
THALASSIOSIRA ALLENII VAR. STRIATA AS A NEW VARIETY IN THE GENUS THALASSIOSIRA CLEVE
-
摘要: 为了澄清海链藻属Thalassiosira的物种多样性, 采用毛细管复洗技术建立了单克隆培养株系。利用光镜和扫描电镜观察形态学特征, 并扩增其核糖体小亚基和大亚基序列, 用于分子系统树的构建。结合形态学和分子系统学数据, 发现艾伦海链藻株系之间存在一定的形态差异和遗传多样性。经过与原始文献的比对, 确认了艾伦海链藻原变种的特征, 并报道了该种的一个新变种艾伦海链藻肋纹变种Thalassiosira allenii var. striata X. H. Guo, Y. Q. Guo & Y. Li。该变种与原变种的形态特征基本相似, 区别在于壳面边缘具有肋纹结构, 原变种则无。基于核糖体小亚基和大亚基的系统树均显示, 肋纹变种与原变种聚在同一个分支上, 形成姐妹支(BPP>0.90), 表明两者之间具有最近的亲缘关系。2个变种的核糖体小亚基序列完全一致, 没有碱基差异。但用于分析的556个核糖体大亚基序列中, 两者存在11个差异碱基, 遗传距离为0.01。Abstract: Thalassiosira Cleve is one of the most speciose marine diatom genera, that has been globally distributed. To assess the diversity of the genus Thalassiosira species, monoclonal strains were isolated and established from Chinese coast. The morphology was observed by using a light microscope (LM) and a scanning electron microscope (SEM). Sequences of the large subunit (LSU) and small subunit (SSU) of the nuclear rRNA-encoding genes (rDNA) were obtained to form a phylogenetic relationship. Among 14 established Thalassiosira allenii (T. allenii) Takano strains, morphological variation was found involving some with and without marginal ribs on the valve edge. After comparing with the original description of T. allenii, the strains without marginal ribs were assigned as a type of T. allenii, and those with marginal ribs were proposed as a new variety, T. allenii var. striata. Morphological divergence was supported by molecular data as well. On the phylogenetic trees inferred from both LSU and SSU of the nuclear rDNA, accordingly, T. allenii was clustered with var. striata, and formed as sister groups (Bayesian posterior probability (BPP) > 0.90), demonstrating their firm relationship. SSU sequences are totally identical, without any different base pair, between these two varieties. However, 11 different base pairs among 556 LSU base pair, which used for analysis, could be found, and divergence of distance among them is only 0.01.
-
Keywords:
- Thalassiosira allenii var. striata /
- Morphology /
- Phylogen /
- Thalassiosira allenii
-
海链藻属(Thalassiosira Cleve)物种是一类常见的海洋浮游植物, 是海洋微型浮游植物的重要组成部分, 海洋初级生产力的重要贡献者, 广泛分布于全球各类水体[1], 有些物种也是近岸海域常见的藻华生物[2]。海链藻属是硅藻门中物种多样性最为丰富的几个大属之一, 已报道270多个分类单位[3], 而我国目前仅记录50余种[4, 5]。相对于我国辽阔的近岸海域而言, 以往研究还相对有限, 仍有较为丰富的物种未被发现和认知。
艾伦海链藻T. allenii Takano是近岸广布种, 全球多个海域均有报道[1, 6]。以往研究认为, 艾伦海链藻部分特征较不稳定, 存在变化的情况, 主要体现在壳面孔纹排列方式、壳缘唇形突的分布位置等[7]。目前该种被广泛认可的特征为: 壳面孔纹六角形或多边形, 壳套具有2—4排孔纹, 具有1个中央支持突和1环壳缘支持突, 有1个唇形突, 位于1环壳缘支持突内[7]。除上述较为稳定的特征之外, (不具有)壳缘肋纹也曾被认为是稳定特征, 可用于区别艾伦海链藻及其相似物种[7, 8]。
为了丰富海链藻属的物种多样性, 作者陆续从我国海域建立了海链藻的单克隆培养株系, 对其形态特征进行观察, 并对核糖体目的基因进行扩增, 用于系统学分析。在建立的14个艾伦海链藻株系中, 发现了细微的形态差异, 并对应一定的遗传多样性。经过与原始文献的比对, 确认了艾伦海链藻原变种的特征, 并发现该种的一个新变种, 艾伦海链藻肋纹变种T. allenii var. striata X. H. Guo, Y. Q. Guo & Y. Li。
1. 材料与方法
1.1 单克隆培养株系的建立
利用浮游植物网(孔径10 μm)进行水平拖网, 采集浮游植物活体样品, 尽快带回实验室进行株系的分离和纯化。在生物倒置显微镜(Mshot MI-12)下, 利用毛细管复洗技术分离单个细胞或单条链状群体, 转移至装有L培养基的48孔细胞培养板内, 置于室内(22±2)℃培养。待细胞个数达到一定数量后(100个以上), 转移到盛有L培养液的100 mL三角锥形瓶中继续扩大培养。
本文建立了14个艾伦海链藻单克隆株系, 详见表 1。
表 1 艾伦海链藻单克隆株系信息Table 1. List of T. allenii monoclonal strains established in this study株系
Strain code分离地点
Isolation location采样时间
Sampling data采集人
CollectorMC308 墨西哥 2015.03 郭雅琼 MC309 墨西哥 2015.03 郭雅琼 MC334 香港 2015.03 郭雅琼 MC346 琼海 2015.04 郭雅琼 MC519 湛江 2015.09 郭雅琼 MC533 青岛 2015.09 郭雅琼 MC545 青岛 2015.09 郭雅琼 MC549 青岛 2015.09 郭雅琼 MC558 大连 2015.09 郭雅琼 MC561 厦门 2015.09 郭雅琼 MC566 厦门 2015.09 郭雅琼 MC598 香港 2015.10 郭雅琼 MC1406 香港 2015.10 郭雅琼 MC1476 香港 2015.11 郭雅琼 1.2 形态学观察
光学显微镜(Light microscopy, LM)观察: 吸取0.1 mL处于对数生长期的藻液, 滴于载玻片上并加盖盖玻片, 利用光学显微镜(Olympus BX53)进行微分干涉(Differential interference contrast, DIC)观察, 并使用Olympus DP27数码相机拍照, 在Olympus CellSens软件上获取图像信息。主要观察特征为群体和色素体。
扫描电镜(Scanning electron microscopy, SEM)观察: 吸取1 mL处于对数生长期的藻液, 转移至试管中, 加入等体积的浓硫酸或者浓盐酸, 静置15min左右, 具体时间可根据有机质含量的多少而定, 酸化以去除有机质[9]。然后加入灭菌水, 静置24h, 吸弃上清, 再次加入灭菌水, 如此多次水洗至中性。用20 mL注射器吸取适量酸化后的样品, 用孔径3 μm的Millipore滤膜过滤, 待滤膜自然晾干后, 用碳导电胶贴于SEM样品铜台上, 溅射仪喷金后即可置于扫描电镜(Zeiss Ultra 55)下进行观察和拍照。主要观察特征有突起的类型和结构、壳面和壳套处孔纹、壳缘肋纹结构等。
1.3 分子系统学分析
采用离心法收集对数生长期的藻细胞, 提取总DNA[10]。利用正向引物SSU-F和反向引物SSU-R[11]扩增核糖体小亚基编码基因(Small subunit ribosomal encoding gene, SSU)的序列, 利用正向引物D1R-F[12]和反向引物D3B-R[13]扩增核糖体大亚基编码基因(Large subunit ribosomal encoding gene, LSU)的D1-D3区序列。PCR产物送上海立菲生物科技有限公司进行纯化和测序。从美国国立生物技术信息中心(NCBI)下载艾伦海链藻的序列信息, 及其形态学相似种、遗传学相似种的SSU和LSU序列信息。运用BioEdit软件进行序列的比对和矩阵[14]。基于MrModeltest 2.3计算[15], 选择最适的模型和参数, 然后分别用RAxML-HPC2[16]和MrBayes 3.2[17]构建最大似然树(Maximum likelihood, ML)和贝叶斯系统发育树(Bayesian inference, BI)。其中ML分析的自检值(Bootstrap test)设定为1000。
2. 结果
2.1 基于形态学特征的初步鉴定
经过光镜和扫描电镜的观察, 发现14个艾伦海链藻株系的形态特征基本一致, 但也存在细微的形态差异, 主要表现为壳缘肋纹的有无, 可据此将实验株系划分为两个小类群, 艾伦海链藻类群一(T. allenii group I, 无壳缘肋纹结构)和艾伦海链藻类群二(T. allenii group II, 有壳缘肋纹结构)。下面将分别对两个小类群的形态特征进行简要描述。
艾伦海链藻类群一: 该类群的形态特征与Takano [18]、Hasle [8]、郭雅琼等[7]记录完全一致。细胞可形成短链群体。壳面直径5.8—16.6 μm。壳面孔纹呈切线状或偏心状排列。壳套高2—4排孔纹。具有1个中央支持突和1个壳缘唇形突。艾伦海链藻类群一包括6个株系, MC334、MC346、MC545、MC558、MC598和MC1406。
艾伦海链藻类群二: 类群二与类群一的形态特征基本一致, 主要区别是壳缘处有肋纹结构, 光镜下亦可见, 密度为每10 μm内30—40条。艾伦海链藻类群二包括8个株系, MC308、MC309、MC519、MC533、MC549、MC561、MC566和MC1476。
2.2 基于核糖体基因的系统学分析
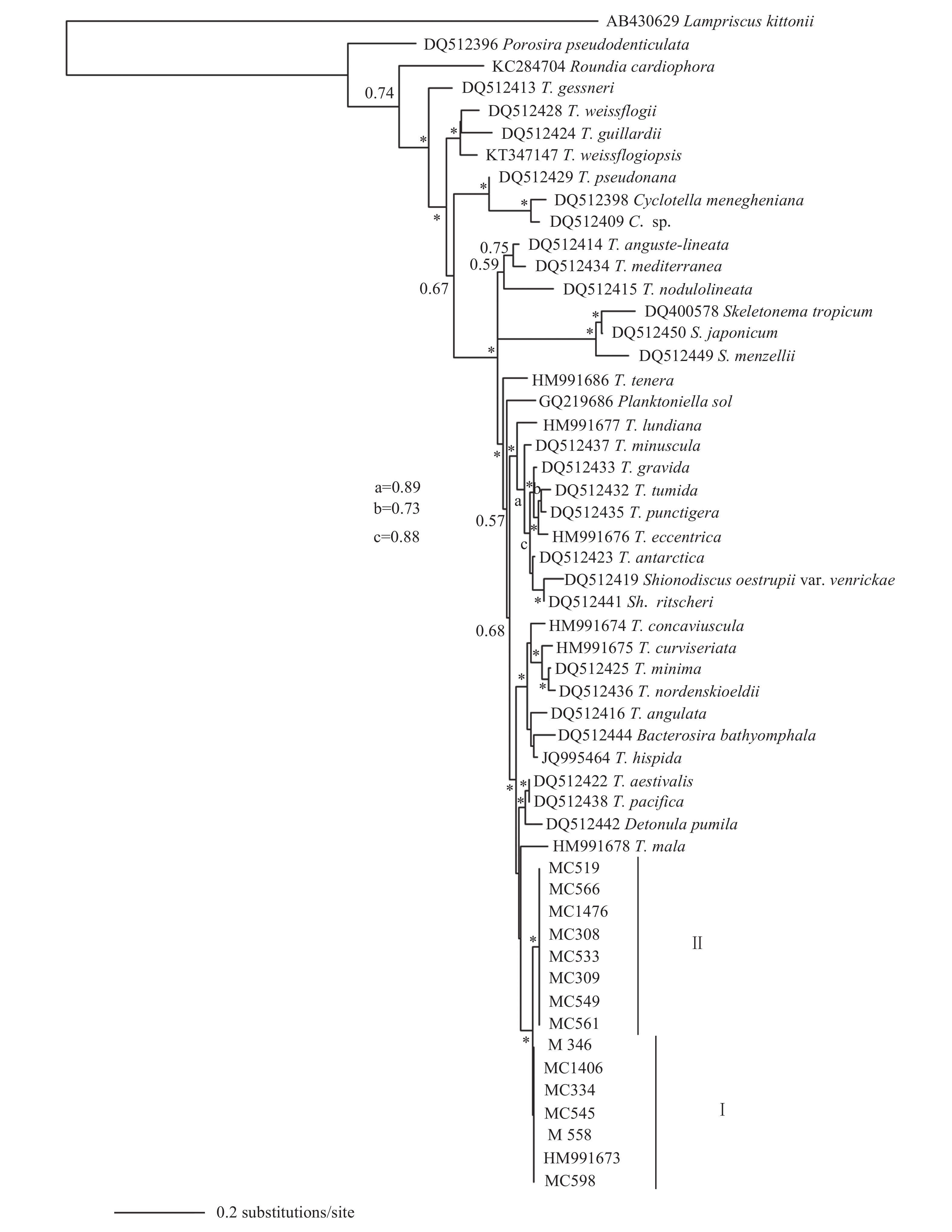
经过SSU序列比对, 用于最后分析的1753个SSU序列碱基中, 本文建立的艾伦海链藻株系(MC309、MC346、MC533、MC558、MC566、MC1406和MC1476)与韩国株系HM991688具有完全相同的碱基序列, 不存在任何差异碱基。从SSU系统发育树(图 1)中可见, 类群一与类群二聚在同一个分支上, 显示2个类群之间最为密切的亲缘关系。
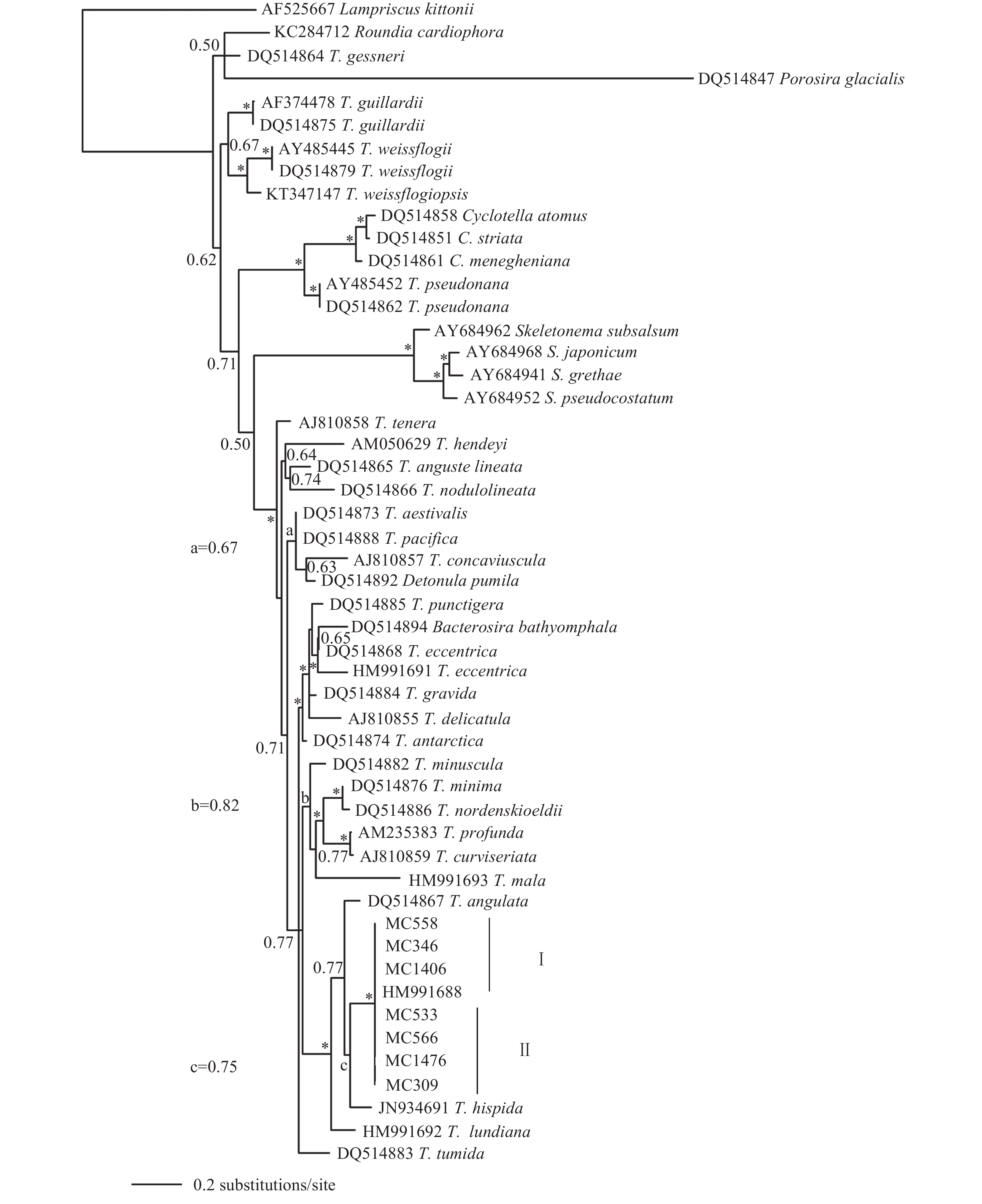
LSU rDNA系统发育树(图 2)显示, 类群一与类群二聚在一起(BPP>0.90), 形成姐妹分支, 类群划分与形态聚类结果相互支持。HM991673、MC334、MC346、MC545、MC558、MC598和MC1406藻株具有完全相同的LSU序列, MC308、MC309、MC519、MC533、MC549、MC561、MC566和MC1476藻株具有完全相同的LSU序列。用于最后分析的556个LSU碱基中, 2个类群之间有11个差异碱基, 遗传距离为0.01。
2.3 新变种确认
基于2个类群之间较为广泛的相似形态学特征, 及其完全一致的SSU序列信息和分子系统学位置(BPP>0.90), 我们认为本文建立的14个株系均属于艾伦海链藻。但是细微的形态学差异, 以及对应的LSU系统学关系, 均显示艾伦海链藻内部存在2个小类群的划分, 说明存在隐藏的多样性, 那么哪个是真正的艾伦海链藻呢?
经过与艾伦海链藻原始文献的比对, 我们发现原始文献并未提及壳缘肋纹的存在, 提供的图版中也没有显示肋纹结构, 因此我们确认了类群一是典型的艾伦海链藻T. allenii。类群二与典型艾伦海链藻的形态特征基本一致, 区别仅在于壳缘肋纹。考虑到该特征在本文建立的8个株系均能稳定出现, 以及类群二株系具有完全一致的SSU和LSU序列, 我们认为类群二可以作为一个独立的分类学单位。另外, 鉴于2个类群具有完全一致的SSU序列, 以及LSU系统学较小的遗传距离(0.01), 我们建议将类群二作为1个新的变种, 艾伦海链藻肋纹变种T. allenii var. striata。
![]() 图 1 基于核糖体小亚基SSU nrDNA构建的贝叶斯系统树(置信值大于0.90用星号显示)Figure 1. Molecular phylogenetic tree inferred from sequences of SSU of the nuclear rDNA (Bayesian posterior probability values greater than 0.90 are shown with an asterisk)
图 1 基于核糖体小亚基SSU nrDNA构建的贝叶斯系统树(置信值大于0.90用星号显示)Figure 1. Molecular phylogenetic tree inferred from sequences of SSU of the nuclear rDNA (Bayesian posterior probability values greater than 0.90 are shown with an asterisk)![]() 图 2 基于核糖体大亚基LSU nrDNA D1-D3构建的贝叶斯系统树(置信值大于0.90用星号显示)Figure 2. Molecular phylogenetic tree inferred from sequences of LSU of the nuclear rDNA (values of BPP greater than 0.90 are shown with an asterisk*)
图 2 基于核糖体大亚基LSU nrDNA D1-D3构建的贝叶斯系统树(置信值大于0.90用星号显示)Figure 2. Molecular phylogenetic tree inferred from sequences of LSU of the nuclear rDNA (values of BPP greater than 0.90 are shown with an asterisk*)艾伦海链藻肋纹变种Thalassiosira allenii var. striata X. H. Guo, Y. Q. Guo & Y. Li var. nov. ( 图版Ⅰ—Ⅱ)
Formal diagnosis: This variety shares the main morphology with var. allenii, and is distinguished by marginal ribs at the valve edge.
Type locality: Jiaozhou Bay (36°07′32.3″N, 120°24′31.6″E), Qingdao, Shandong Province, Yellow Sea, P. R. China.
Holotype: Permanent slide of acid-washed material of strain MC533, deposited at the Marine Diatoms Collection of South China Normal University, with number assigned as PS36.
Isotype: Glutaraldehyde-fixed material of strain MC533, deposited at the Marine Diatoms Collection of South China Normal University, China, catalog number F-MC533.
Live strain: MC533 deposited in the Laboratory of Phycology, School of Life Science, South China Normal University, China.
Habitat: Marine, planktonic.
Known geographic distribution: Dalian, Qingdao, Xiamen, Zhanjiang, Qionghai, Hong Kong, P. R. China; Mexico coast of Pacific.
Etymology: The name refers to the structure of marginal ribs at the valve edge.
细胞单独生活, 或形成两个细胞的短链状群体(图版Ⅰ-1—3)。细胞壳面呈圆盘形, 直径11—25 μm, 内含椭圆形或不定型的色素体(图版Ⅰ-2)。细胞环面观近鼓形, 贯壳轴7—10 μm, 壳面微微隆起, 壳缘可见肋状结构(图版Ⅰ-3)。壳面孔纹切线状排列(图版Ⅰ-4—6), 10 μm内11—22个。孔纹为典型的具筛室孔纹, 在外壳面呈圆形的中央孔, 在内壳面具有独立筛板膜。中央孔内具有放射状硅质肋(图版Ⅰ-6)。壳面孔纹大小基本相似, 壳套处孔纹较小。壳套高2—4排孔纹, 10 μm内21—32个。壳面分布有硅质毛状物(图版Ⅰ-6, 图版Ⅱ-3)。壳面近中央处具有1个支持突, 不具有外管结构(图Ⅰ-6), 基部有4个围孔(图版Ⅰ-7)。外壳面可见与之相邻的1个中央较大孔纹, 有些壳面明显(图版Ⅰ-4), 有些壳面不明显(图版Ⅰ-6)。具有1圈壳缘支持突(图版Ⅰ-4、图版Ⅱ-2), 有明显的外管结构, 基部有4个围孔(图版Ⅱ-4), 密度为每10 μm内6—9个。壳缘处有1个唇形突(图版Ⅱ-1、2), 比壳缘支持突稍微远离壳缘, 位于两个壳缘支持突间(图版Ⅱ-1、2、5)。唇形突的外管明显(图版Ⅱ-1、5)。壳缘处有肋纹结构(图版Ⅱ-1、3、5), 光镜下亦可见(图版Ⅰ-3), 密度为每10 μm内30—40条。
上壳环由6个开放环带组成(图版Ⅱ-5、6)。壳环带靠近壳面区域有5—6行不规则排列的孔纹, 远离壳面区域为无孔区, 约占壳环带宽度的五分之二。第2条环带靠近壳环带处有一行规则的较大孔纹, 密度是每10 μm内67—70个。其余4条环带光滑, 未见孔纹结构。
生境: 海洋浮游生活。
分布: 标本采自湛江(9月)、青岛(9月)、厦门(9月)以及香港(11月), 墨西哥锡那罗亚(3月)也有分布。
![]() 图版Ⅰ 艾伦海链藻肋纹变种1—3. LM; 4—7. SEM. 1. 群体; 2. 壳面观; 3. 环面观; 4. 外壳面观; 5. 内壳面观; 6. 中央支持突外壳面观; 7. 中央支持突内壳面观. 图1—3, 标尺=20 μm; 图4、5, 标尺=4 μm; 图6、7, 标尺=1 μm图版Ⅰ. Thalassiosira allenii var. striata1—3. LM; 4—7. SEM. 1. Colonies; 2. Valve view; 3. Girdle view; 4. External valve view; 5. Internal valve view; 6. Central fultoportula on external valve; 7. Central fultoportula on internal valve. Fig. 1—3, scale bars=20 μm; Fig. 4, 5, scale bars=4 μm; Fig. 6, 7, scale bars=1 μm
图版Ⅰ 艾伦海链藻肋纹变种1—3. LM; 4—7. SEM. 1. 群体; 2. 壳面观; 3. 环面观; 4. 外壳面观; 5. 内壳面观; 6. 中央支持突外壳面观; 7. 中央支持突内壳面观. 图1—3, 标尺=20 μm; 图4、5, 标尺=4 μm; 图6、7, 标尺=1 μm图版Ⅰ. Thalassiosira allenii var. striata1—3. LM; 4—7. SEM. 1. Colonies; 2. Valve view; 3. Girdle view; 4. External valve view; 5. Internal valve view; 6. Central fultoportula on external valve; 7. Central fultoportula on internal valve. Fig. 1—3, scale bars=20 μm; Fig. 4, 5, scale bars=4 μm; Fig. 6, 7, scale bars=1 μm![]() 图版Ⅱ 艾伦海链藻肋纹变种1—6. SEM. 1. 外壳面唇形突和壳缘支持突; 2. 内壳面唇形突; 3. 壳缘肋纹; 4. 内壳面支持突; 5. 环面观及壳套; 6. 环带. 图1—6, 标尺=1 μm图版Ⅱ. Thalassiosira allenii var. striata1—6. SEM. 1. Rimoportula and marginal fultoportulae on external vavle; 2. Rimoportula on internal valve; 3. Marginal ribs; 4. Fultoportulae on internal valve; 5. Girdle view and valve mantle; 6. Enlargement of the band. Fig. 1—6, scale bars=1 μm
图版Ⅱ 艾伦海链藻肋纹变种1—6. SEM. 1. 外壳面唇形突和壳缘支持突; 2. 内壳面唇形突; 3. 壳缘肋纹; 4. 内壳面支持突; 5. 环面观及壳套; 6. 环带. 图1—6, 标尺=1 μm图版Ⅱ. Thalassiosira allenii var. striata1—6. SEM. 1. Rimoportula and marginal fultoportulae on external vavle; 2. Rimoportula on internal valve; 3. Marginal ribs; 4. Fultoportulae on internal valve; 5. Girdle view and valve mantle; 6. Enlargement of the band. Fig. 1—6, scale bars=1 μm3. 讨论
3.1 艾伦海链藻肋纹变种与原变种的比较研究
在形态学特征上, 肋纹变种与原变种基本一致, 具有1个中央支持突、1个中央较大孔纹、1个壳缘唇形突和1圈壳缘支持突, 以及切线或偏心状排列的壳面孔纹等。两者之间的形态相似性亦获得分子系统学的支持, 无论是保守的SSU系统树, 还是相对高变的LSU系统树, 肋纹变种与原变种的株系均聚在一起(BPP>0.90), 形成姐妹分支, 显示两者之间最为紧密的亲缘关系。两者之间的区别特征仅仅体现在壳缘肋纹的有无。细微的形态差异得到相对高变的LSU系统树的验证, 两者虽然形成姐妹分支, 但仍存在一定的遗传差异。鉴于2个类群之间较多的形态相似性, 以及完全一致的SSU序列, 可以确认它们之间紧密的遗传关系。同时鉴于肋纹特征的稳定性, 以及2个类群之间较小的LSU分化和较近的遗传距离(0.01), 将其划分为同一物种下的两个变种是较为合适的。
3.2 肋纹结构在海链藻的分类学探讨
在海链藻属物种的研究中, 壳缘肋纹曾被作为重要分类学依据, 例如结线形海链藻(T. nodulolineata)、细孔海链藻(T. punctigera)、伦德海链藻(T. lundiana)和安达曼海链藻(T. andamanica)等均具有稳定的壳缘肋纹结构[1, 8, 19—21]。此外, 也有一些海链藻物种不具有壳缘肋纹结构, 也被认为是稳定特征, 例如成对海链藻(T. binata)、小字海链藻(T. minuscula)和脆弱海链藻(T. fragilis)等[1, 22, 23]。Hasle[8]曾以壳缘肋纹来区分艾伦海链藻及相似物种, 郭雅琼等[7]也认为壳缘肋纹结构可作为海链藻物种之间的区别特征。以往报道的艾伦海链藻均无壳缘肋纹结构, 而本文发现的新变种具有壳缘肋纹结构, 与原变种之间有明显的形态学与遗传学差异。这个结果也进一步验证了壳缘肋纹结构是稳定特征, 可以作为海链藻物种的鉴定依据。
3.3 艾伦海链藻肋纹变种及其相似种的比较研究
除了艾伦海链藻原变种之外, 还有一些物种具有与艾伦海链藻肋纹变种相似的形态学特征, 如夏眠海链藻(T. aestivalis)、棱角海链藻(T. angulata)、诺氏海链藻(T. nordenskioeldii)、太平洋海链藻(T. pacifica)以及具毛海链藻(T. hispida)。它们的共有特征是具有1个中央支持突, 1个壳缘唇形突和1圈壳缘支持突。本文对它们的形态学特征进行了详细的比较(表 2)。壳缘唇形突的位置可作为艾伦海链藻肋纹变种与其他相似种的显著区别特征, 相比之下, 该变种的壳缘唇形突更远离壳缘, 而其他相似种的壳缘唇形突则位于1圈壳缘支持突内, 或者较靠近壳缘支持突的内侧。另外, 该变种与具毛海链藻的区别在于壳缘肋纹的有无, 与夏眠海链藻、诺氏海链藻、太平洋海链藻的区别在于壳缘肋纹的密度、壳面孔纹排列方式。
表 2 艾伦海链藻肋纹变种及其形似种的形态学比较Table 2. Morphological comparison of T. allenii var. striata and allied species物种
Species直径
Diameter
(μm)壳面孔纹
Areolae壳套高度(孔纹)
Mantle height
(Areolae)壳缘支持突
Marginal
fultoportula
(ind./10 μm)唇形突位置
Location of
rimoportula壳缘肋纹
Marginal ribs
(ind./10 μm)参考文献
Reference排列方式
Pattern壳面密度Density (ind./10 μm) T. allenii var. allenii 5—20 切线状 束状 18—24 2—4 5—7 多变, 主要是占据1个壳缘支持突 无 Hasle[8] T. allenii var. striata 11—25 切线状 11—22 2—3 6—9 位于壳缘支持突间, 但比壳缘支持突远离壳缘 30—40 本文 T. aestivalis 14—56 束状 18 2—3 3—5 占据1个壳缘支持突 14 Hasle[8] T. angulata 12—39 切线形 8—18 2—4 2—4 紧邻1个壳缘支持突 14—24 Hasle[8] T. nordenskioldii 10—50 放射状 14—18 6—8 3 多变, 位于2个壳缘支持突间 18—20 Hasle[8] T. pacifica 7—46 束状 10—14 2—3 4—6 占据1个壳缘支持突 5—7 Hasle[8] T. hispida 5—22 束状 18 3 5 2个壳缘支持突间 无 Syvertsen[24] T. mala 5—7 辐射状 螺旋状 30—60 / 13—19 2个壳缘支持突间 / 程兆第等[9] 具毛海链藻、夏眠海链藻、太平洋海链藻和棱角海链藻是肋纹变种的形态相似种, 也是系统学近缘种。从SSU rDNA系统发育树(图 1)中可见, 艾伦海链藻的两个变种与棱角海链藻和具毛海链藻聚在一起, 但是支持率并不高(BPP=0.75, 0.77)。在LSU rDNA系统发育树(图 2)上, 艾伦海链藻的2个变种与夏眠海链藻和太平洋海链藻聚在相邻的小分支上, 支持率也不高(均低于0.50)。LSU rDNA系统发育树(图 2)显示, 艾伦海链藻的两个变种与萎软海链藻(T. mala)聚在同一个小分支上并具有较高的支持率(BPP>0.90), 但两者的形态学区别较为明显(表 2), 后者常形成胶质块状群体, 并且唇形突远离壳缘, 位于壳面, 这些特征均未见于艾伦海链藻肋纹变种。
-
![]()
图 1 基于核糖体小亚基SSU nrDNA构建的贝叶斯系统树(置信值大于0.90用星号显示)
Figure 1. Molecular phylogenetic tree inferred from sequences of SSU of the nuclear rDNA (Bayesian posterior probability values greater than 0.90 are shown with an asterisk)
![]()
图 2 基于核糖体大亚基LSU nrDNA D1-D3构建的贝叶斯系统树(置信值大于0.90用星号显示)
Figure 2. Molecular phylogenetic tree inferred from sequences of LSU of the nuclear rDNA (values of BPP greater than 0.90 are shown with an asterisk*)
![]()
图版Ⅰ 艾伦海链藻肋纹变种
1—3. LM; 4—7. SEM. 1. 群体; 2. 壳面观; 3. 环面观; 4. 外壳面观; 5. 内壳面观; 6. 中央支持突外壳面观; 7. 中央支持突内壳面观. 图1—3, 标尺=20 μm; 图4、5, 标尺=4 μm; 图6、7, 标尺=1 μm
图版Ⅰ. Thalassiosira allenii var. striata
1—3. LM; 4—7. SEM. 1. Colonies; 2. Valve view; 3. Girdle view; 4. External valve view; 5. Internal valve view; 6. Central fultoportula on external valve; 7. Central fultoportula on internal valve. Fig. 1—3, scale bars=20 μm; Fig. 4, 5, scale bars=4 μm; Fig. 6, 7, scale bars=1 μm
![]()
图版Ⅱ 艾伦海链藻肋纹变种
1—6. SEM. 1. 外壳面唇形突和壳缘支持突; 2. 内壳面唇形突; 3. 壳缘肋纹; 4. 内壳面支持突; 5. 环面观及壳套; 6. 环带. 图1—6, 标尺=1 μm
图版Ⅱ. Thalassiosira allenii var. striata
1—6. SEM. 1. Rimoportula and marginal fultoportulae on external vavle; 2. Rimoportula on internal valve; 3. Marginal ribs; 4. Fultoportulae on internal valve; 5. Girdle view and valve mantle; 6. Enlargement of the band. Fig. 1—6, scale bars=1 μm
表 1 艾伦海链藻单克隆株系信息
Table 1 List of T. allenii monoclonal strains established in this study
株系
Strain code分离地点
Isolation location采样时间
Sampling data采集人
CollectorMC308 墨西哥 2015.03 郭雅琼 MC309 墨西哥 2015.03 郭雅琼 MC334 香港 2015.03 郭雅琼 MC346 琼海 2015.04 郭雅琼 MC519 湛江 2015.09 郭雅琼 MC533 青岛 2015.09 郭雅琼 MC545 青岛 2015.09 郭雅琼 MC549 青岛 2015.09 郭雅琼 MC558 大连 2015.09 郭雅琼 MC561 厦门 2015.09 郭雅琼 MC566 厦门 2015.09 郭雅琼 MC598 香港 2015.10 郭雅琼 MC1406 香港 2015.10 郭雅琼 MC1476 香港 2015.11 郭雅琼  下载: 导出CSV
下载: 导出CSV
表 2 艾伦海链藻肋纹变种及其形似种的形态学比较
Table 2 Morphological comparison of T. allenii var. striata and allied species
物种
Species直径
Diameter
(μm)壳面孔纹
Areolae壳套高度(孔纹)
Mantle height
(Areolae)壳缘支持突
Marginal
fultoportula
(ind./10 μm)唇形突位置
Location of
rimoportula壳缘肋纹
Marginal ribs
(ind./10 μm)参考文献
Reference排列方式
Pattern壳面密度Density (ind./10 μm) T. allenii var. allenii 5—20 切线状 束状 18—24 2—4 5—7 多变, 主要是占据1个壳缘支持突 无 Hasle[8] T. allenii var. striata 11—25 切线状 11—22 2—3 6—9 位于壳缘支持突间, 但比壳缘支持突远离壳缘 30—40 本文 T. aestivalis 14—56 束状 18 2—3 3—5 占据1个壳缘支持突 14 Hasle[8] T. angulata 12—39 切线形 8—18 2—4 2—4 紧邻1个壳缘支持突 14—24 Hasle[8] T. nordenskioldii 10—50 放射状 14—18 6—8 3 多变, 位于2个壳缘支持突间 18—20 Hasle[8] T. pacifica 7—46 束状 10—14 2—3 4—6 占据1个壳缘支持突 5—7 Hasle[8] T. hispida 5—22 束状 18 3 5 2个壳缘支持突间 无 Syvertsen[24] T. mala 5—7 辐射状 螺旋状 30—60 / 13—19 2个壳缘支持突间 / 程兆第等[9]
下载: 导出CSV
-
[1] Hasle G R, Syvertsen E E. Marine Diatoms [M]. San Diego: Academic Press. 1997, 5—387
[2] Fryxell G A, Hasle G R. Taxonomy of Harmful Diatoms [M]. Paris: Imprimerie Landais. 2004, 465—510
[3] Guiry M D, Guiry G M. AlgaeBase. World-wide electronic publication, National Unversitr of Ireland, Galway [EB/OL]. http://www.algaebase.org/
[4] Li Y, Zhao Q L, Lü S H. The genus Thalassiosira off the Guangdong coast, South China Sea [J]. Botanica Marina, 2013, 56(1): 83—110
[5] Li Y, Zhao Q L, Lü S H. Taxonomy and species diversity of the diatom genus Thalassiosira (Bacillariophyceae) in Zhejiang coastal waters, the East China Sea [J]. Nova Hedwigia, 2014, 99(3—4): 373—402
[6] Round F E, Crawford R M, Mann D G. The Diatomes: Biology and Morphology of the Genera [M]. Cambridge: Cambridge University Press. 1990, 747
[7] 郭雅琼, 吴归仪, 李扬. 海链藻属重要形态学特征的二次研究——以艾伦海链藻为例. 植物科学学报, 2017, 35(2): 194—204 Guo Y Q, Wu G Y, Li Y. Re-examination and assessment of the morphological traits of the diatom genus Thalassiosira Cleve, a case study on Thalassiosira alleni Takano [J]. Plant Science Journal, 2017, 35(2): 194—204
郭雅琼, 吴归仪, 李扬. 海链藻属重要形态学特征的二次研究——以艾伦海链藻为例. 植物科学学报, 2017, 35(2): 194—204[8] Hasle G R. Some Thalassiosira species with one central process (Bacillariophyceae) [J]. Norwegian Journal of Botany, 1978, 25(1): 77—110
[9] 程兆第, 高亚辉, 刘师成. 福建沿岸微型硅藻. 北京: 海洋出版社. 1993, 91 Cheng Z D, Gao Y H, Liu S C. Nano-diatoms in Fujian Coast [M]. Beijing: Ocean Press. 1993, 91
程兆第, 高亚辉, 刘师成. 福建沿岸微型硅藻. 北京: 海洋出版社. 1993, 91[10] Lundholm N, Daugbjerg N, Moestrup Ø. Phylogeny of the Bacillariaceae with emphasis on the genus Pseudo-nitzschia (Bacillariophyceae) based on partial LSU rDNA [J]. European Journal of Phycology, 2002, 37(1): 115—134
[11] Zhen Y, Mi T, Yu Z. Detection of Phaeocystis globosa using sandwich hybridization integrated with nuclease protection assay (NPA-SH) [J]. Journal of Environmental Sciences, 2008, 575(12): 1—11
[12] Scholin C A, Herzog M, Sogin M, et al. Identification of group-and strain-specific genetic markers for globally distributed alexandrium (dinophyceae). II. Sequence analysis of a fragment of the LSU rRNA gene [J]. Journal of Phycology, 1994, 30(6): 999—1011
[13] Nunn G B, Theisen B F, Christensen B, et al. Simplicity-correlated size growth of the nuclear 28S ribosomal RNA D3 expansion segment in the crustacean order Isopoda [J]. Journal of Molecular Evolution, 1996, 42(2): 211—223
[14] Hall T A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT [C]. Nucleic acids symposium series. London: Information Retrieval Ltd, c1979-c2000. 1999, (41): 95—98
[15] Nylander J A A. Mrmodeltest v2.3.Program distributed by the auther [CP]. Evolutionary Biology Centre, Uppsala University, Uppsala, Sweden, 2004. https://www.abc.se/~nylander/
[16] Miller M A, Pfeiffer W, Schwartz T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees [C]. Gateway Computing Environments Workshop (GCE), 2010. Ieee. 2010, 1—8
[17] Ronquist F, Teslenko M, Van Der Mark P, et al. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space [J]. Systematic Biology, 2012, 61(3): 539—542
[18] Takano H. New and rare diatoms from Japanese marine waters I. [J]. Bulletin of Tokai Regional Fisheries Research Laboratory, 1965, 42: 1—12
[19] Fryxell G A. Three new species of Thalassiosira, with observations on the occluded process, a newly observed structure of diatom valves [J]. Nova Hedwigia: Beih, 1975, 53(1—2): 57—73
[20] Hasle G R. The marine, planktonic diatoms Thalassiosira oceanica sp. nov. and Thalassiosira partheneia[J]. Journal of Phycology, 1983, 19(2): 220—229
[21] Hasle G R, Fryxell G A. genus Thalassiosira: some species with a linear areola array [J]. Nova Hedwigia, 1977, 54:15—66
[22] Hallegraeff G M. Species of the Diatom Genus Pseudonitzschia in Australian Waters: Botanica Marina [J]. Botanica Marina, 1984, 27(11): 495—514
[23] Herzig W N, Fryxell G A. The Diatom Genus Thalassiosira Cleve in Gulf Stream Warm Core Rings: Taxonomy, with T. intrannula and T. lineoides, spp. Nov [J]. Botanica Marina, 1986, 29(1): 11—26
[24] Syvertsen E E. Thalassiosira hispida sp. nov., a marine planktonic diatom [C]. Proceedings of the 8th International Diatom Symposium. Koenigstein, Koeltz. 1984, 33—42
-
期刊类型引用(1)
1. 郭晓慧,郭雅琼,李扬. 采自广东大亚湾水域的海链藻属一新种——扇形海链藻. 水生生物学报. 2019(05): 1132-1138 .  本站查看
本站查看
其他类型引用(1)


计量
- 文章访问数: 2190
- HTML全文浏览量: 559
- PDF下载量: 39
- 被引次数: 2