
THE REPLACEMENT OF FISH MEAL AND SOYBEAN MEAL TO FERMENTED SOYBEAN MEAL AND ITS EFFECTS ON THE GROWTH PERFORMANCE, SERUM BIOCHEMICAL INDICES, AND IMMUNE GENE EXPRESSION IN GIANT RIVER PRAWN, MACROBRACHIUM ROSENBERGII
-
摘要: 为研究发酵豆粕在罗氏沼虾饲料中的适宜用量及替代后可能造成的影响, 以含有30%鱼粉和18%豆粕的饲料为基础饲料(T0组), 分别用2% (T2组)、5% (T5组)、8% (T8组)、15% (T15组)的发酵豆粕等蛋白替代基础饲料中的鱼粉和豆粕(2﹕1), 共配制5种等氮等能的实验饲料。选用初始均重为(0.17±0.02) g的罗氏沼虾在室内水泥池网箱中进行为期64d的养殖实验。结果显示, 发酵豆粕对鱼粉和豆粕的替代量影响罗氏沼虾的生长、血清生化及免疫基因表达。随着发酵豆粕添加量的增加, 增重率和特定生长率呈先升后降的趋势, 都以T8组最高。血清中丙二醛含量、超氧化物歧化酶活力随发酵豆粕添加量的增加均呈先升后降的趋势, 各替代组丙二醛含量均显著高于对照组(P<0.05); 血清谷丙转氨酶、谷草转氨酶活性和总蛋白含量呈先下降后上升的趋势, 且替代组不同程度低于对照组; T15组Toll受体、NF-κB、HSP70转录水平表达量显著高于其他各组(P<0.05)。以上结果表明用不同水平的发酵豆粕替代鱼粉和豆粕, 显著影响罗氏沼虾的生长性能、抗氧化能力及免疫机能; 在实验条件下, 罗氏沼虾饲料中的发酵豆粕最佳使用量为8%。Abstract: An experiment was conducted to investigate the effects of replacing fish meal (FM) and soybean meal (SBM) by fermented soybean meal (FSBM) on growth performance, serum biochemical indices, and immune genes in giant river prawn, Macrobrachium rosenbergii, and to find a suitable proportion of FSBM to replace FM in diets. Five isonitrogenous and isoenergetic diets were formulated for giant river prawn, among which a basal diet (T0) containing 30% FM and 18% SBM was treated as the control group, and the FM and SBM (2﹕1) in the basal diets replaced by 2% (T2), 5% (T5), 8% (T8) and 15% (T15) of FSBM respectively were treated as experimental diets. Giant river prawns with initial average body weight of (0.17±0.02) g were cultured for 64 days in indoor net cages at a cement tank. Results showed that with increasing FSBM supplemental level, the growth rate and specific growth rate increased firstly and then decreased, and the highest value was found in group T8. Serum superoxide dismutase activity and malondialdehyde content firstly increased and then decreased. MDA of all experimental groups was higher than that of the control group. Serum aspartate transaminase, alanine aminotransferase activity and total protein content had decreasing trend followed by an increasing trend, and the values in all experimental groups were lower than the control group. Gills Toll receptor mRNA and NF-κB mRNA expression levels and hepatopancreas heat shock protein mRNA relative expression levels in group T15 were the highest among all groups. All these results indicated that the replacement of FM and SBM with FSBM would affect growth, antioxidant ability and immunity, and the optimum amount of FSBM was 8% in the diets in this experiment.
-
鱼类早期资源指精卵结合至早期稚鱼阶段, 是鱼类个体生活史的开端[1]。通过对鱼类早期资源的研究, 可以了解鱼类的繁殖活动、产卵场位置分布和繁殖时间等早期生活史特征[1]。同时, 早期资源也是影响种群数量变化的主要原因之一[2]。然而, 捕捞、水利工程建设和江湖阻隔等人类活动造成了鱼类早期资源的急剧减少[3, 4]。例如, Sanches等[5]研究结果显示, 水利工程导致巴拉那河的鱼苗密度有显著变化。过度捕捞造成地中海沿海区域鱼类的产卵活动明显减少[6]。
长江中游是多种鱼类的产卵场和栖息地。刘明典等[7]2014—2015年在宜昌江段调查到27种鱼类卵苗。李世健等[8]在监利江段调查到51种鱼类的鱼卵和仔稚鱼。黎明政等[9]在武穴江段调查到27种鱼类的卵苗。同时, 长江中游也是重要经济鱼类四大家鱼的主要产卵场之一。20世纪60年代, 长江中游四大家鱼的产卵场有25处, 产卵量占长江总量的四分之三。其中, 宜昌江段是四大家鱼产卵规模最大的产卵场, 平均鱼卵径流量可达119.48×108粒, 占长江干流总量的9.7%[10]。20世纪80年代, 在葛洲坝修建之后, 长江中游宜昌至城陵矶江段仍有11处四大家鱼产卵场, 产卵规模为11.14×108粒, 占长江干流总规模的25.8%[11]。但是, 在三峡水库蓄水之后, 长江中游四大家鱼的繁殖活动受到了不利的影响[12]。同时, 过度捕捞, 尤其是对仔幼鱼的捕捞, 造成了鱼类资源的急剧减少[13]。此外, 三峡水库蓄水之后造成长江中游宜昌江段四大家鱼产卵时间推迟约20d, 监利江段宜昌江段四大家鱼的产卵规模减少约75%[14]。为了有效地开展长江鱼类资源的保护工作, 有必要掌握长江中游鱼类早期资源的现状及其变化规律。
本研究通过在宜都断面开展产漂流性卵鱼类早期资源调查, 了解宜昌江段产漂流性卵鱼类的繁殖情况; 结合历史资料, 分析四大家鱼及其他鱼类的繁殖规模和时间变化; 分析水文因子对产漂流性卵鱼类优势种类繁殖的影响; 为长江中游鱼类早期资源保护工作的开展提供科学依据。
1. 材料与方法
1.1 样本的采集
2017年5月12日至7月25日和2018年5月10日至7月24日, 在葛洲坝下游宜都市清江河口上约1.5—3.0 km处设置采样断面(112°26′E, 30°24′N)(图 1)。采用产漂流性卵鱼类早期资源的调查方法。采样网具包括弶网和圆锥网, 采集方法参照曹文宣等[1]和黎明政等[14]的方法。弶网用于每日采样, 记录种类和数量。圆锥网用于采集断面数据, 计算断面系数, 每年采集一次。弶网网口呈半圆形, 网口直径1 m, 网目尺寸500 μm, 网口面积0.393 m2, 网长2 m。网后部连接集苗箱, 集苗箱长40 cm, 宽和高均为30 cm, 上部敞口, 用于收集鱼卵和仔鱼。弶网采集在每日上午和下午各一次, 采集时间为7:30—9:00和16:30—18:00, 每次采集持续30—60min。同时, 测量采样点水温、透明度、流速等环境参数, 水位和流量数据来自于全国水雨情网站(http://xxfb.mwr.cn/)。圆锥网用于断面定量采样。网口呈圆形, 网口直径0.5 m, 网口面积0.196 m2, 网长2 m, 网目尺寸500 μm。圆锥网设置在断面左岸近岸点、左岸至江中心1/2处、江中心、江中心至右岸1/2处以及右岸近岸处5个采样点, 每个采样点设置表层、中层和底层3个采样水层(表层、中层和底层距水面的距离分别占水深的20%、50%和80%), 每个水层采集15min。
![]() 图 1 2017年和2018年长江中游宜都断面鱼类早期资源采样图Figure 1. Samling map of fish eggs and larvae in the Yidu section of the Yangtze River in 2017 and 2018
图 1 2017年和2018年长江中游宜都断面鱼类早期资源采样图Figure 1. Samling map of fish eggs and larvae in the Yidu section of the Yangtze River in 2017 and 20181.2 种类鉴定
鱼卵按照卵膜性质(漂流性、黏性和浮性)分类, 并依据卵径大小、胚体长等标准将大小不同的鱼卵区分培养, 培养水温维持在20—25℃, 直至孵出发育至分类特征明显的阶段, 按照形态、色素分布、肌节数目等进行种类鉴定, 具体方法参照曹文宣等[1]; 仔鱼通过显微镜观察进行种类鉴定, 之后样本用95%的乙醇保存。形态观测不能确定种类的样本用95%的乙醇固定保存在−20℃冰箱, 带回分子实验室, 通过线粒体DNA细胞色素b基因进行种类鉴定, 具体实验方法参照Xiao等[15]的方法进行。
1.3 数据处理
产卵规模计算 鱼卵和仔鱼通过断面径流量的估算方法参照易伯鲁等[10]、黎明政等[14]和母红霞等[16]的方法进行, 具体计算公式如下:
$A = M + M'\qquad\qquad\qquad\qquad\qquad\qquad$

$M = \sum\nolimits_{i = 1}^n {{T_i}} {D_i}{Q_i}C\qquad\qquad\qquad\qquad\quad$

$\begin{aligned}{M' } =& \sum\limits_{i = 1}^{n - 1} {T_{\left( {i,i + 1} \right)}'} \times C \times \left[ {\left( {{Q_i} + {Q_{i + 1}}} \right)/2} \right] \times \;\;\;\\& \left[ {\left( {{D_i} + {D_{i + 1}}} \right)/2} \right]\end{aligned}$

${D_{{i}}} = {N_i}/\left( {S\;{V_i}\;{T_i}} \right)\qquad\qquad\qquad\qquad\qquad\;\;$

$\;C = \left( {\sum\limits_{{{j}} = 1}^m {{D_j}} } \right)/\left( {m\;{D_r}} \right)\qquad\qquad\qquad\qquad$

式中, A为整个采样期内流经采集点江段面的鱼卵、仔鱼总流量; M为所有定时采集时间内鱼卵、仔鱼流量之和; M'为前后两次相邻采集之间非采集时间内计算出的鱼卵、仔鱼流量之和; i为某一特定的常规采样; i+1为第i次采样之后的一次采样; Qi为第i次采样时流经采样断面的水流量(m3/s); Di 为第i次采样时流经采样断面的仔鱼和鱼卵密度(ind./m3); C 为采样断面上的鱼卵和仔鱼的分布系数; T (i,i+1) 为第i次采样和第i+1次采样之间的时间间隔; Ni为第i次采样中采集到的鱼卵和仔鱼数量; S为网口面积(m2); Vi为第i次采样中网口的水流速(m/s); Ti为第i次采样的持续时间(s); Dj为断面采样每个采样点的鱼卵和仔鱼密度(ind./m3); j为第j个断面采样位点; m为断面采样的采样点数量; Dr为断面采样中流经靠近右岸的采样点表层的鱼卵和仔鱼密度(ind./m3)。
确定产漂流性卵鱼类优势种类 物种优势度利用Pinkas相对重要性指数(IRI, Index of relative abundance)[17]确定, 计算公式如下:
$IRI = \left( {N{\text{%}} + W{\text{%}} } \right) \times F{\text{%}} $

式中, N为某一种类鱼卵数量占采获鱼卵总数量的百分比; W为某种鱼卵生物量比例; F为某一种类出现的次数占调查总次数的百分比。由于鱼卵、鱼苗很小, 生物量难以确定, 因此只考虑数量这一因素。将IRI大于1000的种类定义为优势种。
四大家鱼产卵场位置推算 四大家鱼产卵场位置的计算公式如下:
$L = V \times T$

式中, L为四大家鱼卵、苗的漂流距离(m); V为调查江段水流速度(m/s); T为胚胎发育所需要的时间(s)。其中, 四大家鱼胚胎发育所需要的时间参照易伯鲁等[18]。
繁殖活动的水文条件分析 根据相关研究文献资料, 近些年一段断面所采集的仔鱼大多来源于三峡大坝以上江段, 鱼卵来自三峡大坝下游产卵场[14, 19]。因此,鱼类繁殖水文因子影响分析仅针对通过鱼卵鉴定的产漂流性卵鱼类的优势种类。首先, 对物种的密度及水文因子数据进行除趋势对应分析(Detrended correspondence analysis, DCA), 根据每个轴的梯度长度(Lengths of gradient, LGA)选择适合的分析方法。当LGA<3时, 采用冗余分析(Redundancy Analysis, RDA); 当LGA>4时, 采用典型对应分析(Canonical Correlation Analysis, CCA); 当4>LGA>3时, 两种分析方法皆可[20]。DCA分析结果轴梯度长度为1.8, 故本研究利用冗余分析(Redundancy Analysis,RDA)分析鱼卵密度与环境因子(水温、流量、流量日变化、透明度)之间的相关关系。
DCA和RDA分析利用Canoco 5.0[21]软件完成, 其余数据分析与作图用Excel 2010软件中完成。
2. 结果
2.1 种类组成及优势种
2017年和2018年共采集鱼卵21120粒和仔鱼2123尾。物种鉴定结果显示, 鱼卵和仔鱼共5目9科37种。其中, 鲤科(Cyprinidae) 23种, 鳅科(Cobitidae) 5种, 鲿科(Bagridae)及科(Sisoridae)各2种, 银鱼科(Salangidae)、平鳍鳅科(Homalopteridae)、鱵科(Hemiramphidae)、鮨科(Serranidae)和鰕虎鱼科(Gobiidae)各1种, 鲤科鱼类资源数量为20788粒, 占总数量的89.44%,其中鲌亚科为优势类群, 占鲤科鱼类总数量的39.45%。根据卵的性质, 漂流性卵24种, 黏性卵5种, 浮性卵1种, 沉性卵7种(表 1)。
表 1 2017年和2018年调查期间宜都断面鱼类早期资源种类组成Table 1. Species composition of eggs and larvae in the Yidu section in 2017 and 2018目Order 科Family 种Species 卵类型Type of egg 卵Eggs 苗Larvae 目Order 科Family 种Species 卵类型Type of egg 卵Eggs 苗Larvae 鲑形目Salmoniforme
银鱼科Salangidae太湖新银鱼Neosalanx taihuensis 沉性 + 圆筒吻Rhinogobio cylindricus 漂流性 + + 鲤形目Cypriniformes
鲤科Cyprinidae青鱼Mylopharyngodon piceus 漂流性 + + 蛇Saurogobio dabryi 漂流性 + + 草鱼Ctenopharyngodon idellus 漂流性 + + 光唇蛇Saurogobio gymnocheilus 漂流性 + 赤眼鳟Squaliobarbus curriculus 漂流性 + + 宜昌鳅鮀Gobiobotia filifer 漂流性 + 鳡Elopichthys bambusa 漂流性 + + 鲤Cyprinus capio 黏性 + 飘鱼Pseudolaubuca sinensis 黏性 + 鲫Carassius auratus 黏性 + 寡鳞飘鱼Pseudolaubuca engraulis 漂流性 + + 平鳍鳅科Homalopteridae 犁头鳅Lepturichthys fimbriata 漂流性 + + 贝氏䱗Hemiculter bleekeri 漂流性 + + 鳅科Cobitidae 花斑副沙鳅Parabotia fasciata 漂流性 + + 翘嘴鲌Culter alburnus 黏性 + + 武昌副沙鳅Parabotia banarescui 漂流性 + 鳊Parabramis pekinensis 漂流性 + + 紫薄鳅Leptobotia taeniops 漂流性 + 银鲴Xenocypris argentea 漂流性 + + 红唇薄鳅Leptobotia rubrilabris 漂流性 + 黄尾鲴Xenocypris davidi 黏性 + 鲇形目Siluriformes
鲿科Bagridae瓦氏黄颡鱼Pelteobagrus vachelli 沉性 + 似鳊Pseudobrama simoni 漂流性 + 光泽黄颡鱼Pelteobaggrus nitidus 沉性 + 鳙Hypophthalmichthys nobilis 漂流性 + + 科Sisoridae 中华纹胸Glyptothorax sinensis 沉性 + 鲢Hypophthalmichthys molitrix 漂流性 + + 福建纹胸Glyptothorax fukiensis 沉性 + 银Squalidus argentatus 漂流性 + + 颌针鱼目Beloniformes
鱵科Hemiramphidae间下鱵Hyporhamphus intermedius 沉性 + 铜鱼Coreius heterodon 漂流性 + + 鲈形目Perciformes
鮨科Serranidae鳜Siniperca chuatsi 浮性 + + 吻Rhinogobio typus 漂流性 + 鰕虎鱼科Gobiidae 子陵吻鰕虎鱼Rhinogobius giurinus 沉性 + 注: “+”代表可采集到Note: “+” means can be collected 2017年和2018年采集到鱼卵28种, 有青鱼(Mylopharyngodon piceus)、草鱼(Ctenopharyngodon idellus)、赤眼鳟(Squaliobarbus curriculus)、贝氏䱗(Hemiculter bleekeri)、鳙(Hypophthalmichthys nobilis)、鲢(Hypophthalmichthys molitrix)和银(Squalidus argentatus)等种类。采集到仔鱼27种, 有太湖新银鱼(Neosalanx taihuensis)、寡鳞飘鱼(Pseudolanbuca engraulis)、贝氏䱗、间下鱵(Hyporhamphus intermedius)和子陵吻鰕虎鱼(Rhinogobius giurinus)等种类。
2017年, 采集鱼卵10572粒, 仔鱼1221尾, 其中四大家鱼卵3172粒, 贝氏䱗卵2725粒, 分别占卵总数量的30.0%、25.8%; 仔鱼以贝氏䱗和寡鳞飘鱼为主, 分别占比54.3%、24.4%。2018年共采集鱼卵10548粒, 仔鱼902尾, 其中四大家鱼卵4249粒, 贝氏䱗卵2448粒, 分别占卵总数量的40.3%和23.2%; 仔鱼以子陵吻鰕虎鱼、寡鳞飘鱼和贝氏䱗为主, 分别占比22.7%、19.3%和17.7%。根据2017年和2018年采集卵和仔鱼数量分析, 宜都断面鱼卵优势种为四大家鱼(IRI=1433)和贝氏䱗(IRI=1319); 仔鱼优势种为贝氏䱗(IRI=1259)和寡鳞飘鱼(IRI=1169)。
2.2 产卵时间
2017年5月采集鱼卵以银和飘鱼为主, 占当月总卵数的27.13%和19.38%, 6月四大家鱼和贝氏䱗成为主要种类, 数量占比分别为38.89%和26.12%。在7月四大家鱼数量占比下降至18.50%, 而贝氏䱗上升至43.24%。2018年稍有不同, 四大家鱼数量百分比自5月的55.38%逐渐下降至7月的15.04%, 贝氏䱗相对比例依旧在6月达到最高, 占当月数量的42.77%, 7月保持稳定, 占当月数量的41.00%。2017年和2018年调查期间, 飘鱼和银所占数量比自5月至7月逐渐减少, 而花斑副沙鳅和鳊则相反, 自5月比例逐渐上升。2017年四大家鱼的产卵时间是从5月14日到7月2日。2018年则是从5月10日至7月5日。
仔鱼5月采集较少, 2017年5月仅采集仔鱼5尾, 分别为间下鱵、光泽黄颡鱼和光唇蛇, 2018年5月采集仔鱼16尾, 分别为太湖新银鱼、寡鳞飘鱼和贝氏䱗。仔鱼采集数量主要在6月和7月。2017年和2018年采集结果显示, 仔鱼中贝氏䱗和寡鳞飘鱼为优势种类(图 2)。
![]() 图 2 2017年和2018年宜都断面主要类群鱼卵(a)、仔鱼(b)的相对丰度Figure 2. Abundance percentage of eggs (a) and larvae (b) of dominant species in the Yidu section in 2017 and 2018
图 2 2017年和2018年宜都断面主要类群鱼卵(a)、仔鱼(b)的相对丰度Figure 2. Abundance percentage of eggs (a) and larvae (b) of dominant species in the Yidu section in 2017 and 20182.3 产卵规模
2017年, 通过宜都断面的鱼卵径流量为124.45×108粒, 鱼卵中四大家鱼径流量为36.13×108粒, 占总径流量的29.03%; 贝氏䱗径流量为19.96×108粒, 占总径流量的16.04%。2018年通过宜都断面的鱼卵径流量为101.07×108粒, 鱼卵中四大家鱼径流量为35.20×108粒, 占总径流量的34.83%; 贝氏䱗的鱼卵径流量为19.17×108粒, 占总径流量的18.97%。
2017年通过宜都断面的仔鱼径流量为16.43×108尾, 寡鳞飘鱼和贝氏䱗径流量分别为5.89×108尾和2.91×108尾, 占仔鱼总径流量的35.87%和17.72%。四大家鱼的总径流量为4.12×108尾, 占仔鱼总径流量的25.07%。2018年通过宜都断面的仔鱼径流量为8.29×108尾, 寡鳞飘鱼和贝氏䱗分别为0.97×108尾和1.22×108尾, 占仔鱼总径流量的11.72%和14.70%。四大家鱼的总径流量为0.11×108尾, 占仔鱼总径流量的1.32%。
2.4 四大家鱼的产卵场位置
通过对采集到的四大家鱼卵发育期的观测, 0.14%处于128细胞期前, 95.31%处于桑椹期至眼囊期, 4.55%处于尾芽期至心脏搏动期, 距受精1—33h。其中, 65.76%的四大家鱼卵处于原肠早期至原肠晚期, 距受精7—13h。四大家鱼仔鱼均处于鳔雏形期和鳔一室期, 距受精98—146h。根据鱼卵和仔鱼的发育期以及流速分析, 四大家鱼鱼卵来源于宜都断面2.4 km以上的江段。产卵场位置主要分布在葛洲坝下至庙咀、胭脂坝至云池江段。四大家鱼仔鱼应来源于三峡大坝以上江段(图 3)。
![]() 图 3 长江中游宜昌江段四大家鱼产卵场位置Figure 3. Spawning grounds of the four major Chinese carps in the Yichang reach in the middle Yangtze River
图 3 长江中游宜昌江段四大家鱼产卵场位置Figure 3. Spawning grounds of the four major Chinese carps in the Yichang reach in the middle Yangtze River2.5 产漂流性卵鱼类及关键环境因子
2017年最小流量在7月4日, 为6920 m3/s, 最大流量在7月11日, 为30200 m3/s。整个调查期间, 水温介于18.5—26.0℃, 5月12日达到18℃以上, 7月20日达到24℃以上。2017年出现4次产卵高峰, 分别在6月10日、6月11日、6月18日和7月10日。高峰期产卵规模总计为60.7×108粒, 占2017年总产卵规模的48.7%。
2018年6月12日流量最小, 为10200 m3/s, 最大流量在7月14日, 为43800 m3/s。整个调查期间, 水温为18.5—25.2℃, 5月10日达到18℃以上, 7月2日达到24℃以上。2018年出现2次显著的产卵高峰, 分别为5月19日和6月24日, 产卵规模共为35.9×108粒, 占2018年采集鱼卵总规模的35.5%(图 4)。
![]() 图 4 2017年(a)和2018年(b)调查期间宜都断面鱼卵日密度变化图Figure 4. Discharge and eggs density drifting through the Yidu section in 2017 (a) and 2018 (b)
图 4 2017年(a)和2018年(b)调查期间宜都断面鱼卵日密度变化图Figure 4. Discharge and eggs density drifting through the Yidu section in 2017 (a) and 2018 (b)应用RDA分析方法, 对鱼卵漂流密度与水文因子的关系进行分析。在前2个排序轴上, 物种与水文因子的相关性为0.426和0.345, 第一排序轴和第二排序轴的累积贡献率为92.83%。RDA分析结果显示, 四大家鱼卵总密度和花斑副沙鳅卵密度受流量日变化和流量影响较大; 贝氏䱗鱼卵密度受流量影响较大; 鳊鱼卵密度受水温影响较大; 银鱼卵密度主要受透明度影响(图 5)。
![]() 图 5 2017年和2018年宜都断面水文因子对漂流性卵密度的影响Figure 5. Effects of hydrological factors on the density of the drifting eggs in the Yidu section in 2017 and 2018
图 5 2017年和2018年宜都断面水文因子对漂流性卵密度的影响Figure 5. Effects of hydrological factors on the density of the drifting eggs in the Yidu section in 2017 and 20183. 讨论
3.1 长江中游鱼类繁殖现状及变化
本研究结果显示, 2017年和2018年5—7月通过长江中游宜都断面的鱼类早期资源较丰富。已有的研究结果显示, 母红霞[22]发现2009年和2010年5—7月通过宜都断面鱼卵径流量分别为57.26×108粒和64.45×108粒, 年均60.86×108粒。与2009年和2010年同比, 2017年和2018年通过宜都断面的鱼卵径流量明显增加, 年均径流量增加了85.3%。其中, 宜昌江段四大家鱼的产卵规模明显增大。2005—2012年5—7月通过宜都断面四大家鱼鱼卵径流量为(0.03—5.85)×108粒, 年均2.49×108粒[14], 而2017年和2018年四大家鱼卵年均35.67×108粒, 同比增加了13倍, 达到了20世纪80年代的水平[10, 11]。鱼卵径流量增加的主要原因可能有2个。第一, 2009年起, 原农业部在长江中游开展较大规模的四大家鱼亲鱼放流, 增加了繁殖群体数量[7, 23]。第二, 2011年三峡工程开始生态调度试验, 在一定程度上促进了鱼类的繁殖活动。徐薇等[24]的研究结果显示, 2012年三峡水库试验性生态调度期间长江中游沙市江段鱼卵总径流量和家鱼卵径流量均出现高峰, 繁殖种类明显增加。周雪等[25]的研究结果显示, 2013—2017年三峡水库生态调度造成长江中游监利江段涨水持续时间达到4d及以上, 期间四大家鱼的卵苗量较大, 占年总量的45%以上。
在三峡水库蓄水运行之后, 长江中游四大家鱼的产卵时间推迟。在三峡水库蓄水之前, 长江中游宜昌江段四大家鱼产卵时间从4月中下旬开始[10]。在蓄水后, 2005—2012年宜昌江段四大家鱼的产卵时间推迟到5月8日至5月28日, 平均推迟了21.8d, 导致产卵时间推迟的主要原因是下泄水温滞后, 到达18℃的时间推迟[14]。与2005—2012年相比, 2017年和2018年宜昌江段四大家鱼产卵时间没有明显变化, 产卵场位置也没有明显变化。因此, 本研究的结果表明, 在三峡水库蓄水运行之后, 长江中游四大家鱼产卵场的环境条件可能已经趋于稳定。如果亲本增加, 宜昌江段产卵场依然可以满足其完成繁殖活动, 产生较大规模的鱼卵和仔鱼。
本研究结果还显示, 2017年和2018年5—7月有一定规模的仔鱼通过长江中游宜都断面, 其中四大家鱼的仔鱼均来源于三峡大坝以上江段。《长江三峡水利枢纽环境影响报告书》认为, 在三峡水库蓄水之后, 长江上游涪陵以下的8个四大家鱼产卵场将消失, 产卵场将上移。根据母红霞[22]、王红丽等[26]、Li等[14]和马琴[27]在三峡水库库区的鱼类早期资源调查结果显示, 涪陵以下江段仍有产卵规模较小的四大家鱼的产卵场。2005—2012年, 通过宜都断面四大家鱼仔鱼径流量年均值为4.65×108尾[14]。2013年和2014年分别为1.52×108尾和1.27×108尾[27]。与2005—2014年相比, 2017年和2018年没有明显的变化。研究结果表明, 三峡水库库区内在一定水文条件下仍具备四大家鱼繁殖的条件, 且可以通过三峡大坝进入长江中游, 可以为长江中游鱼类资源提供一定的补充。
3.2 鱼类早期资源与环境因子的关系
本研究结果显示, 四大家鱼、贝氏䱗和花斑副沙鳅等种类的产卵规模与流量和流量日变化有着密切的关系。鱼类的繁殖活动受外界环境因子的影响[28]。已有的研究提出“最佳环境窗口”假说, 认为每个物种都有特定的环境条件需求, 以满足繁殖活动和保证产生最多的后代[29—31]。Li等[32]的研究结果显示, 四大家鱼的产卵活动需要18—24℃的水温和0.55 m/d的水位涨幅。Wang等[12]也发现, 四大家鱼的仔鱼丰度与流量增加持续时间显著相关。因此, 水温应该是四大家鱼等产漂流性卵鱼类的繁殖限制条件, 达到了一定水温鱼类才能开始繁殖活动[10, 33]。流量等则是刺激鱼类繁殖和促进繁殖活动的重要环境条件[10, 34]。
3.3 长江中游鱼类早期资源保护措施
鱼类早期资源减少导致种群补充量减少[35, 36], 是造成长江鱼类资源急剧减少的主要原因之一[1, 37]。Ma等[38]的研究结果显示, 长江中游对于通江湖泊中鱼类早期资源的补充, 维持鱼类多样性有重要意义。为了长江鱼类资源保护, 2019年农业农村部、财政部和人力资源社会保障部联合印发《长江流域重点水域禁捕和建立补偿制度实施方案》, 长江禁渔已经逐步实施, 2021年长江将全面禁渔。这一措施将消除捕捞对鱼类资源的影响, 尤其是对湖区等仔幼鱼资源的影响, 能够显著提高鱼类早期生活史阶段的存活率, 增加种群的补充量。长江鱼类的早期生活史阶段还面临着栖息地减少的问题。长江中下游的湖泊和河漫滩原本是仔幼鱼的主要栖息地。但是, 长江中下游的通江湖泊仅剩石臼湖、洞庭湖和鄱阳湖, 其余湖泊都已经被阻隔, 仔幼鱼无法进入湖泊生长, 其栖息地功能已经完全丧失。Liu等[39]的研究结果显示, 长江中下游江湖连通性减少已经严重影响了鱼类多样性。同时, 港口建设、防洪护坡和城市景观等建设破坏了河漫滩的自然环境条件, 减少了仔幼鱼的栖息地[40, 41]。因此, 为了更加有效地开展鱼类资源保护, 结合本研究结果, 还建议通过恢复江湖连通、灌江纳苗, 以及维持河漫滩自然环境条件, 增加仔幼鱼的栖息地面积; 开展重要经济鱼类亲本放流, 增加繁殖群体规模; 开展生态调度, 促进繁殖活动。
致谢:
感谢中国科学院水生生物研究所胡华明、李文静、杨萍、张文武和张晨在样本采集、数据处理上给予的帮助!
-
![]()
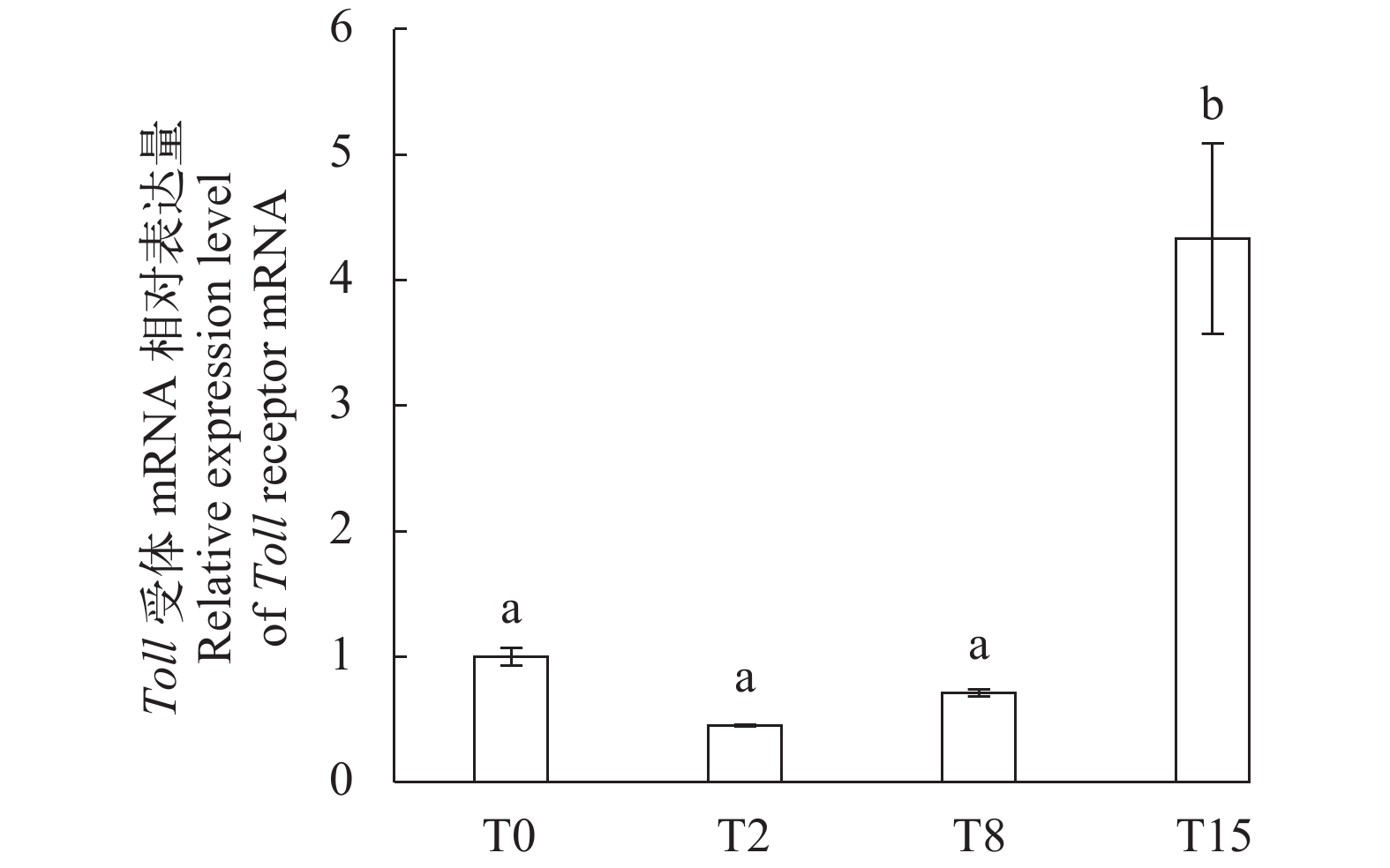
图 1 不同饲料组罗氏沼虾Toll受体mRNA在鳃中的相对表达量
图中相同或无小写字母表示差异不显著(P>0.05), 不同小写字母表示差异显著(P<0.05); 下图同
Figure 1. Toll receptor mRNA expression levels in gills of shrimp fed with different diets
Values with the same small letter superscripts or none superscripts mean no significant difference (P>0.05), while different small letter superscripts mean significant difference (P<0.05). The same applies below
![]()
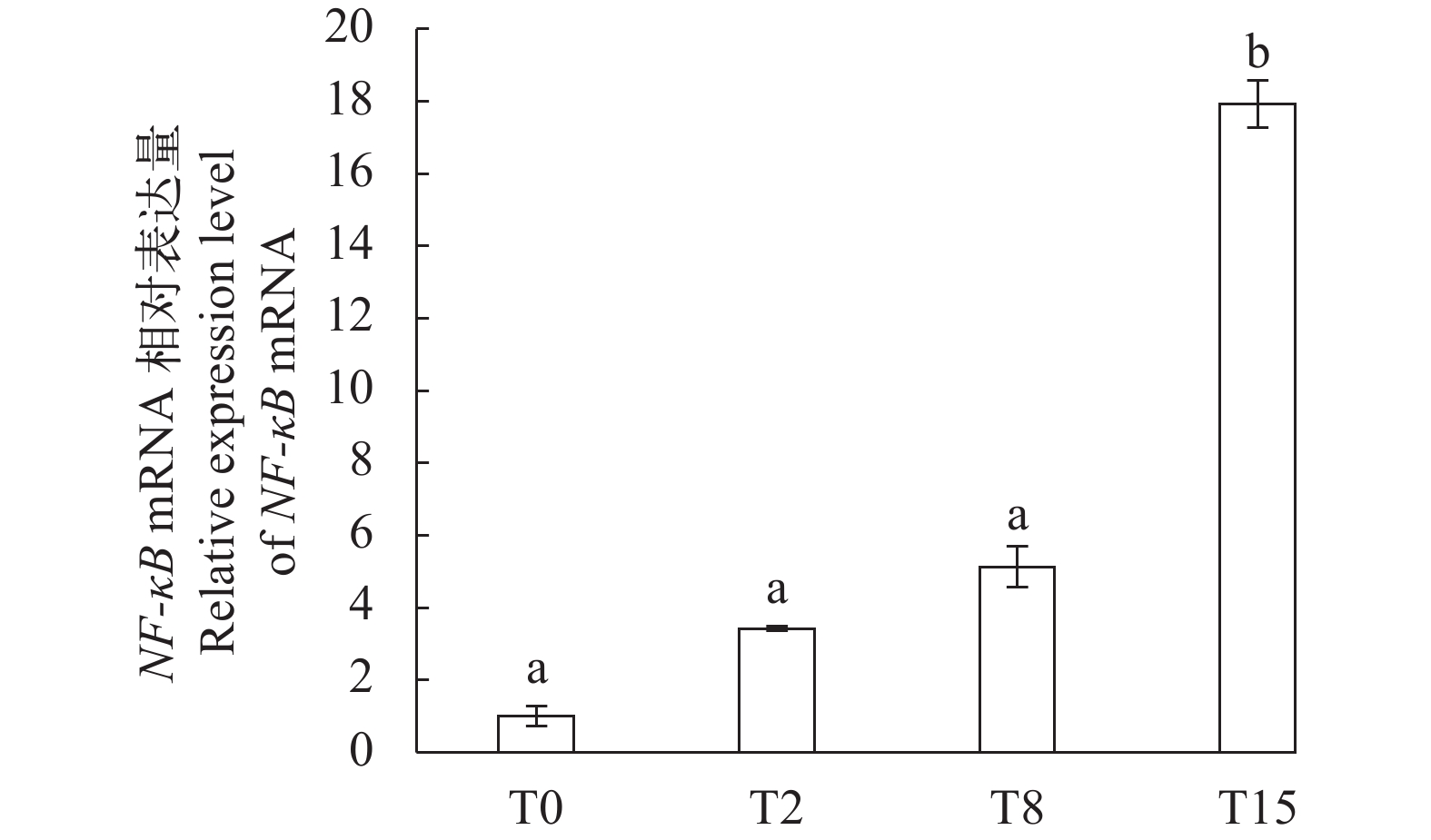
图 2 不同饲料组罗氏沼虾NF-κB mRNA在鳃中的相对表达量
Figure 2. NF-κB mRNA expression levels in gills of shrimp fed with different diets
![]()
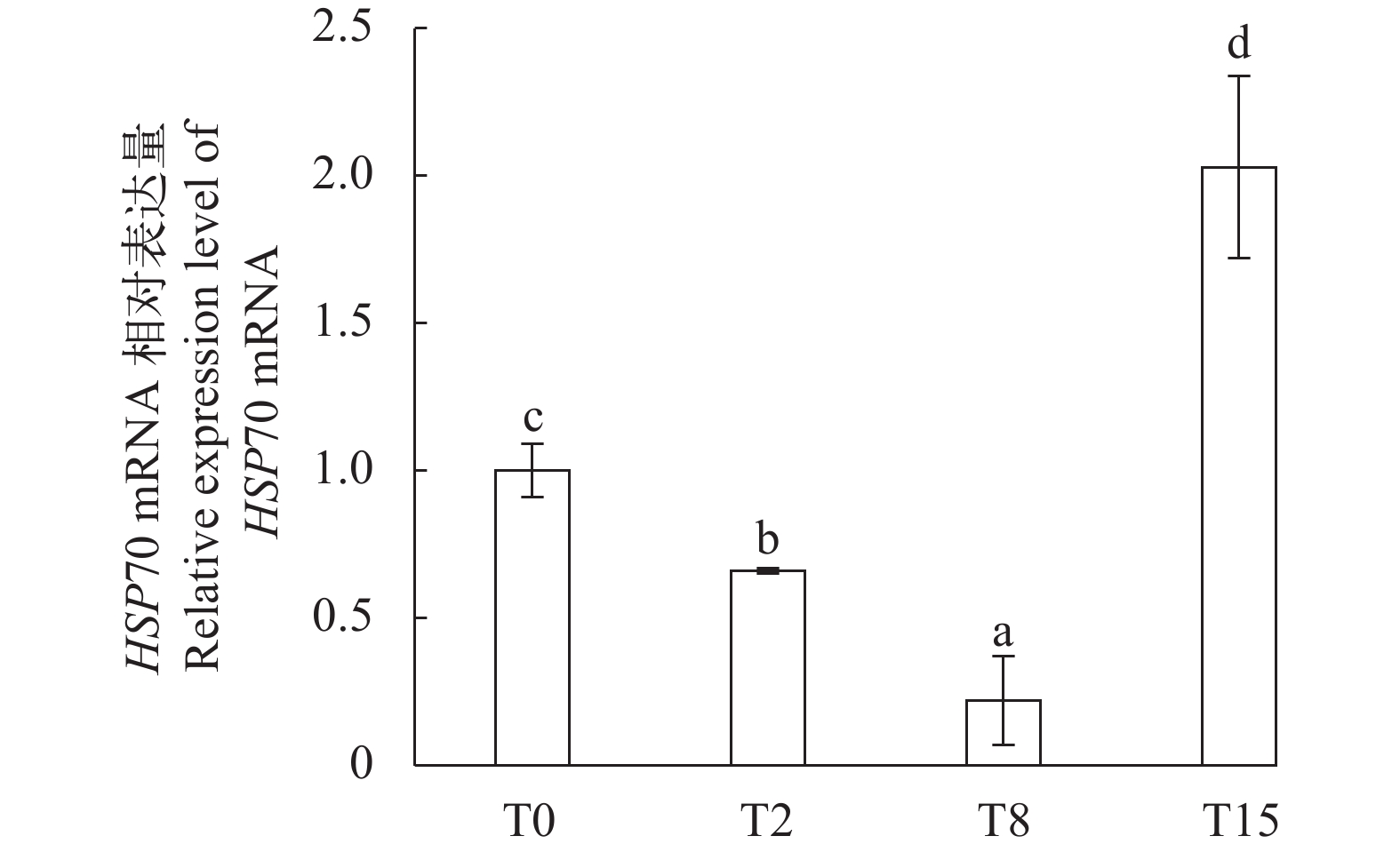
图 3 不同饲料组罗氏沼虾HSP70 mRNA在肝胰腺中的相对表达量
Figure 3. HSP70 mRNA expression levels in hepatopancreas of shrimp fed with different diets
表 1 饲料组成和营养水平(%, 风干样品)
Table 1 Composition and nutrient levels of diets (%, air dry basis)
项目Item 饲料Diet T0 T2 T5 T8 T15 秘鲁鱼粉Peru fish meal 30.00 28.80 27.00 25.50 21.50 玉米蛋白粉Corn gluten meal 5.00 5.00 5.00 5.00 5.00 豆粕Soybean meal 18.00 17.64 17.10 16.65 15.45 发酵豆粕Fermented soybean meal 0.00 2.00 5.00 8.00 15.00 花生粕Peanut meal 7.00 7.00 7.00 7.00 7.00 肉粉Meat meal 7.00 7.00 7.00 7.00 7.00 面粉Wheat flour 25.10 24.66 24.00 22.95 20.80 复合多矿1Mineral premix 0.70 0.70 0.70 0.70 0.70 复合多维2Vitamin premix 0.80 0.80 0.80 0.80 0.80 乌贼膏Squid meal 2.00 2.00 2.00 2.00 2.00 鱼油Fish oil 1.20 1.20 1.20 1.20 1.40 磷脂粉Phospholipid meal 2.00 2.00 2.00 2.00 2.00 蛋氨酸Met 0.30 0.30 0.30 0.30 0.35 赖氨酸Lys 0.70 0.70 0.70 0.70 0.80 苏氨酸Thr 0.20 0.20 0.20 0.20 0.20 合计Total 100.00 100.00 100.00 100.00 100.00 营养水平Nutrient levels 粗蛋白Crude protein 43.29 43.26 43.23 43.38 43.40 粗脂肪Crude fat 7.07 7.02 6.94 6.88 6.89 粗灰分Ash 9.12 9.03 9.04 8.97 8.90 水分Moisture 10.73 11.31 10.85 10.37 11.40 蛋氨酸Met 1.11 1.10 1.08 1.07 1.09 赖氨酸Lys 3.20 3.19 3.16 3.15 3.21 苏氨酸Thr 1.77 1.77 1.77 1.78 1.77 注: 1. 每千克矿物质预混料含有mineral premix content per kg: Ca 10.5 g, K 90 g, Mg 12 g, Fe 1.0 g, Cu 3.0 g, Zn 10 g, M n 3.8 g, Co 0.8 g, Se 20 mg; 2. 每千克维生素预混料含有vitamin premix content per kg: VA 8000000 IU, VD 2000000 IU, VE 50 g, VK 10 g, VB1 5 g, VB2 15 g, VB6 8 g, VB12 0.02 g, 烟酰胺nicotinamide 40 g, D-泛酸钙calcium D-pantothenate 25 g, 叶酸folic acid 2.5 g, 生物素biotin 0.08 g, 肌醇inositol 100 g  下载: 导出CSV
下载: 导出CSV
表 2 qRT-PCR引物序列
Table 2 Primers sequence for qRT-PCR
引物名称
Primer nameGenBank登录号
GenBank accession No.引物序列
Primer sequence (5′—3′)引物类型
Primer typeToll receptor JF895474.1 TCTACGACCGCAACGAGC Forward CGGAGTGGGAGTGAACAG Reverse HSP70 HG001455.1 CTCTGCCCAAGCAAGTAT Forward GAATCTGTGCCTTATCCA Reverse NF-κB KR827675.1 GTGGCTCACTTACGACTC Forward AAGGTCCATACTCTTTGC Reverse β-actin AF221096.1 GTGCGTGACATCAAGGAA Forward TTGTAGGTGGTCTCGTGAAT Reverse
下载: 导出CSV
表 3 发酵豆粕替代鱼粉和豆粕对罗氏沼虾生长性能的影响
Table 3 Effects of replacement of FM and SBM by FSBM on growth indices of Macrobrachium rosenbergii
组别Group 成活率SR (%) 增重率WGR (%) 特定生长率SGR (%/d) 饲料系数FCR T0 90.00±3.33 2290.58±60.58c 4.90±0.03b 1.12±0.13 T2 93.34±6.67 2050.53±45.27ab 4.78±0.02a 1.18±0.05 T5 95.83±6.51 2195.30±169.19bc 4.89±0.11b 1.21±0.21 T8 87.50±0.83 2299.44±38.34c 4.96±0.02b 1.17±0.04 T15 89.17±1.67 2024.71±4.71a 4.78±0.01a 1.33±0.09 注: 同列数据肩标相同或无小写字母表示差异不显著(P>0.05), 不同小写字母表示差异显著(P<0.05); 下表同 Note: Values with the same small letter superscripts or none letter superscripts in the same row mean no significant difference (P>0.05), while different small letter superscripts mean significant difference (P<0.05). The same applies below
下载: 导出CSV
表 4 发酵豆粕替代鱼粉和豆粕对罗氏沼虾血清抗氧化指标的影响
Table 4 Effects of replacement of FM and SBM by FSBM on antioxidant indices in serum of Macrobrachium rosenbergii
组别Group 超氧化物歧化酶SOD (U/mL) 丙二醛MDA (nmol/mL) T0 343.45±7.09b 40.63±0.31a T2 357.62±12.70b 90.61±0.86c T5 399.58±13.28c 98.21±2.34d T8 399.91±9.08c 94.05±1.06c T15 312.24±10.36a 78.09±1.85b
下载: 导出CSV
表 5 发酵豆粕替代鱼粉和豆粕对罗氏沼虾血清生化指标的影响
Table 5 Effects of replacement of FM and SBM by FSBM on biochemical indices in serum of Macrobrachium rosenbergii
组别Group 谷草转氨酶AST (U/L) 谷丙转氨酶ALT (U/L) 白蛋白ALB (g/L) 总蛋白TP (g/L) T0 74.95±3.11c 68.55±7.01c 11.75±0.21 100.05±1.48b T2 67.60±1.20c 42.23±1.10b 12.13±1.31 97.55±6.01b T5 54.33±1.59b 37.58±2.15b 11.93±0.57 88.40±2.34a T8 12.10±0.99a 29.30±3.84a 13.00±1.56 88.78±1.78a T15 49.42±6.78b 37.53±4.48b 12.63±2.55 94.98±3.06ab
下载: 导出CSV
-
[1] 杨国梁, 陈雪峰, 王军毅, 等. 罗氏沼虾产业在中国持续增长的经济与社会原因分析. 浙江海洋学院学报(自然科学版), 2011, 30(5): 450—457 Yang G L, Chen X F, Wang J Y, et al. Social and economical factors of sustained growth of Macrobrachium rosenbergii industry in mainland China [J]. Journal of Zhejiang Ocean University, 2011, 30(5): 450—457
杨国梁, 陈雪峰, 王军毅, 等. 罗氏沼虾产业在中国持续增长的经济与社会原因分析. 浙江海洋学院学报(自然科学版), 2011, 30(5): 450—457[2] 王秋举, 张曦, 邓君明, 等. 豆粕替代鱼粉在鱼类饲料中的研究进展. 农业与技术, 2012, 32(1): 33—34 Wang Q J, Zhang X, Deng J M, et al. Research progress on soybean meal substitute for fish meal in fish feed [J]. Agriculture and Technology, 2012, 32(1): 33—34
王秋举, 张曦, 邓君明, 等. 豆粕替代鱼粉在鱼类饲料中的研究进展. 农业与技术, 2012, 32(1): 33—34[3] Friedman M. Nutritional value of proteins from different food sources. A review [J]. Journal of Agricultural and Food Chemistry, 1996, 44(1): 6—29
[4] Hasan M R, Macintosh D J, Jauncey K. Evaluation of some plant ingredients as dietary protein sources for common carp (Cyprinus carpio L) fry [J]. Aquaculture, 1997, 151(1–4): 55—70
[5] Bureau D P, Harris A M, Cho C Y. The effects of purified alcohol extracts from soy products on feed intake and growth of chinook salmon (Oncorhynchus tshawytscha) and rainbow trout (Oncorhynchus mykiss) [J]. Aquaculture, 1998, 161(1–4): 27—43
[6] Gomes E F, Rema P, Kaushik S J. Replacement of fish meal by plant proteins in the diet of rainbow trout (Oncorhynchus mykiss): digestibility and growth performance [J]. Aquaculture, 1995, 130(2–3): 177—186
[7] Krogdahl A, Bakkemckellep A M, Roed K H, et al. Feeding Atlantic salmon Salmo salar L. soybean products: effects on disease resistance (furunculosis), and lysozyme and IgM levels in the intestinal mucosa [J]. Aquaculture Nutrition, 2015, 6(2): 77—84
[8] Ringø E, Sperstad S, Myklebust R, et al. Characterisation of the microbiota associated with intestine of Atlantic cod (Gadus morhua L): The effect of fish meal, standard soybean meal and a bioprocessed soybean meal [J]. Aquaculture, 2006, 261(3): 829—841
[9] Francis G, Makkar H P S, Becker K. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish [J]. Aquaculture, 2001, 199(3): 197—227
[10] Heikkinen J, Vielma J, Kemilainen O, et al. Effects of soybean meal based diet on growth performance, gut histopathology and intestinal microbiota of juvenile rainbow trout (Oncorhynchus mykiss) [J]. Aquaculture, 2006, 261(1): 259—268
[11] Yue H, Zhan Z F, Guo P F, et al. Soybean beta -conglycinin-induced gut hypersensitivity reaction in a piglet model [J]. Archives of Animal Nutrition, 2009, 63(3): 188—202
[12] 张锦秀. 大豆蛋白源对幼建鲤肠道免疫功能的影响. 四川农业大学. 2003 Zhang J X. The effects of soybean protein source on growth performance and intestinal immune of juvenile Jian carp (Cyprinus carpiovar Jian) [D]. Sichuan Agricultural University. 2003
张锦秀. 大豆蛋白源对幼建鲤肠道免疫功能的影响. 四川农业大学. 2003[13] Baeverfjord G, Krogdahl A. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L. distal intestine: a comparison with the intestines of fasted fish [J]. Journal of Fish Diseases, 2010, 19(5): 375—387
[14] 陈晓明, 华雪铭, 朱伟星, 等. 大豆抗原蛋白对南美白对虾生长、消化及非特异性免疫的影响. 动物营养学报, 2015, 27(7): 2115—2127 Chen X M, Hua X M, Zhu W X, et al. Effects of soybean allergic proteins on growth, digestion and non-specific immune of Litopenaeus vannamei [J]. Acta Zoonutrimenta Sinica, 2015, 27(7): 2115—2127
陈晓明, 华雪铭, 朱伟星, 等. 大豆抗原蛋白对南美白对虾生长、消化及非特异性免疫的影响. 动物营养学报, 2015, 27(7): 2115—2127[15] Ouoba L I I, Cantor M D, Diawara B, et al. Degradation of African locust bean oil by Bacillus subtilis and Bacillus pumilus isolated from soumbala, a fermented African locust bean condiment [J]. Journal of Applied Microbiology, 2003, 167: 273—281
[16] Ilham I, Fotedar R. Growth, enzymatic glutathione peroxidase activity and biochemical status of juvenile barramundi (Lates calcarifer) fed dietary fermented soybean meal and organic selenium [J]. Fish Physiology and Biochemistry, 2017, 43(3): 775—790
[17] 吕云云, 常青, 陈四清, 等. 发酵豆粕对圆斑星鲽生长及消化能力的影响. 水生生物学报, 2016, 40(1): 10—18 Lv Y Y, Chang Q, Chen S Q, et al. The effects of dietary fermented soybean meal on the growth and digestive ability of spotted halibut, Verasper variegatus [J]. Acta Hydrobiologica Sinica, 2016, 40(1): 10—18
吕云云, 常青, 陈四清, 等. 发酵豆粕对圆斑星鲽生长及消化能力的影响. 水生生物学报, 2016, 40(1): 10—18[18] Liang X F, Hu L, Dong Y C, et al. Substitution of fish meal by fermented soybean meal affects the growth performance and flesh quality of Japanese seabass (Lateolabrax japonicus) [J]. Animal Feed Science & Technology, 2017, 229: 1—12
[19] 冷向军, 王文龙, 李小勤. 发酵豆粕部分替代鱼粉对凡纳滨对虾的影响. 粮食与饲料工业, 2007, 2007(3): 40—41 Leng X J, Wang W L, Li X Q. Experiment on Feeding Penaeus Vannamei boones with fermented soybean meal as partial substitute for fish meal [J]. Cereal & Feed Industry, 2007, 2007(3): 40—41
冷向军, 王文龙, 李小勤. 发酵豆粕部分替代鱼粉对凡纳滨对虾的影响. 粮食与饲料工业, 2007, 2007(3): 40—41[20] 董云伟, 牛翠娟. 豆粕替代鱼粉对罗氏沼虾生长和消化酶活性的影响. 北京师范大学学报(自然科学版), 2000, 36(2): 260—263 Dong Y W, Niu C J. Effect of dietary protein sources on growth and activities of digestive enzymes of giant freshwater prawn (Macrobrachium rosenbergii) [J]. Journal Of Beijing Normal University Natural Sciences, 2000, 36(2): 260—263
董云伟, 牛翠娟. 豆粕替代鱼粉对罗氏沼虾生长和消化酶活性的影响. 北京师范大学学报(自然科学版), 2000, 36(2): 260—263[21] Kiers J L, Meijer J C, Nout M J R, et al. Effect of fermented soya beans on diarrhoea and feed efficiency in weaned piglets [J]. Journal of Applied Microbiology, 2003, 95(3): 545—52
[22] Liu X, Feng J, Xu Z, et al. The effects of fermented soybean meal on growth performance and immune characteristics in weaned piglets [J]. Turkish Journal of Veterinary & Animal Sciences, 2007, 31(5): 341—345
[23] Hardy R W, Gatlin D M, Bureau D P, et al. Nutrient Requirements of Fish and Shrimp [M]. National Research Council (NRC). 2011, 242
[24] Fish B C, Thompson L U. Lectin-tannin interactions and their influence on pancreatic amylase activity and starch digestibility [J]. Journal of Agricultural & Food Chemistry, 1991, 39(4): S91—S92
[25] 宋文新. 黑鲷幼鱼饲料中发酵豆粕部分替代鱼粉的研究. 浙江大学. 2009 Song W X. Partial replacement of fish meal by fermented soybean meal in diets for juvenile black sea bream, Sparus macrocepalus [D]. Zhejiang University. 2009
宋文新. 黑鲷幼鱼饲料中发酵豆粕部分替代鱼粉的研究. 浙江大学. 2009[26] 孔纯. 暗纹东方鲀对大豆抗原蛋白的摄食调节与肠道健康响应研究. 上海海洋大学. 2017 Kong C. Study on feeding regulation and intestinal health response of obscure puffer (Takifugu fasciatus) to soybean antigen protein [D]. Shanghai Ocean University. 2017
孔纯. 暗纹东方鲀对大豆抗原蛋白的摄食调节与肠道健康响应研究. 上海海洋大学. 2017[27] 孟兆娜, 陈玉春, 管雪婷, 等. 复方中草药对镜鲤(Cyprinus carpio L)血清转氨酶及红细胞抗氧化酶活性的影响. 东北农业大学学报, 2010, 41(8): 75—80 Yao Z N, Chen Y C, Guan X T. et al. Effect of Chinese herb compounds on activities of transaminase in serum and antioxidase in erythrocyte of Cyprinus carpio L [J]. Journal of Northeast Agricultural University, 2010, 41(8): 75—80
孟兆娜, 陈玉春, 管雪婷, 等. 复方中草药对镜鲤(Cyprinus carpio L)血清转氨酶及红细胞抗氧化酶活性的影响. 东北农业大学学报, 2010, 41(8): 75—80[28] Richard J S, Carl B S, James T G. Corticoid stress responses to handling and temperature in Salmonids [J]. Transactions of the American Fisheries Society, 1977, 106(3): 213—218
[29] 陈晓明. 发酵豆粕替代鱼粉对凡纳对虾摄食生理和抗氨氮应激的影响. 上海海洋大学. 2015 Cheng X M. Effect of fish meal replacement by fermented soybean meal on physiology of feeding and the resistance to ammonia stress of Litopenaeus vannamei [D]. Shanghai Ocean University. 2015
陈晓明. 发酵豆粕替代鱼粉对凡纳对虾摄食生理和抗氨氮应激的影响. 上海海洋大学. 2015[30] Djangmah J S. The effects of feeding and starvation on copper in the blood and hepatopancreas, and on blood proteins of Crangon vulgaris (Fabricius) [J]. Comparative Biochemistry & Physiology, 1970, 32(4): 709—718
[31] Yildirim M, Lim C, Wan P J, et al. Growth performance and immune response of channel catfish (Ictalurus punctatus) fed diets containing graded levels of gossypol-acetic acid [J]. Aquaculture, 2003, 219(1): 751—768
[32] 赵娟. 军曹鱼(Rachycentron canadum)对几种动植物蛋白源的利用研究. 广东海洋大学. 2011 Zhao J. Study on utilization of several animal and plant protein sources for juvenile cobia, Rachycentron canadum [D]. Guangdong Ocean University. 2011
赵娟. 军曹鱼(Rachycentron canadum)对几种动植物蛋白源的利用研究. 广东海洋大学. 2011[33] 刘襄河, 叶继丹, 王子甲, 等. 饲料中豆粕替代鱼粉比例对牙鲆生长性能及生化指标的影响. 水产学报, 2010, 34(3): 450—458 Liu X H, Ye J D, Wang Z J, et al. Partial replacement of fish meal by soybean in diets for juvenile Japanese flounder (paralichthys olivaceus) [J]. Journal of Fisheries of China, 2010, 34(3): 450—458
刘襄河, 叶继丹, 王子甲, 等. 饲料中豆粕替代鱼粉比例对牙鲆生长性能及生化指标的影响. 水产学报, 2010, 34(3): 450—458[34] 黄旭雄, 罗词兴, 郭腾飞, 等. 对虾Toll受体及其在虾类营养免疫评价中的应用. 水产学报, 2012, 36(6): 930—936 Huang X X, Luo C X, Guo T F, et al. Toll receptor in prawn and its application in nutrition-immunity assessing on prawn [J]. Journal of Fisheries of China, 2012, 36(6): 930—936
黄旭雄, 罗词兴, 郭腾飞, 等. 对虾Toll受体及其在虾类营养免疫评价中的应用. 水产学报, 2012, 36(6): 930—936[35] Sturve J, Almroth B C, Förlin L. Oxidative stress in rainbow trout (Oncorhynchus mykiss) exposed to sewage treatment plant effluent [J]. Ecotoxicology & Environmental Safety, 2008, 70(3): 446—452
[36] Bai S C, Kyeongjun L, Wilson R P. Different levels of dietary DL-α-tocopheryl acetate affect the vitamin E status of juvenile Korean rockfish, Sebastes schlegeli [J]. Aquaculture, 1998, 161(1): 405—414
[37] Han H J, Kim D H, Lee D C, et al. Pathogenicity of Edwardsiella tarda to olive flounder, Paralichthys olivaceus (Temminck & Schlegel) [J]. Journal of Fish Diseases, 2006, 29(10): 601—609
[38] 刘雪兰, 余为一. 甲壳动物免疫因子的研究进展. 水生生物学报, 2003, 27(4): 418—421 Liu X L, Yu W Y. A review on immune factors of crustaceans [J]. Acta Hydrobiologica Sinica, 2003, 27(4): 418—421
刘雪兰, 余为一. 甲壳动物免疫因子的研究进展. 水生生物学报, 2003, 27(4): 418—421[39] Asea A, Kraeft S K, Kurt-Jones E A, et al. HSP70 stimulates cytokine production through a CD14-dependant pathway, demonstrating its dual role as a chaperone and cytokine [J]. Nature Medicine, 2000, 6(4): 435—42
[40] Gay N J, Keith F J. Toll and IL-1 receptor [J]. Nature, 1991, 351(6325): 355—356
[41] May M J, Ghosh S. NF- kappaB and Rel proteins: Evolutionarily conserved medators of immune responses [J]. Immunology Today, 1998, 16: 225—260
-
期刊类型引用(20)
1. 米玮洁,张舜,胡俊,高少波,胡菊香. 黄河内蒙古河段水文情势变化及生态效应初探. 水生态学杂志. 2024(01): 42-49 .  百度学术
百度学术
2. 班学君,樊博,刘瀚,俞立雄,林俊强,夏继红,张迪. 鱼类产卵行为与生态水文指标响应关系研究——以长江四大家鱼为例. 水生态学杂志. 2024(01): 67-74 . 百度学术
3. 李莎,姜伟,李博,党莹超,苏巍,戴凌全. 长江葛洲坝坝下江段鱼类群落结构及多样性. 水生态学杂志. 2024(03): 121-130 . 百度学术
4. 张云昌,丁洋,赵进勇. 河流伦理建构中的八个理论和实践问题探讨. 中国水利. 2024(16): 7-11 . 百度学术
5. 易燃,陶江平,杨志,刘宏高,徐薇,曹俊,张鹏. 改进的栖息地法求解多种产漂流性卵鱼类生态流量方法研究. 中国农村水利水电. 2024(09): 45-51+59 . 百度学术
6. 蒋祥龙,黎明政,杨少荣,林鹏程,常涛,王春伶,张晨,高欣. 鄱阳湖鱼类多样性的时空变化特征研究. 水生生物学报. 2023(03): 376-388 . 本站查看
7. 周月娟,高雷,段辛斌,姜伟,黄安阳,王珂,陈大庆,汪登强. 基于耳石微结构的长江监利断面草鱼和鲢仔鱼群体日龄结构研究. 长江流域资源与环境. 2023(02): 324-330 . 百度学术
8. 徐薇,金瑶,陈桂亚,董纯,易燃,赵娜,陶江平. 三峡水库十年生态调度(2011—2020年)期间下游沙市江段产漂流性卵鱼类自然繁殖变化. 湖泊科学. 2023(05): 1729-1741 . 百度学术
9. 高雷,郭国忠,杨浩,陈大庆,王珂,段辛斌. 长江中游洪湖江段四大家鱼早期资源及生态调度影响. 中国水产科学. 2023(12): 1556-1565 . 百度学术
10. 高欣,赵亚辉,田菲,王晓爱,黎明政,林鹏程,常涛,俞丹,刘焕章. 中国内陆水体鱼类多样性监测专项网的监测和研究进展. 生物多样性. 2023(12): 199-210 . 百度学术
11. 霍堂斌,宋聃,刘伟,唐富江,王继隆,张澜澜,李喆,赵荣伟. 松花江下游富锦江段鱼类早期资源状况. 中国水产科学. 2022(01): 91-101 . 百度学术
12. 彭云鑫,方弟安,徐东坡,丁隆强,黎加胜,任鹏,何晓辉,罗宇婷. 长江安庆段草鱼仔稚鱼资源时空动态特征与水文因子的相关性. 中国水产科学. 2022(02): 314-324 . 百度学术
13. 蒋祥龙,黎明政,杨少荣,林鹏程,王春伶,张晨,高欣. 鄱阳湖鱼类集合群落结构特征及其时间变化研究. 长江流域资源与环境. 2022(03): 588-601 . 百度学术
14. 李鹏程,荣义峰,杜浩,王普渊,刘文成,刁亚芹. 基于深度学习的青海湖裸鲤产卵场遥感识别方法. 中国水产科学. 2022(03): 398-407 . 百度学术
15. 王小豪,方弟安,孙海博,何苗,薛向平. 长江如皋江段仔稚鱼资源现状调查分析. 南方农业学报. 2022(06): 1742-1751 . 百度学术
16. 向浩,林枫,伍遇普,田波,蔡志宇,吴益平,黄盼君,刘千凡,周志国. 长江中游宜昌段浮游动物群落结构特征与环境因子相关性分析. 黑龙江水产. 2022(05): 3-12 . 百度学术
17. 汤正阳,蒋爱萍,林俊强,任玉峰,张迪,刘新波,杨靖,赵庆绪. 长江流域水工程生态环境保护研究进展. 水利水电技术(中英文). 2022(S2): 297-309 . 百度学术
18. 祁涛,班璇,杜浩,郭文献,杜耘,龙奥运,南璐艺,石小涛,郑楚洋. 长江中游鱼类敏感水文期内水文情势对其资源量的影响评估. 长江流域资源与环境. 2022(12): 2621-2632 . 百度学术
19. 胡春明,董熙,尤立,李曜. 红花尔基水利枢纽对下游河道水文情势影响研究. 干旱区资源与环境. 2021(06): 89-95 . 百度学术
20. 王慧,张建新,田翊霏,陈君,耿硕,杨元昊,王立新. 湑水河国家级水产种质资源保护区鱼类资源调查与评价. 家畜生态学报. 2021(11): 54-59 . 百度学术
其他类型引用(6)





计量
- 文章访问数: 2108
- HTML全文浏览量: 580
- PDF下载量: 91
- 被引次数: 26