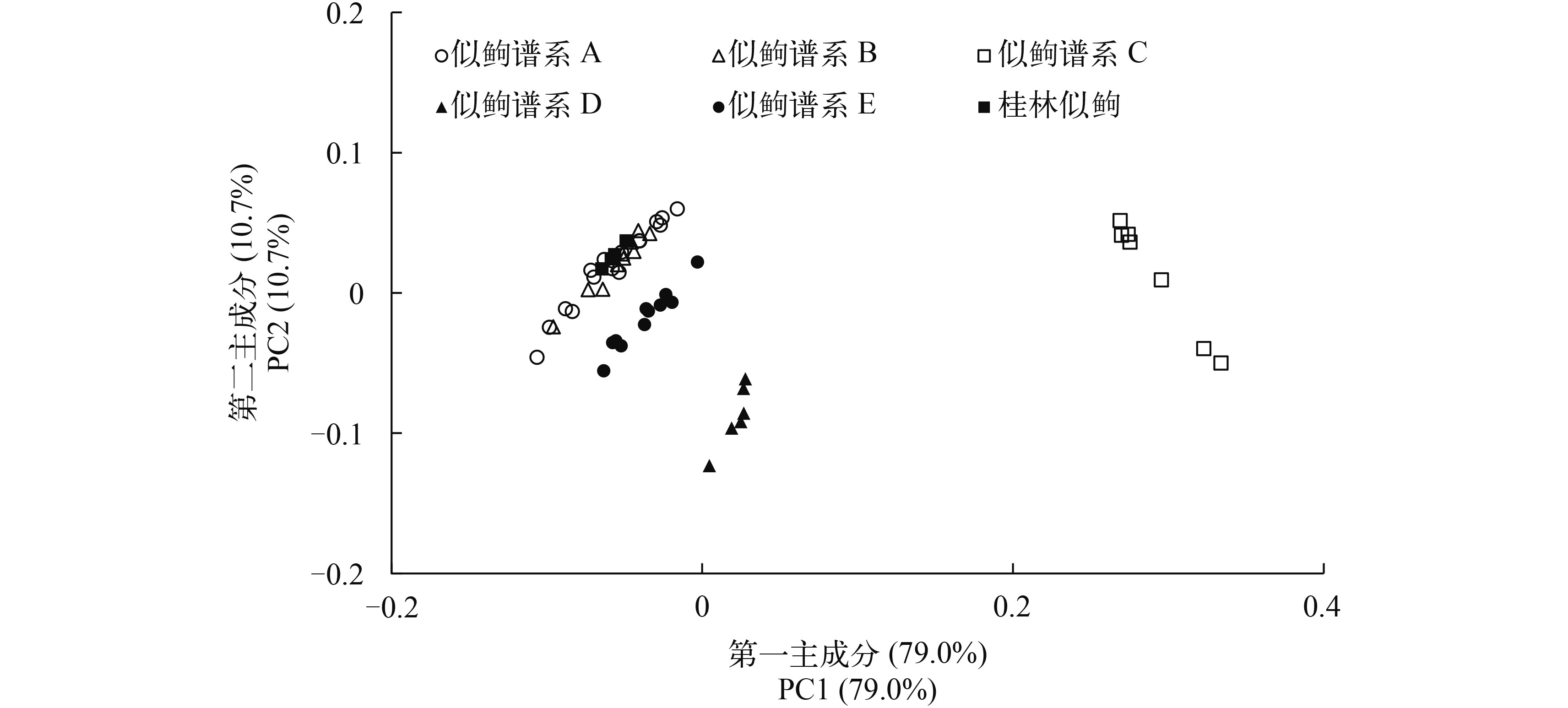
似逗、桂林似逗与平江似逗的形态特征见表 3。可量性状的主成分分析结果(图 5)表明不能分开桂林似逗、似逗谱系A与B, 但能很好地分开似逗谱系C、D与E; 桂林似逗+似逗谱系A与B能与似逗谱系C至E分开。在可数性状中(表 3), 侧线鳞的数目能够很好地分开桂林似逗(43)与似逗谱系A至E+平江似逗(40—42)。似逗谱系C侧线鳞的数目(41—42)及背鳍前鳞的数目(12—13)和似逗谱系A+B侧线鳞的数目(40—41)及背鳍前鳞的数目(10—12)也有一定差异。腹鳍分枝鳍条数与背鳍前鳞的数目能够区分平江似逗(腹鳍分枝鳍条数8; 背鳍前鳞的数目15)和桂林似逗+似逗谱系A至E(腹鳍分枝鳍条数7; 背鳍前鳞的数目10—13)。似逗谱系A与B比较、似逗谱系D与E比较, 在可数性状上几乎没有差异。
表 3 似鳇谱系A至E、桂林似鳇和平江似鳇的形态特征Table 3. Morphological characters of Pseudogobio vaillanti Clades A-E, Pseudogobio guilinensis and Pseudogobio banggiangensis可数性状与可量性状Morphometrics and Meristics 似逗谱系AP. vaillanti Clade A (n=12) 似逗谱系BP. vaillanti Clade B (n=10) 似逗谱系CP. vaillanti Clade C (n=7) 似逗谱系DP. vaillanti Clade D (n=5) 似逗谱系EP. vaillanti Clade E (n=5) 桂林似逗P. guilinensis (n=4) 平江似逗P. banggiangensis (n=1) 背鳍Dorsal fin iii-7 (12) iii-7 (10) iii-7 (7) iii-7 (5) iii-7 (5) iii-7 (4) iii-7 胸鳍Pectoral fin i-13 (3), i-14 (9) i-13 (4), i-14 (6) i-12 (1), i-13 (6) i-14 (4), i-15 (1) i-14 (5) i-14 (4) i-15 腹鳍Pelvic fin i-7 (12) i-7 (10) i-6 (1), i-7 (6) i-7 (5) i-7 (5) i-7 (4) i-8 臀鳍Anal fin iii-6 (12) iii-6 (10) iii-6 (7) iii-6 (5) iii-6 (5) iii-6 (4) iii-6 侧线鳞Lateral-line scales 40 (7), 41 (5) 40 (9), 41 (1) 41 (3), 42 (4) 41 (3), 42 (2) 40 (1), 41 (4) 43 (4) 41 侧线上鳞Scales between lateral line and dorsal-fin origin 5 (7), 6 (5) 6 (10) 6 (7) 6 (5) 6 (5) 6 (4) 6 至腹鳍侧线下鳞Scales between lateral line and pelvic-fin origin 3 (12) 3 (10) 3 (7) 3 (5) 3 (5) 3 (4) 3 至臀鳍侧线下鳞Scales between lateral line and anal-fin origin 4 (12) 4 (10) 4 (7) 4 (5) 4 (5) 4 (4) 4 背鳍前鳞Pre-dorsal scales 10 (2), 11 (8), 12 (2) 11 (7), 12 (3) 12 (5), 13 (2) 11 (2), 12 (3) 11 (2), 12 (3) 12 (4) 15 围尾柄鳞Circumpeduncular scales 12 (12) 12 (10) 12 (7) 12 (5) 12 (5) 12 (4) 12 体长Standard length (mm) 92.7—117.9 90.8—136.7 94.83—114.94 97.2—139.7 96.0—119.5 86.4—120.7 113.13 体长的百分比(%) 头长Head length 24.2—29.2 25.8—31.0 25.2—29.3 27.1—28.4 24.9—28.5 26.6—30.1 28.1 背侧头长Dorsal head length 22.1—25.9 23.3—26.8 22.7—26.6 24.1—26.4 22.4—26.1 24.2—27.8 24.6 体高Body depth 16.0—21.4 17.5—21.0 16.0—19.3 17.9—19.6 17.1—20.4 16.2—22.0 19.3 背鳍前长Pre-dorsal length 41.4—46.8 42.7—46.4 43.2—47.3 44.6—47.2 42.7—46.2 44.4—48.2 47.9 背鳍后长Post-dorsal length 39.7—46.1 41.0—45.1 40.2—44.9 40.1—42.9 40.7—43.9 38.9—43.1 37.5 腹鳍前长Pre-pelvic length 46.7—53.6 47.5—52.6 47.3—50.6 47.2—50.3 46.3—48.6 48.6—52.7 51.0 臀鳍前长Pre-anal length 76.2—82.6 77.5—82.1 79.0—83.9 76.6—82.8 76.6—79.5 78.1—82.3 83.0 胸腹鳍起点间距离Distance between pectoral- and pelvic-fin origins 21.1—27.2 20.4—24.3 20.9—24.7 21.5—25 19.5—23.0 21.7—25.1 24.4 腹鳍肛门起点间距离Distance between pelvic-fin and anus origins 5.9—7.4 6.4—8.3 7.0—8.2 5.6—7.4 6.5—7.6 6.3—6.8 7.3 肛门臀鳍起点间距离Distance between anus and anal-fin origins 18.0—27.0 20.1—25.4 21.1—25.4 22.8—25.4 22.1—24.4 21.2—25.0 24.7 背鳍长Length of dorsal fin 20.0—23.6 19.7—23.1 21.5—22.7 19.8—23.7 19.8—22.6 21.1—22.1 22.1 胸鳍长Length of pectoral fin 19.2—22.8 20.2—23.8 21.4—23.2 21.8—26 20.1—21.1 20.4—24.0 21.6 腹鳍长Length of pelvic fin 15.4—18.5 15.2—18.7 16.5—18.4 16.3—20 16.2—17.9 17.2—18.1 18.0 背鳍基长Length of dorsal-fin base 11.5—13.5 11.8—13.7 11.4—13.1 11.5—12.7 12.3—13.9 12.0—13.2 14.5 臀鳍基长Length of anal-fin base 6.5—8.6 6.0—7.1 6.2—7.0 6.1—7.2 6.5—7.2 6.2—7.1 6.6 尾柄高Depth of caudal peduncle 6.9—8.3 6.8—7.5 6.9—7.6 6.5—7.2 7.0—7.6 6.4—6.9 7.4 尾柄长Length of caudal peduncle 11.9—16.5 13.6—15.6 12.7—16.4 12.3—14.3 12.7—16.1 14.1—16.2 11.4 头长的百分比(%) 吻长Snout length 40.6—51.9 45.5—50.5 45.7—52.0 48.1—51.2 46.4—49.7 47.2—51.0 43.9 眼后头长Postorbital length 25.5—35.1 26.8—33.7 25.1—30.6 23.1—30.5 27.2—29.4 27.4—30.8 31.8 眼间距Interorbital width 14.7—21.2 18.4—23.1 15.9—18.7 19.0—20.2 18.3—21.6 18.4—20.1 18.6 眼径Eye diameter 20.9—26.2 20.4—23.2 22.0—24.8 21.3—25.7 22.4—25.4 21.2—23.6 24.3 注: 显示可数性状的记数(括号中是个体数)和可量性状的测量范围, 似逗谱系定义依据图 2 Note: The count of meristic characters (individual numbers in bracket) and the range of morphometric characters were shown. Pseudogobio vaillanti Clades defined as inFig. 2 ![]() 图 5 似逗谱系A至E和桂林似逗的第一与第二主成分得分散点图Figure 5. Scatter plots of scores on the first and second principal components for Pseudogobio vaillanti Clades A-E and P. guilinensis
图 5 似逗谱系A至E和桂林似逗的第一与第二主成分得分散点图Figure 5. Scatter plots of scores on the first and second principal components for Pseudogobio vaillanti Clades A-E and P. guilinensis3. 讨论
3.1 我国似鳇属鱼类的分类厘定
杨秀平等[6]基于形态可量性状的主成分分析结果认为我国似逗属鱼类仅似逗一种。然而, 本研究基于分子的物种界定分析结果支持桂林似逗与平江似逗是有效种, 并认为广义似逗谱系A至E以及广义扁嘴似逗谱系A与B的每个谱系是不同种。由于扁嘴似逗谱系A与B缺乏形态数据, 它们的分类有待进一步的研究再确定。下面结合形态证据就我国似逗属鱼类的分类进行讨论。
尽管分子证据支持分布于韩江至长江的似逗谱系A与分布于长江水系的似逗谱系B为不同种, 然而可数性状与可量性状分析结果不支持把似逗谱系A与B划分为不同种。另外, 似逗谱系A与B是姊妹群关系, 两者在长江水系均有分布。因此, 从形态证据、系统发育关系与地理分布三方面结合来看, 似逗谱系A与B应属于同一物种的不同遗传谱系。基于似逗谱系A+B分布范围包括似逗模式地长江水系以及其形态性状符合Sauvage[45]对似逗模式种的形态描述, 本研究限定似逗谱系A+B为严格意义似逗。
本研究结果表明分布于我国北方辽河、大洋河与鸭绿江水系的似逗谱系C与分布于日本的扁嘴似逗谱系A形成姊妹群关系, 它们一起是严格意义似逗(= 似逗谱系A+B)的姊妹群(图 4), 暗示似逗谱系C是不同种。分子的物种界定分析结果支持似逗谱系C作为种成立。可量性状分析结果能很好地区分似逗谱系C与严格意义似逗。在可数性状中, 似逗谱系C侧线鳞的数目(41—42)及背鳍前鳞的数目(12—13)与严格意义似逗侧线鳞的数目(40—41)及背鳍前鳞的数目(10—12)也有一定差异。依据罗云林等[5]对我国似逗属鱼类的分类厘订, 似逗谱系C为他们描述的长吻似逗亚种。因此, 结合分子证据、形态证据、系统发育关系与地理分布, 本研究恢复长吻似逗(Pseudogobio longi-rostrisMori, 1934, 模式地辽河水系辽宁凌源)给予似逗谱系C分类名。本研究系统发育关系结果亦表明严格意义似逗(=似逗谱系A+B)、长吻似逗(=似逗谱系C)和似逗谱系D+E均不是姊妹群关系, 暗示似逗谱系D+E属于不同种。分子证据和形态证据进一步确认分布于长江的似逗谱系D与分布于瓯江的似逗谱系E各自是不同种, 并支持平江似逗是有效种, 它们一起形成一单系群。
杨秀平等[6]基于形态可量性状的主成分分析结果认为桂林似逗是似逗的同物异名。本研究基于形态可量性状的主成分分析结果亦不能分开桂林似逗和严格意义似逗, 但它们一起能很好地与长吻似逗、似逗谱系D及E分开。然而, 系统发育关系结果表明桂林似逗与严格意义似逗+长吻似逗+扁嘴似逗谱系A形成姊妹群关系, 暗示桂林似逗是有效种。分子的物种界定分析结果确认桂林似逗是有效种。侧线鳞的数目亦能够很好地区分桂林似逗(43)和我国分布的似逗属鱼类其他种(40—42)。因此, 结合分子证据、形态证据与系统发育关系, 本研究支持桂林似逗是有效种。
3.2 似鳇属鱼类的系统发育关系与生物地理过程推测
夏曦中等[13]采纳杨秀平等[6]基于形态可量性状的主成分分析结果认为我国似逗属鱼类仅似逗单种, 并利用线粒体细胞色素b基因开展了我国广义似逗的谱系地理研究。随后Tominaga等[17,18]使用线粒体细胞色素b基因对分布于日本的扁嘴似逗进行了谱系地理研究。他们的研究均发现似逗属鱼类包括多个遗传谱系, 但都没有清晰解决似逗属鱼类谱系间的系统发育关系。本研究利用核基因多位点数据首次清楚地解析了似逗属鱼类的系统发育关系, 揭示平江似逗是似逗谱系D+E的姊妹群; 桂林似逗是严格意义似逗(=似逗谱系A+B)+长吻似逗(=似逗谱系C)+扁嘴似逗谱系A的姊妹群; 所有谱系一起与扁嘴似逗谱系B形成姊妹群关系。依据Tominaga等[18]的广义扁嘴似逗谱系地理研究, 扁嘴似逗谱系A与B均为日本本岛特有。依据本研究对我国似逗属鱼类的分类厘定, 平江似逗与桂林似逗分布于珠江水系, 严格意义似逗分布于韩江至长江的水系, 长吻似逗分布于辽河、大洋河与鸭绿江水系, 似逗谱系D分布于长江, 似逗谱系E分布于瓯江。结合上述系统发育关系与谱系的地理分布, 本研究提出以下生物地理过程是似逗属鱼类物种分化的可能原因: (1)扁嘴似逗谱系B位于系统发育树的基部位置, 暗示似逗属鱼类可能起源于东亚北部地区, 扁嘴似逗谱系B的物种形成可能是地理隔离造成; (2)桂林似逗、严格意义似逗、长吻似逗与扁嘴似逗谱系A一起构成一单系群, 四者地理分布不重叠, 推测四者的物种形成可能是地理隔离造成。同时, 扁嘴似逗谱系A是长吻似逗的姊妹群, 嵌套在似逗属鱼类内部, 暗示扁嘴似逗谱系A的物种形成亦可能是扩散过程造成, 即从大陆扩散到日本后的地理隔离; (3)平江似逗、似逗谱系D与E一起构成一单系群, 三者地理分布不重叠, 推测三者的物种形成可能是地理隔离造成; (4)平江似逗与桂林似逗均分布于珠江水系, 且两者不是姊妹群, 其系统位置均嵌套在似逗属鱼类内部, 推测平江似逗与桂林似逗的物种形成亦可能是扩散过程造成, 即似逗属鱼类祖先通过两次扩散进入珠江水系, 随后的地理隔离造成。
综上所述, 本研究清楚地对我国似逗属鱼类进行了物种界定, 解析了似逗属鱼类的系统发育关系。进一步的研究应基于动物命名法则给予似逗谱系D与E相应的分类名称, 结合形态数据厘清广义扁嘴似逗的分类。同时, 运用分子钟技术与祖先重建技术测试本研究提出的似逗属鱼类物种分化的生物地理过程, 为理解东亚地区淡水鱼类多样性空间格局形成过程提供案例。
-
![]()
图 1 可量性状示意图(参考Kottelat & Jorg[38])
a. 体长; b. 头长; c. 背侧头长; d. 体高; e. 背鳍前长; f. 背鳍后长; g. 腹鳍前长; h. 臀鳍前长; i. 胸腹鳍起点间距离; j. 腹鳍肛门起点间距离; k. 肛门臀鳍起点间距离; l. 背鳍长; m. 胸鳍长; n. 腹鳍长; o. 背鳍基长; p. 臀鳍基长; q. 尾柄高; r. 尾柄长; s. 吻长; t. 眼后头长; u. 眼径; v. 眼间距(图中未显示)
Figure 1. The diagram of morphometric characters (referring to Kottelat & Jorg[38])
a. Standard length (SL); b. Head length (HL); c. Dorsal head length; d. Body depth; e. Pre-dorsal length; f. Post-dorsal length; g. Pre-pelvic length; h. Pre-anal length; i. Distance between pectoral- and pelvic-fin origins; j. Distance between pelvic-fin and anus origins; k. Distance between anus and anal-fin origins; l. Length of dorsal fin; m. Length of pectoral fin; n. Length of pelvic fin; o. Length of dorsal-fin base; p. Length of anal-fin base; q. Depth of caudal peduncle; r. Length of caudal peduncle; s. Snout length; t. Postorbital length; u. Eye diameter; v. Eye diameter (not shown in this figure)
![]()
图 2 基于核基因多位点联合序列重建的似逗属鱼类贝叶斯系统发育树
节点上方为贝叶斯后验概率, 外类群没有显示
Figure 2. Bayesian tree of Pseudogobio fishes based on concatenated sequences of nuclear loci
Above nodes are Bayesian posterior probabilities, and outgroup taxa are not shown
![]()
![]()
![]()
图 5 似逗谱系A至E和桂林似逗的第一与第二主成分得分散点图
Figure 5. Scatter plots of scores on the first and second principal components for Pseudogobio vaillanti Clades A-E and P. guilinensis
表 1 种名、采样地点、个体编号及GenBank登录号
Table 1 Species, sampling sites, code and GenBank accession number of all individuals
采样地点Sampling locality 编号Code GenBank登录号GenBank accession number MLH3 MSH6 ENC1 RAG1 rpS7 Intron 1 广义似逗Pseudogobio vaillanti sensu lato 安徽省潜山县 (长江下游) PV-A-QS01 KX078872 KX078974 KX079148 KX078923 KX079002 安徽省潜山县 (长江下游) PV-A-QS02 KX079298 KX079221 KX079147 KX079062 KX078990 安徽省潜山县 (长江下游) PV-A-QS03 KX079297 KX079226 KX079146 KX079063 KX079000 江西省靖安县 (长江下游) PV-A-YC KX079313 KX079222 KX079152 KX079071 KX078991 江西省奉新县 (长江下游) PV-A-FX KX079312 KX079225 KX079167 KX079053 KX078992 江西省余江县 (长江下游) PV-A-YJ KX079295 KX079219 KX079150 KX079072 KX078999 江西省安福县 (长江下游) PV-A-AF KX079303 KX079212 KX079171 KX079049 KX079013 江西省新余市 (长江下游) PV-A-XX KU715610* KX079217 KX079145 KX079074 KX079005 江西省永丰县 (长江下游) PV-A-YF KX079296 KX079224 KX079151 KX079059 KX079006 江西省永新县 (长江下游) PV-A-YX KX079294 KX079223 KX079149 KX079073 KX079001 江西省遂川县 (长江下游) PV-A-JA KX079301 KX079220 KX079165 KX079055 KX079004 江西省崇仁县 (长江下游) PV-A-CR KX079302 KX079207 KX079170 KX079050 KX079007 江西省广昌县 (长江下游) PV-A-GC KX079311 KX079206 KX079166 KX079054 KX078993 浙江省桐庐县 (钱塘江) PV-A-TL01 KX079310 KX079210 KX079159 KX079064 KX078989 浙江省桐庐县 (钱塘江) PV-A-TL02 KX079292 KX079211 KX079158 KX079065 KX078987 浙江省永嘉县 (瓯江) PV-A-YoJ KX079304 KX079214 KX079155 KX079068 KX078998 浙江省仙居县 (灵江) PV-A-XJ01 KX079299 KX079208 KX079154 KX079069 KX079011 浙江省仙居县 (灵江) PV-A-XJ02 KX079309 KX079213 KX079153 KX079070 KX079012 福建省松溪县 (闽江) PV-A-SoX01 KX079291 KX079205 KX079161 KX079060 KX079008 福建省松溪县 (闽江) PV-A-SoX02 KX079293 KX079218 KX079160 KX079061 KX078994 福建省建宁县 (闽江) PV-A-JN01 KX079306 KX079209 KX079164 KX079056 KX078988 福建省建宁县 (闽江) PV-A-JN02 KX079300 KX079216 KX079163 KX079057 KX078997 福建省长汀县 (韩江) PV-A-CT01 KX079308 KX079230 KX079169 KX079051 KX079009 福建省长汀县 (韩江) PV-A-CT02 KX079307 KX079229 KX079168 KX079052 KX079010 福建省上杭县 (韩江) PV-A-ShH KX079305 KX079215 KX079162 KX079058 KX079003 福建省武平县 (韩江) PV-A-WP01 KX079290 KX079228 KX079157 KX079066 KX078995 福建省武平县 (韩江) PV-A-WP02 KX079289 KX079227 KX079156 KX079067 KX078996 陕西省勉县 (长江中游) PV-B-MX KX079271 KX079202 KX079122 KX079092 KX079023 河南省新野县 (长江中游) PV-B-XiY01 KX079270 KX079195 KX079123 KX079099 KX079025 河南省新野县 (长江中游) PV-B-XiY02 KX079273 KX079197 KX079124 KX079090 KX079026  下载: 导出CSV
下载: 导出CSV
表 2 分子标记扩增使用的引物
Table 2 Primers for the amplification of molecular markers
基因Gene 正向引物Forward primer (5′-3′) 反向引物Reverse primer (5′-3′) 来源Source ENC1 AGGCCGGTGACATGCTGGA GGAGGTGYTTCCAGGTGCTGA 本研究 MLH3 TAGACATAGAAGCCTGCAA TGGTTCRAGGCATATGTCAT 本研究 CGTGAGGGYCATTACAA CCGCCATGTTRGTGACATC MSH6 CAGACAGCCCTGTRAAAC GAGTACCCATGCTRTGGAT 本研究 AACCTTGAGATCCTTCAGAA GAAGAGTCTCCTCATGCAGTC RAG1 CAGTAYCAYAAGATGTACCG TTGTGAGCYTCCATRAACTT Kim & Bang[23] rpS7 intron 1 GCCTCTTCCCAGGCCGTC AACTCATCTGGCTTTTCGCC Chow & Hazama[24]
下载: 导出CSV
表 3 似鳇谱系A至E、桂林似鳇和平江似鳇的形态特征
Table 3 Morphological characters of Pseudogobio vaillanti Clades A-E, Pseudogobio guilinensis and Pseudogobio banggiangensis
可数性状与可量性状Morphometrics and Meristics 似逗谱系AP. vaillanti Clade A (n=12) 似逗谱系BP. vaillanti Clade B (n=10) 似逗谱系CP. vaillanti Clade C (n=7) 似逗谱系DP. vaillanti Clade D (n=5) 似逗谱系EP. vaillanti Clade E (n=5) 桂林似逗P. guilinensis (n=4) 平江似逗P. banggiangensis (n=1) 背鳍Dorsal fin iii-7 (12) iii-7 (10) iii-7 (7) iii-7 (5) iii-7 (5) iii-7 (4) iii-7 胸鳍Pectoral fin i-13 (3), i-14 (9) i-13 (4), i-14 (6) i-12 (1), i-13 (6) i-14 (4), i-15 (1) i-14 (5) i-14 (4) i-15 腹鳍Pelvic fin i-7 (12) i-7 (10) i-6 (1), i-7 (6) i-7 (5) i-7 (5) i-7 (4) i-8 臀鳍Anal fin iii-6 (12) iii-6 (10) iii-6 (7) iii-6 (5) iii-6 (5) iii-6 (4) iii-6 侧线鳞Lateral-line scales 40 (7), 41 (5) 40 (9), 41 (1) 41 (3), 42 (4) 41 (3), 42 (2) 40 (1), 41 (4) 43 (4) 41 侧线上鳞Scales between lateral line and dorsal-fin origin 5 (7), 6 (5) 6 (10) 6 (7) 6 (5) 6 (5) 6 (4) 6 至腹鳍侧线下鳞Scales between lateral line and pelvic-fin origin 3 (12) 3 (10) 3 (7) 3 (5) 3 (5) 3 (4) 3 至臀鳍侧线下鳞Scales between lateral line and anal-fin origin 4 (12) 4 (10) 4 (7) 4 (5) 4 (5) 4 (4) 4 背鳍前鳞Pre-dorsal scales 10 (2), 11 (8), 12 (2) 11 (7), 12 (3) 12 (5), 13 (2) 11 (2), 12 (3) 11 (2), 12 (3) 12 (4) 15 围尾柄鳞Circumpeduncular scales 12 (12) 12 (10) 12 (7) 12 (5) 12 (5) 12 (4) 12 体长Standard length (mm) 92.7—117.9 90.8—136.7 94.83—114.94 97.2—139.7 96.0—119.5 86.4—120.7 113.13 体长的百分比(%) 头长Head length 24.2—29.2 25.8—31.0 25.2—29.3 27.1—28.4 24.9—28.5 26.6—30.1 28.1 背侧头长Dorsal head length 22.1—25.9 23.3—26.8 22.7—26.6 24.1—26.4 22.4—26.1 24.2—27.8 24.6 体高Body depth 16.0—21.4 17.5—21.0 16.0—19.3 17.9—19.6 17.1—20.4 16.2—22.0 19.3 背鳍前长Pre-dorsal length 41.4—46.8 42.7—46.4 43.2—47.3 44.6—47.2 42.7—46.2 44.4—48.2 47.9 背鳍后长Post-dorsal length 39.7—46.1 41.0—45.1 40.2—44.9 40.1—42.9 40.7—43.9 38.9—43.1 37.5 腹鳍前长Pre-pelvic length 46.7—53.6 47.5—52.6 47.3—50.6 47.2—50.3 46.3—48.6 48.6—52.7 51.0 臀鳍前长Pre-anal length 76.2—82.6 77.5—82.1 79.0—83.9 76.6—82.8 76.6—79.5 78.1—82.3 83.0 胸腹鳍起点间距离Distance between pectoral- and pelvic-fin origins 21.1—27.2 20.4—24.3 20.9—24.7 21.5—25 19.5—23.0 21.7—25.1 24.4 腹鳍肛门起点间距离Distance between pelvic-fin and anus origins 5.9—7.4 6.4—8.3 7.0—8.2 5.6—7.4 6.5—7.6 6.3—6.8 7.3 肛门臀鳍起点间距离Distance between anus and anal-fin origins 18.0—27.0 20.1—25.4 21.1—25.4 22.8—25.4 22.1—24.4 21.2—25.0 24.7 背鳍长Length of dorsal fin 20.0—23.6 19.7—23.1 21.5—22.7 19.8—23.7 19.8—22.6 21.1—22.1 22.1 胸鳍长Length of pectoral fin 19.2—22.8 20.2—23.8 21.4—23.2 21.8—26 20.1—21.1 20.4—24.0 21.6 腹鳍长Length of pelvic fin 15.4—18.5 15.2—18.7 16.5—18.4 16.3—20 16.2—17.9 17.2—18.1 18.0 背鳍基长Length of dorsal-fin base 11.5—13.5 11.8—13.7 11.4—13.1 11.5—12.7 12.3—13.9 12.0—13.2 14.5 臀鳍基长Length of anal-fin base 6.5—8.6 6.0—7.1 6.2—7.0 6.1—7.2 6.5—7.2 6.2—7.1 6.6 尾柄高Depth of caudal peduncle 6.9—8.3 6.8—7.5 6.9—7.6 6.5—7.2 7.0—7.6 6.4—6.9 7.4 尾柄长Length of caudal peduncle 11.9—16.5 13.6—15.6 12.7—16.4 12.3—14.3 12.7—16.1 14.1—16.2 11.4 头长的百分比(%) 吻长Snout length 40.6—51.9 45.5—50.5 45.7—52.0 48.1—51.2 46.4—49.7 47.2—51.0 43.9 眼后头长Postorbital length 25.5—35.1 26.8—33.7 25.1—30.6 23.1—30.5 27.2—29.4 27.4—30.8 31.8 眼间距Interorbital width 14.7—21.2 18.4—23.1 15.9—18.7 19.0—20.2 18.3—21.6 18.4—20.1 18.6 眼径Eye diameter 20.9—26.2 20.4—23.2 22.0—24.8 21.3—25.7 22.4—25.4 21.2—23.6 24.3 注: 显示可数性状的记数(括号中是个体数)和可量性状的测量范围, 似逗谱系定义依据图 2 Note: The count of meristic characters (individual numbers in bracket) and the range of morphometric characters were shown. Pseudogobio vaillanti Clades defined as inFig. 2
下载: 导出CSV
-
[1] Bǎnǎrescu P, Nalbant T T. Pisces, Teleostei, Cyprinidae (Gobioninae) [M]. Das Tierreich, Lieferung 93, Berlin: Walter de Guryter. 1973, 1—304
[2] 乐佩琦. 似逗属. 见: 陈宜瑜, 中国动物志硬骨鱼纲鲤形目(中卷). 北京: 科学出版社. 1998, 374—378 Yue P Q. Pseudogobio Bleeker, 1860 [A]. In: Chen Y Y (Eds.), Fauna Sinica, Osteichthyes, Cypriniformes (II) [C]. Beijing: Science Press. 1998, 374—378
乐佩琦. 似逗属. 见: 陈宜瑜, 中国动物志硬骨鱼纲鲤形目(中卷). 北京: 科学出版社. 1998, 374—378[3] 谢从新, 刘齐德. 似逗生物学的初步研究. 华中农业大学学报, 1986, 5(1): 73—82 http://www.cnki.com.cn/Article/CJFDTOTAL-HZNY198601012.htm Xie C X, Liu Q D. A Preliminary study on the biology of Pseudogobio vaillanti vaillanti (Sauvage) [J]. Journal of Huazhong Agricultural University, 1986, 5(1): 73—82
谢从新, 刘齐德. 似逗生物学的初步研究. 华中农业大学学报, 1986, 5(1): 73—82 http://www.cnki.com.cn/Article/CJFDTOTAL-HZNY198601012.htm[4] Eschmeyer W N, Fricke R, Van der Laan R. Catalog of Fishes: Genera, Species, References, http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
[5] 罗云林, 乐佩琦, 陈宜瑜. 似逗属. 见: 伍献文, 中国鲤科鱼类志(下卷). 上海: 上海人民出版社. 1977, 512—516 Luo Y L, Yue P Q, Chen Y Y. Pseudogobio Bleeker, 1860 [A]. In: Wu X W (Eds.), Fauna of Cyprinidae in China (II) [C]. Shanghai: Shanghai People’s Publishing House. 1977, 512—516
罗云林, 乐佩琦, 陈宜瑜. 似逗属. 见: 伍献文, 中国鲤科鱼类志(下卷). 上海: 上海人民出版社. 1977, 512—516[6] 杨秀平, 张敏莹, 刘焕章. 中国似逗属鱼类的形态变异及地理分化研究. 水生生物学报, 2002, 26(3): 281—285 http://www.cnki.com.cn/Article/CJFDTOTAL-SSWX200203012.htm Yang X P, Zhang M Y, Liu H Z. Morphological varia-tions and geographical differentiation of the genus Pseudogobio (Teleostei: Cyprinidae) in China [J]. Acta Hydrobiologica Sinica, 2002, 26(3): 281—285
杨秀平, 张敏莹, 刘焕章. 中国似逗属鱼类的形态变异及地理分化研究. 水生生物学报, 2002, 26(3): 281—285 http://www.cnki.com.cn/Article/CJFDTOTAL-SSWX200203012.htm[7] Hosoya K. Interrelationships of the Gobioninae (Cypri-nidae) [A]. In: Uyeno T, Arai T, Taniuchi T, et al. (Eds.), Indo-Pacific Fish Biology: Proceeding of the Second International Conference on Indo-Pacific Fishes [C]. Tokyo: The Ichthyological Society of Japan. 1986, 484—501
[8] 俞利荣, 乐佩琦. 似逗类鱼类的系统发育研究(鲤形目: 鲤科). 动物分类学报, 1996, 21(2): 244—255 http://www.cnki.com.cn/Article/CJFDTOTAL-DWFL602.022.htm Yu L R, Yue P Q. Studies on phylogeny of the Pseudogobiini fishes (Cypriniformes: Cyprinidae) [J]. Acta Zootaxonomica Sinica, 1996, 21(2): 244—255
俞利荣, 乐佩琦. 似逗类鱼类的系统发育研究(鲤形目: 鲤科). 动物分类学报, 1996, 21(2): 244—255 http://www.cnki.com.cn/Article/CJFDTOTAL-DWFL602.022.htm[9] Yang J Q, He S P, Freyhof J, et al. The phylogenetic relationships of the Gobioninae (Teleostei: Cyprinidae) inferred from mitochondrial cytochrome b gene sequences [J]. Hydrobiologia, 2006, 553(1): 255—266 doi: 10.1007/s10750-005-1301-3
[10] Liu H Z, Yang J Q, Tang Q Y. Estimated evolutionary tempo of East Asian gobionid fishes (Teleostei: Cyprini-dae) from mitochondrial DNA sequence data [J]. Hydrobio-logy, 2010, 55(15): 1502—1510
[11] Tang K L, Agnew M K, Chen W J, et al. Phylogeny of the gudgeons (Teleostei: Cyprinidae: Gobioninae) [J]. Molecular Phylogenetics and Evolution, 2011, 61(1): 103—124 doi: 10.1016/j.ympev.2011.05.022
[12] Jiang Z G, Zhang E. Molecular evidence for taxonomic status of the gudgeon genus Huigobio Fang, 1938 (Teleo-stei: Cypriniformes), with a description of a new species from Guangdong Province, South China [J]. Zootaxa, 2013, 3731(1): 171—182 doi: 10.11646/zootaxa.3731.1
[13] 夏曦中, 张训蒲, 刘焕章. 似逗(Pseudogobio vaillanti)的分子生物地理学分析. 自然科学进展, 2005, 15(9): 1064—1069 Xia X Z, Zhang X P, Liu H Z. Molecular biological geography analysis of Pseudogobio vaillanti [J]. Progress in Natural Science, 2005, 15(9): 1064—1069
夏曦中, 张训蒲, 刘焕章. 似逗(Pseudogobio vaillanti)的分子生物地理学分析. 自然科学进展, 2005, 15(9): 1064—1069[14] Ward R D. DNA barcode divergence among species and genera of birds and fishes [J]. Molecular Ecology Resources, 2009, 9(4): 1077—1085 doi: 10.1111/men.2009.9.issue-4
[15] 郭立, 李隽, 王忠锁, et al. 基于四个线粒体基因片段的银鱼科鱼类系统发育. 水生生物学报, 2011, 35(3): 449—459 http://ssswxb.ihb.ac.cn/CN/abstract/abstract2402.shtml Guo L, Li J, Wang Z S, et al. Phylogenetic relationships of noodle-fishes (Osmeriformes: Salangidae) based on four mitochondrial genes [J]. Acta Hydrobiologica Sinica, 2011, 35(3): 449—459
郭立, 李隽, 王忠锁, et al. 基于四个线粒体基因片段的银鱼科鱼类系统发育. 水生生物学报, 2011, 35(3): 449—459 http://ssswxb.ihb.ac.cn/CN/abstract/abstract2402.shtml[16] 陈卉, Polgar G, 傅萃长, 等. 西北太平洋海岸带大弹涂鱼复合体的隐存种与进化历史. 水生生物学报, 2014, 38(1): 75—86 http://ssswxb.ihb.ac.cn/CN/abstract/abstract40.shtml Chen H, Polgar G, Fu C Z, et al. Cryptic species and evolutionary history of Boleophthalmus pectinirostris complex along the northwestern Pacific coast [J]. Acta Hydrobiologica Sinica, 2014, 38(1): 75—86
陈卉, Polgar G, 傅萃长, 等. 西北太平洋海岸带大弹涂鱼复合体的隐存种与进化历史. 水生生物学报, 2014, 38(1): 75—86 http://ssswxb.ihb.ac.cn/CN/abstract/abstract40.shtml[17] Tominaga K, Watanabe K, Kakioka R, et al. Two highly divergent mitochondrial DNA lineages within Pseudogobio esocinus populations in central Honshu, Japan [J]. Ichthyological Research, 2009, 56(2): 195—199 doi: 10.1007/s10228-008-0071-0
[18] Tominaga K, Nakajima J, Watanabe K. Cryptic divergence and phylogeography of the pike gudgeon Pseudogobio esocinus (Teleostei: Cyprinidae): a comprehensive case of freshwater phylogeography in Japan [J]. Ichthyo-logical Research, 2016, 63(1): 79—93 doi: 10.1007/s10228-015-0478-3
[19] Hedin M. High-stakes species delimitation in eyeless cave spiders (Cicurina, Dictynidae, Araneae) from central Texas [J]. Molecular Ecology, 2015, 24(2): 346—361 doi: 10.1111/mec.13036
[20] Karanovic T, Djurakic M, Eberhard S M. Cryptic species or inadequate taxonomy? implementation of 2D geometric morphometrics based on integumental organs as landmarks for delimitation and description of copepod taxa [J]. Systematic Biology, 2016, 65(2): 304—327 doi: 10.1093/sysbio/syv088
[21] Ojanguren-Affilastro A A, Mattoni C I, Ochoa J A, et al. Phylogeny, species delimitation and convergence in the South American bothriurid scorpion genus Brachistosternus Pocock 1893: Integrating morphology, nuclear and mitochondrial DNA [J]. Molecular Phylogenetics and Evolution, 2016, 94(2016): 159—170
[22] Miller S A, Dykes D D, Polesky H F. A simple salting out procedure for extracting DNA from human nucleated cells [J]. Nucleic Acids Research, 1988, 16(3): 12—15
[23] Kim K Y, Bang I C. Molecular phylogenetic position of Abbottina springeri (Cypriniformes, Cyprindae) based on nucleotide sequences of RAG1 gene [J]. Korean Journal of Ichthyology, 2010, 22(4): 273—278
[24] Chow S, Hazama K. Universal PCR primers for S7 ribosomal protein gene introns in fish [J]. Molecular Ecology, 1998, 7(9): 1255—1256
[25] Xia X H. DAMBE5: A comprehensive software package for data analysis in molecular biology and evolution [J]. Molecular Biology and Evolution, 2013, 30(7): 1720—1728 doi: 10.1093/molbev/mst064
[26] Katoh K, Standley D M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in performance and usability [J]. Molecular Biology and Evolution, 2013, 30(4): 772—780 doi: 10.1093/molbev/mst010
[27] Tamura K, Stecher G, Peterson D, et al. MEGA6: Molecular evolutionary genetics analysis version 6.0 [J]. Molecular Biology and Evolution, 2013, 30(12): 2725—2729 doi: 10.1093/molbev/mst197
[28] Darriba D, Taboada G L, Doallo R, et al. jModelTest 2: more models, new heuristics and parallel computing [J]. Nature Methods, 2012, 9(9): 772
[29] Ronquist F, Teslenko M, Van der Mark P, et al. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space [J]. Systematic Biology, 2012, 61(3): 539—542 doi: 10.1093/sysbio/sys029
[30] Joly S, Bruneau A. Incorporating Allelic Variation for Reconstructing the Evolutionary History of Organisms from Multiple Genes: An Example from Rosa in North America [J]. Systematic Biology, 2006, 55(4): 623—636 doi: 10.1080/10635150600863109
[31] Huson D H, Bryant D. Application of phylogenetic networks in evolutionary studies [J]. Molecular Biology and Evolution, 2006, 23(2): 254—267
[32] Huelsenbeck J P, Andolfatto P. Inference of population structure under a Dirichlet process model [J]. Genetics, 2007, 175(4): 1787—1802 doi: 10.1534/genetics.106.061317
[33] Yang Z H, Rannala B. Bayesian species delimitation using multilocus sequence data [J]. Proceedings of the National Academy of Sciences, 2010, 107(20): 9264—9269 doi: 10.1073/pnas.0913022107
[34] Yang Z H, Rannala B. Unguided species delimitation using DNA sequence data from multiple loci [J]. Molecular Biology and Evolution, 2014, 31(12): 3125—3135 doi: 10.1093/molbev/msu279
[35] Bouckaert R, Heled J, Kuhnert D, et al. BEAST 2: A software platform for bayesian evolutionary analysis [J]. PLoS Computational Biology, 2014, 10(4): 1—6
[36] Rambaut A, Drummond A. Tracer v1. 5. 2007. Available at: http://tree.bio.ed.ac.uk/software/tracer.
[37] 陈宜瑜. 中国动物志硬骨鱼纲鲤形目(中卷). 北京: 科学出版社. 1998, 16—18 Chen Y Y. Fauna Sinica, Osteichthyes, Cypriniformes (II) [M]. Beijing: Science Press. 1998, 16—18
陈宜瑜. 中国动物志硬骨鱼纲鲤形目(中卷). 北京: 科学出版社. 1998, 16—18[38] Kottelat M, Freyhof J. Handbook of European freshwater fishes [M]. Delémont: Imprimerie du Démocrate SA. 2007, 15—16
[39] Hammer Ø, Harper D A T, Ryan P D. Past: Paleontological statistics software package for education and data analysis [J]. Palaeontologia Electronica, 2001, 4(1): 9
[40] Burnaby T P. Growth-invariant discriminant functions and generalized distances [J]. Biometrics, 1966, 22(1): 96—110 doi: 10.2307/2528217
[41] Rolf F J, Bookstein F L. A comment on shearing as a method for ‘size correlation’ [J]. Systematic Zoology, 1987, 36(4): 356—367 doi: 10.2307/2413400
[42] Waters J M, Rowe D L, Burridge C P. Gene trees versus species trees: reassessing life-history evolution in a freshwater fish radiation [J]. Systematic Biology, 2010, 59(5): 504—517 doi: 10.1093/sysbio/syq031
[43] Fu C Z, Guo L, Xia R. A multilocus phylogeny of Asian noodlefishes Salangidae (Teleostei: Osmeriformes) with a revised classification of the family [J]. Molecular Phylogenetics and Evolution, 2012, 62(3): 848—855 doi: 10.1016/j.ympev.2011.11.031
[44] Xia R, Durand J D, Fu C Z. Multilocus resolution of Mugilidae phylogeny (Teleostei: Mugiliformes): Implications for the family’s taxonomy [J]. Molecular Phylogene-tics and Evolution, 2016, 96: 161—177 doi: 10.1016/j.ympev.2015.12.010
[45] Sauvage H E. Rhinogobio Vaillanti, n. sp [A]. In: Bulletin de la Societe philomathique de Paris [C]. Paris: Socie-te philomathique de Paris. 1878, 87
计量
- 文章访问数: 1941
- HTML全文浏览量: 547
- PDF下载量: 35