
THE DIFFERENT RESPONSE OF THE INTESTINAL MUCOSAL HISTOLOGICAL FEATURES IN DIFFERENT SECTIONS OF JUVENILE CHINESE SOFT-SHELLED TURTLE UNDER ACUTE COLD STRESS
-
摘要: 为了探究急性冷胁迫对中华鳖(Pelodiscus sinensis)幼鳖肠道不同区段黏膜组织学特征的影响, 实验检测了急性冷胁迫前后血清二胺氧化酶(Diamine oxidase, DAO)的活性, 同时观察了急性冷胁迫前后中华鳖肠道黏膜的相关组织形态的差异。DAO活性显示: (1)在第一次急性冷胁迫实验中, 中华鳖血清DAO活性随急性冷胁迫时间的增加而呈现降低趋势, 并在冷胁迫到达48h降到了最低水平; (2)在急性冷胁迫及复温实验中, 中华鳖血清DAO的活性, 在冷胁迫3d后显著降低, 但随着温度的恢复, DAO活性又恢复到正常水平。组织病理结果显示: (1)急性冷胁迫对中华鳖肠道(回肠后段和大肠)黏膜上皮的形态没有明显影响; (2)急性冷胁迫对回肠后段的杯状细胞数目、肠绒毛长度和绒毛长度/隐窝深度的比值没有显著影响, 但会使回肠后段黏膜厚度显著降低; (3)急性冷胁迫会使大肠的杯状细胞数目降低。这表明急性冷胁迫会改变中华鳖肠道黏膜的结构, 但在不同的肠段, 这种改变是不同的。回肠后段和大肠在同样的冷胁迫方式下黏膜机械屏障的不同变化情况, 提示中华鳖肠道各段对急性冷胁迫具有特殊的应对方式。Abstract: To investigate the effects of acute cold stress on the mucosal histology in different sections of the juvenile Pelodiscus sinensis, the activity of serum diamine oxidase (DAO) and histological features of intestinal mucosa in Chinese soft-shelled turtle in culture temperature and acute cold stress were conducted. The first acute cold stress reduced the DAO activity with the lowest level at 48h after cold stress. In the acute cold stress and rewarming experiment, the DAO activity decreased significantly after 3 days of cold stress and then restored to normal level with the recovery of temperature. Histopathological results showed no significant difference in the morphology of mucosal epithelium in the intestine (posterior segment of ileum and large intestine) between acute cold stress group and control group. There were no significant difference in the number of goblet cells, the length of villus and the ratio of villus length to crypt depth in the posterior segment of ileum between acute cold stress group and control group, while the mucosal thickness in the posterior segment of ileum significantly decreased in the acute cold stress group in comparison to control group. Acute cold stress significantly reduced the large intestine number of goblet cells. The structures of the intestinal mucosal mechanical barrier of Chinese soft-shelled turtle under acute cold stress are an intestinal segment dependent pattern. These results suggest that the various segments of the intestine of Chinese soft-shelled turtle have a special response style to acute cold stress.
-
近几十年来, 随着西藏经济社会的快速发展, 西藏鱼类资源也面临着国内其他水域渔业发展所出现的一些共性问题, 如部分流域过度捕捞[1, 2—5]、水利设施建设导致的大坝阻隔、栖息地丧失、生境片段化等[6]。而且由于西藏特殊的地理位置和社会经济特点, 它还面临着更为严峻的生态环境问题和挑战, 如外来物种入侵[7—9]、全球气候变化[10]等, 加之高原生态环境脆弱、生态系统结构简单、生产力低下[11]以及鱼类生长缓慢、早期资源发育周期较长[12]、对生境高度适应和依赖等特点[2—5], 西藏水生生态更容易受到外界的影响。双须叶须鱼处于这种生存环境中, 本身的种群结构是不容乐观的, 对于这种生态系统的扰动, 都将造成鱼类资源的不同程度上的破坏, 恢复过程都将是十分缓慢的, 甚至无法恢复[11]。双须叶须鱼(Ptychobarbus dipogon Regan), 隶属裂腹鱼亚科, 叶须鱼属, 为雅鲁藏布江中游的特有鱼类[1]。2016年双须叶须鱼列入中国脊椎动物红色名录[13]。因此, 必须加快推动双须叶须鱼的养护工作[14]。目前, 有关双须叶须鱼的研究报道, 见于渔业资源调查[15]、营养价值评定[16]、摄食器官与食性[17]、繁殖策略[18]、染色体多样性[19]、性腺发育[20]、年龄和生长及死亡率的关系[21]、年龄鉴定[22]、染色体数目[23]、体长体重关系[24, 25]、线粒体序列[26]等。
鱼类的早期发育研究, 一方面为鱼类的繁殖生物学, 鱼类资源保护与利用提供科学依据和理论基础。另一方面, 鱼类早期发育研究为鱼类的规模化生产提供技术支持。鱼类的受精卵在不同的发育时期对外界环境的反应也不一样, 在生产实践过程中, 应该密切关注外界环境, 规避风险, 减少损失。陈礼强等[27]指出细鳞裂腹鱼的早期发育过程, 原肠期对外界环境变化最为敏感。在原肠期, 由于细胞的分裂迅速和流动量大, 代谢旺盛, 耗氧多, 对环境要求高, 处在这个时期的胚胎如果遇到水温突变等异常情况都可能导致细胞重排时间和空间程序的混乱, 容易造成胚胎的死亡或器官出现畸形[28]。关于裂腹鱼类的早期发育研究, 主要有: 四川裂腹鱼(Schizothorax kozlovi)[29]、青海湖裸鲤(Gymnocypis przewalskii)[30]、松潘裸鲤(Gymnocypris potanini)[31]、齐口裂腹鱼(Schizothorax prenanti)[32]、小裂腹鱼(Schizothorax parvus)[33]、新疆扁吻鱼(Aspiorhynchus laticeps)[34, 35]、塔里木裂腹鱼(Schizothorax (Racoma) biddulphi)[36]、宝兴裸裂尻鱼(Schizopygopsis malacanthus baoxingensis)[37]、昆明裂腹鱼(Schizothorax grahami)[38]、细鳞裂腹鱼(Schizothorax chongi)[27]、黄河裸裂尻鱼(Schizopygopsis pylzovi)[39, 40]、伊犁裂腹鱼(Schizothorax pseudaksaiensis)[41], 异齿裂腹鱼(Schizothorax o’connori)[42]、尖裸鲤(Oxygymnocypris stewartii)[43]、光唇裂腹鱼(Schizothorax lissolabiatus)[44]、厚唇裸重唇鱼(Gymnodiptychus pachycheilus)[45]、祁连山裸鲤(Gymnocypris chilianensis)[46]、短须裂腹鱼(Schizothorax wangchiachii)[47—49]等。未见双须叶须鱼早期发育的研究报道。因此, 开展双须叶须鱼胚胎和仔鱼发育的时序与特点研究就显得尤为重要, 进而为双须叶须鱼的人工繁育技术、资源保护和增殖提供科学依据。
1. 材料与方法
1.1 双须叶须鱼亲鱼获取及孵化
2014年2月, 将采自位于拉萨市墨竹工卡县扎雪乡拉萨河段的双须叶须鱼亲鱼运至西藏农牧学院藏鱼繁育基地。其中, 雌性1尾(体重599.5 g, 体长374 mm), 雄性1尾(体重327 g, 体长310 mm)。通过干法人工授精, 自然产卵, 得到3314粒受精卵。平均分为3组, 将受精卵移入孵化框(55 cm×33 cm×9 cm), 孵化框置于塑料缸(85 cm×65 cm×75 cm) 中, 采用上海海圣控温养殖系统控制水温在10℃左右, 溶解氧8 mg/L以上。用吸管及时剔除死卵, 死鱼苗。
1.2 双须叶须鱼的早期发育观察
使用Nikon SMZ1500体式显微镜(生产商: 日本尼康株式会社)进行观察, ScopePhoto 3.0图像测量软件进行测量、Adobe Photoshop CS6软件处理图片。考虑到各发育期的时间存在个体差异, 将半数以上个体出现新的特征作为发育时期的划分标准, 依据相关文献对双须叶须鱼的胚胎发育过程进行判别[29, 43]。
1.3 数据处理
采用Matlab 2011软件的神经网络聚类算法(Self Organizing Maps, SOM)对已有的裂腹鱼亚科和鲃亚科鱼类的相关文献数据进行收集[29—55], 如卵径、初孵仔鱼长度、吸水卵径、卵周隙、总积温等, 进行聚类分析。
2. 结果
2.1 双须叶须鱼的胚胎发育过程
![]() 图版Ⅰ 双须叶须鱼胚胎发育图1. 受精卵; 2. 卵黄周隙最大; 3. 胚盘隆起; 4. 2细胞期; 5. 4细胞期; 6. 8细胞期; 7. 16细胞期; 8. 32细胞期; 9. 64细胞期; 10. 多细胞期; 11. 桑葚期; 12. 囊胚早期; 13. 囊胚中期; 14. 囊胚晚期; 15. 原肠早期; 16. 原肠中期; 17. 原肠晚期; 18. 神经胚期; 19. 体节出现期; 20. 胚孔封闭期; 21. 眼原基出现期; 22. 眼囊出现期; 23. 听囊期; 24. 耳石出现期; 25. 尾牙出现期; 26. 眼晶体出现期; 27. 肌肉效应期; 28. 心脏原基出现期; 29. 嗅囊期; 30. 心搏期; 31. 胸鳍原基出现期; 32. 肛板期; 33. 血液循环; 34. 尾部鳍褶期; 35. 出膜; 胚盾(16-1); 脊索(18-1); 体节(19-1); 脑泡原基(20-1); 眼原基(21-1); 听囊(23-1); 耳石(24-1); 眼晶体(26-1); 围心腔(28-1); 心脏原基(28-2); 嗅囊(29-1); 胸鳍原基(31-1); 消化道(31-2); 肛板(32-1)图版Ⅰ. The embryonic development of Ptychobarbus dipogon1. Fertilized egg; 2. Perivitelline space maximum; 3. Blastdisc stage; 4. 2 cell stage; 5. 4 cell stage; 6. 8 cell stage; 7. 16 cell stage; 8. 32 cell stage; 9. 64 cell stage; 10. Morula stage; 11. Morula stage; 12. Early blastula stage; 13. Mid-blastula stage; 14. Late gastrula stage; 15. Early gastrula stage; 16. Mid-gastrula stage; 17. Late gastrula stage; 18. Neural embryo formation; 19. Metameres appearance; 20. Blastopore formation; 21. Optic anlage; 22. Eye bud formation; 23. Otic vesicle phase; 24. Otoliths stage; 25. Tail bud stage; 26. Eye lens formation stage; 27. Muscular contraction stage; 28. Heart rudiment stage; 29. Olfactory capsule stage; 30. Heart pulsation stage; 31. Pectoral fin stage; 32. Anal membrane stage; 33. Blood circulation stage; 34. Caudal fin fold stage; 35. Newly hatching stage; Embryonic shield (16-1); Notochord (18-1); Somite (19-1); Brain vesicle primordium (20-1); Eye primordial (21-1); Otic vesicle (23-1); Otoliths (24-1); Eye lens (26-1); Cardiocoelom (28-1); Cardiac primordia (28-2); Olfactory capsule (29-1); Pectoral fin primordial (31-1); Alimentary canal (31-2); Anal membranae (32-1)表 1 双须叶须鱼胚胎发育特征描述Table 1. The embryonic development features of Ptychobarbus dipogon
图版Ⅰ 双须叶须鱼胚胎发育图1. 受精卵; 2. 卵黄周隙最大; 3. 胚盘隆起; 4. 2细胞期; 5. 4细胞期; 6. 8细胞期; 7. 16细胞期; 8. 32细胞期; 9. 64细胞期; 10. 多细胞期; 11. 桑葚期; 12. 囊胚早期; 13. 囊胚中期; 14. 囊胚晚期; 15. 原肠早期; 16. 原肠中期; 17. 原肠晚期; 18. 神经胚期; 19. 体节出现期; 20. 胚孔封闭期; 21. 眼原基出现期; 22. 眼囊出现期; 23. 听囊期; 24. 耳石出现期; 25. 尾牙出现期; 26. 眼晶体出现期; 27. 肌肉效应期; 28. 心脏原基出现期; 29. 嗅囊期; 30. 心搏期; 31. 胸鳍原基出现期; 32. 肛板期; 33. 血液循环; 34. 尾部鳍褶期; 35. 出膜; 胚盾(16-1); 脊索(18-1); 体节(19-1); 脑泡原基(20-1); 眼原基(21-1); 听囊(23-1); 耳石(24-1); 眼晶体(26-1); 围心腔(28-1); 心脏原基(28-2); 嗅囊(29-1); 胸鳍原基(31-1); 消化道(31-2); 肛板(32-1)图版Ⅰ. The embryonic development of Ptychobarbus dipogon1. Fertilized egg; 2. Perivitelline space maximum; 3. Blastdisc stage; 4. 2 cell stage; 5. 4 cell stage; 6. 8 cell stage; 7. 16 cell stage; 8. 32 cell stage; 9. 64 cell stage; 10. Morula stage; 11. Morula stage; 12. Early blastula stage; 13. Mid-blastula stage; 14. Late gastrula stage; 15. Early gastrula stage; 16. Mid-gastrula stage; 17. Late gastrula stage; 18. Neural embryo formation; 19. Metameres appearance; 20. Blastopore formation; 21. Optic anlage; 22. Eye bud formation; 23. Otic vesicle phase; 24. Otoliths stage; 25. Tail bud stage; 26. Eye lens formation stage; 27. Muscular contraction stage; 28. Heart rudiment stage; 29. Olfactory capsule stage; 30. Heart pulsation stage; 31. Pectoral fin stage; 32. Anal membrane stage; 33. Blood circulation stage; 34. Caudal fin fold stage; 35. Newly hatching stage; Embryonic shield (16-1); Notochord (18-1); Somite (19-1); Brain vesicle primordium (20-1); Eye primordial (21-1); Otic vesicle (23-1); Otoliths (24-1); Eye lens (26-1); Cardiocoelom (28-1); Cardiac primordia (28-2); Olfactory capsule (29-1); Pectoral fin primordial (31-1); Alimentary canal (31-2); Anal membranae (32-1)表 1 双须叶须鱼胚胎发育特征描述Table 1. The embryonic development features of Ptychobarbus dipogon阶段Stage 发育时期Developmental stages 时间记录Time records 发育累计时间Cumulative time (h) 特征Character 前期Earlier stage 受精卵 3.24 (23:29) 圆形、淡黄色或橙色具有半透明光泽, 沉性卵, 鱼卵具有微黏性, 受精遇水后失黏, 富含卵黄且分布均匀, 卵径3.7—3.9 mm, 卵黄极性明显, 卵在受精75min后, 膨胀至最大, 卵膜直径5.1—5.3 mm, 卵黄直径3.7—4.3 mm, 胶膜弹性非常强(图版Ⅰ-1)。 胚盘隆起 3.25 (04:57) 4.47 原生质丝向动物极靠拢, 并随着时间推移逐渐变大, 变高, 形成的边缘轮廓并不完整呈星射辐射状, 此刻胚盘的最大高度占卵径1/5, 宽度占卵径2/3 (图版Ⅰ-3)。 卵裂期Cleavage stage 2细胞期 3.25 (07:24) 6.92 胚盘顶部出现一条纵沟, 将动物极分成两个大小相等的分裂球, 卵径4.94 mm, 卵黄直径3.72 mm, 卵膜平均0.5 mm, 分裂球高度平均0.8 mm (图版Ⅰ-4)。 4细胞期 3.25 (10:57) 10.47 出现第二次分裂, 分裂沟与第一次分裂沟垂直, 形成四个大小相等的分裂球(图版Ⅰ-5)。 8细胞期 3.25 (13:15) 13.77 出现第三次分裂, 形成2排8个分裂球, 胚盘逐渐向植物极延伸(图版Ⅰ-6)。 16细胞期 3.25 (17:05) 17.60 出现第四次分裂, 形成16个分裂球, 4排4列整齐排列(图版Ⅰ-7)。 32细胞期 3.25 (20:24) 20.92 完成第五次分裂, 有四个经裂面, 且与第三次分裂面平行, 4排8列整齐的排在同一个平面上(图版Ⅰ-8)。 64细胞期 3.25 (21:54) 22.42 完成第六次分裂, 因细胞分裂速度不一致, 故分裂球的大小, 形态差异明显, 64个细胞排列在一个分裂球面上(图版Ⅰ-9)。 多细胞期 3.25 (23:41) 24.20 出现水平分裂和切线分裂, 分裂球越来越小, 卵裂的速度加快, 形态、体积差异明显, 细胞排列无规律, 细胞界限模糊无法计数细胞的个数(图版Ⅰ-10)。 桑葚期 3.26 (04:26) 28.95 细胞分裂不同步, 细胞数目不断增加, 细胞体积逐渐变小, 多层细胞叠加如同桑葚, 高度占卵黄直径的1/3 (图版Ⅰ-11)。 囊胚期Blastula stage 囊胚早期 3.26 (08:50) 33.52 细胞界限模糊, 分裂球组成的囊胚层隆起, 位于卵黄之上, 高度达卵黄的1/5 (图版Ⅰ-12)。 囊胚中期 3.26 (14:51) 39.52 囊胚层细胞向动物极移动, 变低, 变薄, 胚体高度下降约为卵黄1/4 (图版Ⅰ-13)。 囊胚晚期 3.27 (06:42) 56.52 囊胚细胞向卵黄部位下包, 约占整个细胞的1/3, 动物极较植物色彩暗, 去掉卵膜卵黄破裂, 该期未受精卵开始裂解(图版Ⅰ-14)。 原肠期Gastrula stage 原肠早期 3.28 (19:01) 92.84 胚层细胞约占卵黄的1/3, 背唇呈新月状, 出现光亮明黄的胚环, 胚胎外形呈蘑菇朵状(图版Ⅰ-15)。 原肠中期 3.29 (08:49) 106.64 胚层细胞占卵黄的1/2, 帽状胚层细胞覆盖在卵黄囊上, 在胚胎的背唇处出现箭头似的隆起即是胚盾(16-1, 图版Ⅰ-16)。 原肠晚期 3.29 (16:27) 114.27 胚层下包3/4, 胚盾加厚加长, 胚盾向动物极发展, 胚体的雏形逐渐形成(图版Ⅰ-17)。 神经胚期 Neural embryo formation 神经胚期 3.29 (20:06) 117.92 胚体下包4/5, 能清晰看到胚体的轮廓, 胚盾前端出现神经板, 胚胎外露很小部分卵黄, 细胞内卷内陷形成神经沟, 侧面观察, 胚体背面增厚隆起, 胚体靠近卵黄部形成一条可见的模糊透明圆柱形脊索(18-1, 图版Ⅰ-18)。 体节出现期 3.30 (02:30) 124.32 胚体中部出现2—3对体节(19-1), 胚体占卵黄囊周长1/2—2/3, 神经板头端隆起加大, 尾部也逐渐变大, 头部, 尾部厚于胚体背部, 卵膜可剥, 从顶看胚体不平整(图版Ⅰ-19)。 胚孔封闭期 3.30 (12:18) 134.12 背唇, 腹唇, 侧唇在胚孔处汇合, 将胚孔封闭, 背部颜色较深的条纹为神经沟, 中间凸起明显, 脊索逐渐清晰向尾部延长, 以胚孔方向为正, 左面少于右面, 头部开始膨大隆起的为脑泡原基(20-1, 图版Ⅰ-20)。 器官分化期Qrgan differentiation stage 眼原基出现期 3.30 (20:09) 143.97 脑泡两侧出现椭圆形的隆起即眼原基(21-1), 体节从背部开始向头尾两边增加至6对, 此时胚体绕卵黄4/5 (图版Ⅰ-21)。 眼囊期 3.31 (07:28) 155.29 眼基中央出现像“I”的横凹, 并逐渐扩大变深, 脑泡已经开始分化为前, 中, 后三脑模型, 体节9—10对, 头部出现的铭文皱褶开始加深, 卵黄囊的韧性开始变强, 体节14—15对(图版Ⅰ-22)。 听囊期 3.31 (11:30) 159.29 胚体后脑出现椭圆透明的囊腔, 即为听囊(23-1), 体节17—18对, 此时尾牙模型基本已形成, 呈箭头状脑部铭文状的纹路开始延伸到眼囊后部, 胚体环绕卵黄约5/8, 脊索已成细线条状(图版Ⅰ-23)。 耳石出现期 3.31 (23:36) 171.39 体节19—24对, 听囊增大且清晰, 耳囊内出现两颗透明的小斑即是耳石(24-1, 图版Ⅰ-24)。 尾芽期 4.2 (04:12) 199.99 体节28对, 胚体后端膨大隆起, 卵黄囊椭圆形, 胚体首尾逐步靠近, 尾牙游离于胚体, 靠近尾部的卵黄出现凹陷(图版Ⅰ-25)。 2.2 双须叶须鱼仔稚鱼发育过程
表 2 双须叶须仔稚鱼发育特征描述Table 2. The larvae and juveniles development features of Ptychobarbus dipogon器官出现时期
Qrgan appearance stage时间记录
Time record (d)特征Characters 体色素出现Body pigment
胸鳍上翘Pectoral upward
鳃盖骨出现Operculum
下颌原基出现Underjaw primordial1 全长(12.44±0.15) mm, 肛前长9.39 mm, 肛后长2.78 mm, 心率55—60次/min, 体节60对, 初孵仔鱼颜色透明, 胸鳍上翘, 头部游离卵黄囊, 身体呈S型, 卵黄囊前部呈椭圆型, 后部呈短状棒形。心脏由心房(图版Ⅱ-1d-2)、心室(图版Ⅱ-1d-3)和静脉窦(图版Ⅱ-1d-5)组成, 位于头部之下、卵黄囊前方, 血液从心脏发出沿着背大动脉向前颈动脉流向脑眼等部位, 向后流向尾动脉, 在尾的中部折向, 下入尾静脉在脊索下方入主静脉, 在脑部方向与来自头部的前主静脉汇合, 通过居维氏管进入静脉窦返回心脏, 卵黄囊前端腹部存有血窦(图版Ⅱ-1d-4)。头部靠后1/3处可见食道(图版Ⅱ-1d-7)雏形, 泄殖孔(图版Ⅱ-1d-8)和鳃盖骨(图版Ⅱ-1d-6)均清晰可见, 耳石上方出现环状耳蜗(图版Ⅱ-1d-9), 下颌原基(图版Ⅱ-1d-1)出现。鱼侧卧静息在水底, 扭动频繁, 此时仔鱼为内源营养需要。 鳃弓原基出现Branchial arch primordial 2 全长14.81 mm, 肛前长11.14 mm, 眼睛的直径0.40 mm, 心率62次/min, 鳃部鳃弓原基(图版Ⅱ-2d-1)出现且有血液流过, 脊柱内血液循环明显, 尾部稍显平直。 消化道出现Alimentary canal
肝胰脏原基出现Hepatopancreas primordial3 心率60—65次/min, 消化道(图版Ⅱ-3d-3)明显, 下方出现肝胰脏原基(图版Ⅱ-3d-2), 泄殖孔处凹陷, 下颌(图版Ⅱ-3d-1)形成, 卵黄囊上的色素斑点变深, 能清晰看到卵黄囊上的血管(图版Ⅱ-3d-4), 仔鱼趋于平直, 胸鳍牙增大呈圆扇形, 活动能力进一步加强。 鳃耙出现Gill raker
体表色素细胞带出现Body pigment cell mass4 全长15.48 mm, 肛前长11.39 mm, 肛后长3.91 mm, 心率60—65次/min, 眼晶体向外凸出, 鳃弓有4对, 鳃弓内出现凸起为鳃耙(图版Ⅱ-4d-1), 口凹加深, 头向前伸, 鳃盖伸长, 心脏靠近胸腔, 仔鱼躯体已有明显的细胞色素团(图版Ⅱ-4d-2), 尾鳍的辐射状条纹下叶多于上叶。 口凹、鳃丝形成Stomatodeum, Gill filament 5 口凹(图版Ⅱ-5d-1)形成但未形成口裂, 口不能张合, 下颌开始微微抽动, 鳃丝(图版Ⅱ-5d-2)形成, 围心腔收缩, 仔鱼集群静息于池角或尺边底。 胸鳍褶、背鳍褶、腹鳍褶出现弹性丝Elastic thread of pectoral fin fold, dorsal fin fold, pelvic fin fold 6 下颌下上抽动回缩频率加大, 口裂(图版Ⅱ-6d-1)清晰, 眼前缘的嗅窝加深, 鱼体腹部出现鳍褶且逐渐增大, 尾椎微微上翘, 尾鳍鳍褶加大, 卵黄囊继续收缩形成哑铃或者棒状的结构, 头部、背部零星地出现黑色素, 星芒状, 鳃盖伸长, 下部可盖住鳃丝, 胸鳍褶、背鳍褶、腹鳍褶出现弹性丝。 鼻凹出现Nose concave
星芒状色素团出现Astral form pigment group7 鼻凹(图版Ⅱ-7d-1)出现, 位于眼前方, 鳃丝血流量加大加快, 鳃盖透明可略微张合, 腹部出现星芒状的黑色素(图版Ⅱ-7d-2), 头骨进一步隆起, 胸鳍进一步变大, 脊索与卵黄囊接触处出现星状黑色素细胞(图版Ⅱ-7d-2), 脊椎模型成形, 仔鱼可以上下游动, 鱼苗进入混合营养期。 鳔前原基出现Swim bladder primordial 9 头部色素斑点增多增大, 眼睛可转动, 胸鳍后部卵黄囊中间有一突起, 为鳔前原基(图版Ⅱ-9d-1), 消化道前端变粗且有皱褶, 血液循环延伸至最末脊椎骨, 下颌开闭自如。胸鳍摆动以便平衡, 鳃丝数量增多。 尾鳍鳍条开始出现Tail fin 11 尾鳍中间间质细胞形成了鳍条, 下颌运动伴随着围心腔的斜上方向的收张, 毛细血管呈网状分布在卵黄囊上。 鳔一室出现Swim bladder one room
半规管形成Semicircular canal13 鳔一室(图版Ⅱ-13d-1)出现, 色素细胞呈片状相连布满体表, 伴随着下颌的张合鳃盖开闭, 卵黄囊呈棒状, 胸鳍末端呈圆弧状, 位于胸位, 消化道有褶皱且增粗, 充塞有饵料, 背鳍原基出现, 清晰可见上颌、下颌、上唇、下唇, 耳囊内半规管形成。 背鳍原基出现Dorsal fin primordial 17 背鳍原基(图版Ⅱ-17d-1)出现, 透明无鳍条。胸鳍的鳍条明显, 可见胸鳍支鳍骨。尾鳍圆弧形, 尾鳍鳍条数目增多且有黑色素。肌节清晰, 腹鳍变宽, 腹部有点状色素, 眼膜络网、晶状体、视网膜清晰可见, 肝胰脏增大, 有排泄物排出。 胸鳍鳍条出现Pectoral
fins鳔二室出现Swim bladder two room19 鳔二室出现(图版Ⅱ-19d-1), 鳃盖骨可见清晰横状纹路。 腹部鳍褶变大Pelvic fin fold became bigger 21 卵黄消失, 背鳍透明无鳍条, 仔鱼体色呈褐色, 舌颌骨(图版Ⅱ-21d-1)出现, 围心腔不透明。 臀鳍原基出现Anal fin primordial
脾脏出现Spleen
腹鳍原基出现Pelvic fin primordial28 吻部较尖, 鳔室增大明显, 鳔前室变大, 仍未充气胸鳍透明, 鳔前端腹部有一黑褐色圆饼状组织为脾脏(图版Ⅱ-28d-1), 尾鳍鳍条20条左右, 背鳍较小, 外缘呈锯齿状, 背鳍原基出现3根辐射状纹, 泄殖孔前方出现腹鳍原基(图版Ⅱ-28d-2), 泄殖孔后方出现臀鳍原基(图版Ⅱ-28d-3)。鳃盖上具有一条纹路。 侧线出现Lateral line 29 鳔二室充气明显, 体侧中央可见一条黑色细线, 为侧线(图版Ⅱ-29d-1)。 腹鳍出现Pelvic fin ray 33 腹部中央可见呈辐射状腹鳍(图版Ⅱ-33d-1), 尾部鳍条清晰, 尾鳍边缘呈锯齿状, 背部的肌肉增厚。 鳞片出现Scale 34 两鳔室下方可见闪光质的鳞片(图版Ⅱ-34d-1), 消化道前端膨大, 后端渐小, 鳞片逐渐由腹部下方向上方覆盖, 由鳔向泄殖孔方向覆盖。 38 泄殖孔附近出现两条白色的管道, 并在泄殖孔口汇合, 臀鳍有模糊不清的条纹。 76 尾鳍后缘呈叉型, 鱼体体色近成鱼, 呈黄褐色, 鳞片覆盖体表。 ![]() 图版Ⅱ 双须叶须鱼仔稚鱼发育图1d-1. 下颌原基; 1d-2. 心房; 1d-3. 心室; 1d-4. 血窦; 1d-5. 静脉窦; 1d-6. 鳃盖骨; 1d-7. 食道; 1d-8.泄殖孔; 1d-9. 耳蜗; 2d-1. 鳃弓原基; 3d-1. 下颌; 3d-2. 肝胰脏原基; 3d-3. 消化道; 3d-4. 血管; 4d-1. 鳃耙; 4d-2. 体色素细胞团; 5d-1. 口凹; 5d-2. 鳃丝; 6d-2. 胸鳍褶 6d-3. 背鳍褶; 6d-4. 腹鳍褶; 7d-1. 鼻凹; 7d-2. 星芒状色素团; 9d-1. 鳔前原基; 13d-1. 鳔一室; 17d-1. 背鳍原基; 17-2. 胸鳍鳍条; 19d-1. 鳔二室; 21d-1. 舌颌骨; 28d-1. 脾脏; 28d-2. 腹鳍原基; 28d-3. 臀鳍原基; 29d-1. 侧线; 33d-1. 腹鳍鳍条; 34d-1. 鳞片图版Ⅱ. The larvae and juveniles development features of Ptychobarbus dipogon1d-1. Underjaw primordial; 1d-2. Atrium; 1d-3. Ventricle; 1d-4. Blood sinus; 1d-5. Venous sinus; 1d-6. Operculum; 1d-7. Esophagus; 1d-8. Cloacal orifice; 1d-9. Cochlea; 2d-1. Branchial arch primordial; 3d-1. Underjaw; 3d-2. Hepatopancreas primordial; 3d-3. Alimentary canal; 3d-4. Blood vessel; 4d-1. Gill raker; 4d-2. Body pigment cell mass; 5d-1. Stomatodeum; 5d-2. Gill filament; 6d-2. Pectoral fin fold; 6d-3. Dorsal fin fold; 6d-4. Pelvic fin fold; 7d-1. Nose concave; 7d-2. Astral form pigment group; 9d-1. Swim bladder primordial; 13d-1. Swim bladder one room; 17d-1. Dorsal fin primordial; 17-2. Pectoral fins; 19d-1. Swim bladder two room; 21d-1. Hyomandibular; 28d-1. Spleen; 28d-2. Pelvic fin primordial; 28d-3. Anal fin primordial; 29d-1. Lateral line; 33d-1. Pelvic fin ray; 34d-1. Scale
图版Ⅱ 双须叶须鱼仔稚鱼发育图1d-1. 下颌原基; 1d-2. 心房; 1d-3. 心室; 1d-4. 血窦; 1d-5. 静脉窦; 1d-6. 鳃盖骨; 1d-7. 食道; 1d-8.泄殖孔; 1d-9. 耳蜗; 2d-1. 鳃弓原基; 3d-1. 下颌; 3d-2. 肝胰脏原基; 3d-3. 消化道; 3d-4. 血管; 4d-1. 鳃耙; 4d-2. 体色素细胞团; 5d-1. 口凹; 5d-2. 鳃丝; 6d-2. 胸鳍褶 6d-3. 背鳍褶; 6d-4. 腹鳍褶; 7d-1. 鼻凹; 7d-2. 星芒状色素团; 9d-1. 鳔前原基; 13d-1. 鳔一室; 17d-1. 背鳍原基; 17-2. 胸鳍鳍条; 19d-1. 鳔二室; 21d-1. 舌颌骨; 28d-1. 脾脏; 28d-2. 腹鳍原基; 28d-3. 臀鳍原基; 29d-1. 侧线; 33d-1. 腹鳍鳍条; 34d-1. 鳞片图版Ⅱ. The larvae and juveniles development features of Ptychobarbus dipogon1d-1. Underjaw primordial; 1d-2. Atrium; 1d-3. Ventricle; 1d-4. Blood sinus; 1d-5. Venous sinus; 1d-6. Operculum; 1d-7. Esophagus; 1d-8. Cloacal orifice; 1d-9. Cochlea; 2d-1. Branchial arch primordial; 3d-1. Underjaw; 3d-2. Hepatopancreas primordial; 3d-3. Alimentary canal; 3d-4. Blood vessel; 4d-1. Gill raker; 4d-2. Body pigment cell mass; 5d-1. Stomatodeum; 5d-2. Gill filament; 6d-2. Pectoral fin fold; 6d-3. Dorsal fin fold; 6d-4. Pelvic fin fold; 7d-1. Nose concave; 7d-2. Astral form pigment group; 9d-1. Swim bladder primordial; 13d-1. Swim bladder one room; 17d-1. Dorsal fin primordial; 17-2. Pectoral fins; 19d-1. Swim bladder two room; 21d-1. Hyomandibular; 28d-1. Spleen; 28d-2. Pelvic fin primordial; 28d-3. Anal fin primordial; 29d-1. Lateral line; 33d-1. Pelvic fin ray; 34d-1. Scale3. 讨论
Hora[56]指出, 裂腹鱼亚科起源于鲃亚科, 喜马拉雅山北面的鱼类区系, 可能是在一个较早的时期, 由具有共同的性状的祖先分化而来的。据曹文宣等[57]对骨骼的比较研究, 裂腹鱼类与鲃亚科中一些原始的属, 如四须鲃属、突吻鱼属和鲃属等的颅骨结构非常相似。这进一步证实了, 裂腹鱼类的祖先, 是近似于鲃亚科中原始属的种类。裂腹鱼亚科和鲃亚科鱼类在生态上的区别, 正如它们目前的分布区域的自然条件所显示, 前者是适应寒冷的气候, 后者是适应于温暖的气候(表 3)。但是, 在气候比较寒冷的青藏高原地区在晚第三纪分布有鲃亚科鱼类, 这说明, 裂腹鱼类的出现和发展, 是与青藏高原的急剧隆起, 及随之发生的自然条件的显著改变息息相关的[57]。
表 3 裂腹鱼亚科与鲃亚科鱼类胚胎发育比较Table 3. Comparison of embryonic development between Schizothoracinae fish and Barbinae fishes鱼类Fish 代码Code 卵径ED 初孵仔鱼长度LNH 吸水卵径
EDI卵周隙PA 总积温AT 数据来源
Data source●裂腹鱼亚科Schizothoracinae ●●裂腹鱼属Schizothorax 光唇裂腹鱼Schizothorax lissolabiatus Fish1 2.20 8.50 3.20 1.00 2189.76 文献[44] 齐口裂腹鱼Schizothorax prenanti Fish2 2.95 11.00 4.20 1.25 2211.00 文献[32] 四川裂腹鱼Schizothorax kozlovi Fish3 2.65 8.30 3.70 1.05 2125.50 文献[29] 塔里木裂腹鱼
Schizothorax (Racoma) biddulphiFish4 1.75 7.50 2.75 1.00 1656.20 文献[36] 细鳞裂腹鱼Schizothorax chongi Fish5 2.80 10.00 3.75 1.45 2108.00 文献[27] 伊犁裂腹鱼
Schizothorax pseudaksaiensisFish6 1.75 7.90 3.03 1.28 2510.00 文献[41] 异齿裂腹鱼Schizothorax o’connori Fish9 2.92 8.97 3.72 0.80 2656.80 文献[42] 异齿裂腹鱼Schizothorax o’connori Fish10 2.40 9.84 3.67 1.27 2451.00 文献[50] 拉萨裂腹鱼Schizothorax waltoni Fish13 2.95 10.67 4.03 1.08 2904.00 文献[50] 昆明裂腹鱼Schizothorax grahami Fish14 2.70 11.10 4.01 1.31 2556.00 文献[38] 小裂腹鱼Schizothorax parvus Fish19 1.90 7.50 3.00 1.10 2495.37 文献[33] 短须裂腹鱼Schizothorax wangchiachii Fish20 2.70 11.36 3.68 0.92 2539.98 文献[49] 短须裂腹鱼Schizothorax wangchiachii Fish21 2.36 8.70 3.68 1.32 3565.30 文献[47] 短须裂腹鱼Schizothorax wangchiachii Fish22 3.18 10.88 3.96 0.78 2633.68 文献[48] ●●裸重唇鱼属Gymnodiptychus 厚唇裸重唇鱼
Gymnodiptychus pachycheilus2.50 − 4.41 1.91 1962.00 文献[45] ●●裸裂尻鱼属Schizopygopsis 黄河裸裂尻鱼Schizopygopsis pylzovi Fish7 2.30 10.10 3.20 0.90 2727.50 文献[40] 黄河裸裂尻鱼Schizopygopsis pylzovi Fish8 2.20 11.20 3.20 1.00 3880.80 文献[39] 宝兴裸裂尻鱼
Schizopygopsis malacanthus baoxingensis3.41 8.99 − − 4006.15 文献[37] 拉萨裸裂尻鱼
Schizopygopsis younghusbandiFish12 2.50 10.86 3.54 1.04 3038.50 文献[50] ●●尖裸鲤属Oxygymnocypris 尖裸鲤Oxygymnocypris stewartii Fish11 2.57 10.27 3.22 0.65 2726.40 文献[43] ●●裸鲤属Gymnocypris 祁连山裸鲤Gymnocypris chilianensis Fish15 1.94 8.20 3.24 1.30 2429.15 文献[46] 松潘裸鲤Gymnocypris potanini Fish16 2.80 8.00 4.00 1.15 2520.00 文献[31] 青海湖裸鲤Gymnocypis przewalskii Fish17 2.10 9.05 4.00 1.90 2574.00 文献[30] ●●叶须鱼属Ptychobarbus 双须叶须鱼Ptychobarbus dipogon Fish18 3.80 12.50 5.17 1.37 3360.20 本研究 ●●扁吻鱼属Aspiorhynchus 新疆扁吻鱼Aspiorhynchus laticeps Fish23 1.87 7.50 3.37 1.50 2511.04 文献[34] 新疆扁吻鱼Aspiorhynchus laticeps Fish24 1.60 7.50 2.70 1.10 2000.00 文献[35] ●鲃亚科Barbinae ●●光唇鱼属Acrossocheilus 云南光唇鱼
Acrossocheilus yunnanensisFish25 1.93 7.50 4.05 2.12 2019.00 文献[51] ●●结鱼属Tor 瓣结鱼Tor brevifilis brebifilis Fish26 1.80 6.60 2.60 0.80 1514.80 文献[52] ●●倒刺鲃属Spinibarbus 黑脊倒刺鲃Spinibarbus caldwelli Fish27 2.00 7.40 3.00 1.00 1199.10 文献[53] 注: (1) ED: Egg diameter (mm); LNH: Length of hatched larvae (mm); EDI: Egg diameter after inflation (mm); PA: perivitelline space (mm); AT: accumulated temperature (h·℃); (2)表格中数据是平均值The data in the form are average values 因此, 通过对24种裂腹鱼类和3种鲃亚科鱼类的5种早期发育特征参数的神经网络聚类分析(图 1), 发现鲃亚科鱼类(3种鱼类)与之演化较为接近的裂腹鱼亚科鱼类(裂腹鱼属, 扁吻鱼属)聚为F1、F2两类(不包括裸鲤属鱼类), 此类群较其他裂腹鱼亚科鱼类早期发育具有如下特点: 卵径较小(2 mm左右)、初孵仔鱼长度较短(8 mm左右), 吸水后卵径小(3 mm左右), 卵周隙较大(>1 mm), 总积温较少(2000h·℃左右)。卵径大小对鱼的早期发育和存活具有重要的生物学意义[54], 同一个物种, 小卵死亡率更高, 而大卵具备更高的生存潜力[55]。裂腹鱼类卵径的变化和演化, 是对分布区域自然环境的不断适应。卵径比较小的裂腹鱼类, 其分布海拔较低, 以部分裂腹鱼属鱼类为例, 主要分布在海拔1250—2500 m之内[57], 海拔低, 气候较为温和, 适合卵径较小的裂腹鱼类的发育, 能够保证物种的延续。而卵径比较大的裂腹鱼类, 其分布海拔较高, 以裸裂尻属鱼类和裸鲤属鱼类为例, 主要分布海拔在3750—4750 m内[57], 海拔高, 气候较为恶劣, 大的卵径, 可以提供较多卵黄等营养物质, 从而保证物种的延续。另外一个佐证就是, 卵径最大的一个类别的鱼类(F3), 与其他3个类别存在显著差异(P<0.05), 其初孵仔鱼长度不是最大, 却小于卵径次之的F4的裂腹鱼类(表 4), 可以推断, 为了适应高海拔的恶劣环境, 卵黄一部分作为内源性营养物质进行消化, 另外一部分则用于适应环境的能量消耗。同时, 卵径究竟与早期发育哪些关键参数存在关系? 发现与总积温对数值存在线性相关, 也就是说卵径越大, 总积温越大(图 2), 可以推断, 卵径大的裂腹鱼类, 其自然分布水域的温度较低, 相关数据有待进一步搜集。
![]() 图 1 裂腹鱼亚科与鲃亚科鱼类胚胎发育重要参数SOM聚类a. 根据ward联系方法, 采用欧氏距离进行聚类分析, 分为四类; b. 根据卵径、初孵仔鱼长度、吸水卵径、卵周隙总积温对裂腹鱼亚科与鲃亚科鱼类胚胎发育进行SOM聚类; Fish1—Fish27参见表 3Figure 1. SOM cluster of key parameters of embryonic development about Schizothoracinae fish and Barbinae fishesa. Dendrogram of the hierarchical cluster analysis according to the ward linkage method using Euclidean distance, defining four cluster; b. Classification of embryonic development about Schizothoracinae fish and Barbinae fishes. Based on egg diameter, length of newly hatched larve, egg diameter after inflation, perivitelline space and accumulated temperature; Fish1—Fish27 referred to Tab. 3表 4 基于SOM聚类出四个类别鱼类胚胎发育特征分析Table 4. Characteristics of SOM clusters for four groups fish embryonic development
图 1 裂腹鱼亚科与鲃亚科鱼类胚胎发育重要参数SOM聚类a. 根据ward联系方法, 采用欧氏距离进行聚类分析, 分为四类; b. 根据卵径、初孵仔鱼长度、吸水卵径、卵周隙总积温对裂腹鱼亚科与鲃亚科鱼类胚胎发育进行SOM聚类; Fish1—Fish27参见表 3Figure 1. SOM cluster of key parameters of embryonic development about Schizothoracinae fish and Barbinae fishesa. Dendrogram of the hierarchical cluster analysis according to the ward linkage method using Euclidean distance, defining four cluster; b. Classification of embryonic development about Schizothoracinae fish and Barbinae fishes. Based on egg diameter, length of newly hatched larve, egg diameter after inflation, perivitelline space and accumulated temperature; Fish1—Fish27 referred to Tab. 3表 4 基于SOM聚类出四个类别鱼类胚胎发育特征分析Table 4. Characteristics of SOM clusters for four groups fish embryonic development指标Index F1 F2 F3 F4 卵径Egg diameter (mm) 1.88±0.21b 2.09±0.32b 2.91±0.43a 2.62±0.34a 初孵仔鱼长度Length of hatching after larvae (mm) 7.5±0.6b 8.16±0.58b 10.44±1.39a 10.52±0.82a 吸水卵径Egg diameter after inflation (mm) 2.88±0.23c 3.58±0.38b 4.12±0.5a 3.5±0.3b 卵周隙Perivitelline
space (mm)1±0.11b 1.5±0.38a 1.27±0.13a 0.87±0.14b 总积温Accumulated temperature (h·℃) 1842.54±474.91b 2533.43±501.56a 2587.17±426.83a 2886.24±465.37a SOM聚类 涉及的鱼类 涉及的属 胚胎发育特征 n° F1 Fish1, Fish4, Fish19, Fish24, Fish26, Fish27 裂腹鱼属, 扁吻鱼属, 结鱼属, 倒刺鲃属 卵径小, 初孵仔鱼长度小, 吸水卵径小, 卵周隙适中, 总积温少。 6 F2 Fish3, Fish6, Fish15, Fish17, Fish21, Fish23, Fish25 裂腹鱼属, 裸鲤属, 扁吻鱼属, 光唇鱼属 卵径适中, 初孵仔鱼长度适中, 吸水卵径适中, 卵周隙大, 总积温适中。 7 F3 Fish2, Fish5, Fish10, Fish13, Fish14, Fish16, Fish18 裂腹鱼属, 叶须鱼属, 尖裸鲤属, 裸裂尻鱼属 卵径大, 初孵仔鱼长度适中, 吸水卵径大, 卵周隙适中, 总积温适中。 7 F4 Fish7, Fish8, Fish9, Fish11, Fish12, Fish20, Fish22 裂腹鱼属, 裸裂尻鱼属 卵径适中, 初孵仔鱼长度大, 吸水卵径适中, 卵周隙小, 总积温多。 7 注: n°表示样本数
Note: n°: Number of samples![]() 图 2 裂腹鱼亚科鱼类与鲃亚科鱼类卵径和总积温时间关系Figure 2. The relationship between egg diameter and accumulated temperature about Schizothoracinae fish and Barbinae fishes
图 2 裂腹鱼亚科鱼类与鲃亚科鱼类卵径和总积温时间关系Figure 2. The relationship between egg diameter and accumulated temperature about Schizothoracinae fish and Barbinae fishes在一定温度范围内, 水温升高可以加快胚胎的发育速度; 相反, 水温降低会减缓胚胎发育速度, 超过温度范围可能引起胚胎发育停滞、畸形或死亡[35, 58, 59]。通常在一定的温度范围内, 温度越高, 发育越快, 但并非总是呈线性增加[43]。因此, 在不影响胚胎正常发育的前提下, 提高孵化的温度, 这对于提高生产效率、降低管理成本和风险、减少疾病感染的机会有着积极的意义[32]。新疆扁吻鱼在适宜温度范围下, 高温(18—21℃)较常温(13—19℃)不仅胚胎发育快(发育速度4倍有余), 而且孵化率高[35]。祁连山裸鲤孵化率随水温升高呈现出先升高后降低的趋势, 畸形率随水温的升高呈现出先降低后升高的趋势[46]。松潘裸鲤的孵化和幼鱼饲养水温不宜超过23℃[31], 而齐口裂腹鱼的孵化水温可高达24℃而不出现异常和畸形[32]。细鳞裂腹鱼在水10.4—22.8℃的条件下能够正常发育[27]。伊犁裂腹鱼自然产卵孵化水温在 18—22℃ 左右, 平均为20℃[41]。
双须叶须鱼卵径在已报道裂腹鱼类卵径最大, 其分布水域海拔从2900 m (林芝县)到4700 m (萨嘎县)均有分布, 我们的调查数据显示, 在水温高于15℃的水域, 就没有该鱼类的分布, 这也是我们控制繁殖水温在10℃的原因所在。
Lake[60]和Matsuura[61]认为在一些鱼类受精卵吸水膨胀后有较大的卵周隙, 可以更好地抗击外界环境的冲击, 提高受精卵的成活率。而卵周隙大小, 直接决定卵的性质, 沉性卵或者漂浮性卵。我们统计发现, 虽然第二类别和第三类别的卵周隙较第一类别和第四类别大, 存在显著差异(P<0.05), 但是裂腹鱼亚科鱼类和鲃亚科鱼类的卵周隙未超过2.0 mm, 而四大家鱼卵周隙均超过了2.0 mm。这也从另外一个角度说明, 裂腹鱼类属沉性卵, 而四大家鱼则属于漂浮性卵。裂腹鱼亚科鱼类区别于鲃亚科的主要形态差异在于肛门和臀鳍基部的两侧各具有一列变形的大鳞片, 称为臀鳞, 其产生与裂腹鱼类适应流水环境繁殖习性有关[57]。裂腹鱼类产卵前, 雄鱼通过尾部的摆动, 以臀鳍和尾鳍在河底掘出小坑, 产出的沉性卵粒便聚集在小坑内进行胚胎发育, 从而避免了被水流冲到河流下游的不适宜环境中去[57]。除裸鲤属鱼类外, 大部分裂腹鱼类产卵集中在3—5月份, 如实验对象双须叶须鱼产卵集中在3月中下旬, 其水域在这个时期恰逢枯水期, 水体较为温和, 相对于底质平坦的浅水滩而言, 沉积在小坑或者鹅卵石缝隙里的裂腹鱼类受精卵在较为稳定水体中发育, 减少了鱼卵被水冲涮和减弱了光线的照射强度。而四大家鱼繁殖期正值涨水季节, 受水流的冲击, 卵周隙大对于卵的保护起着至关重要的作用。
-
![]()
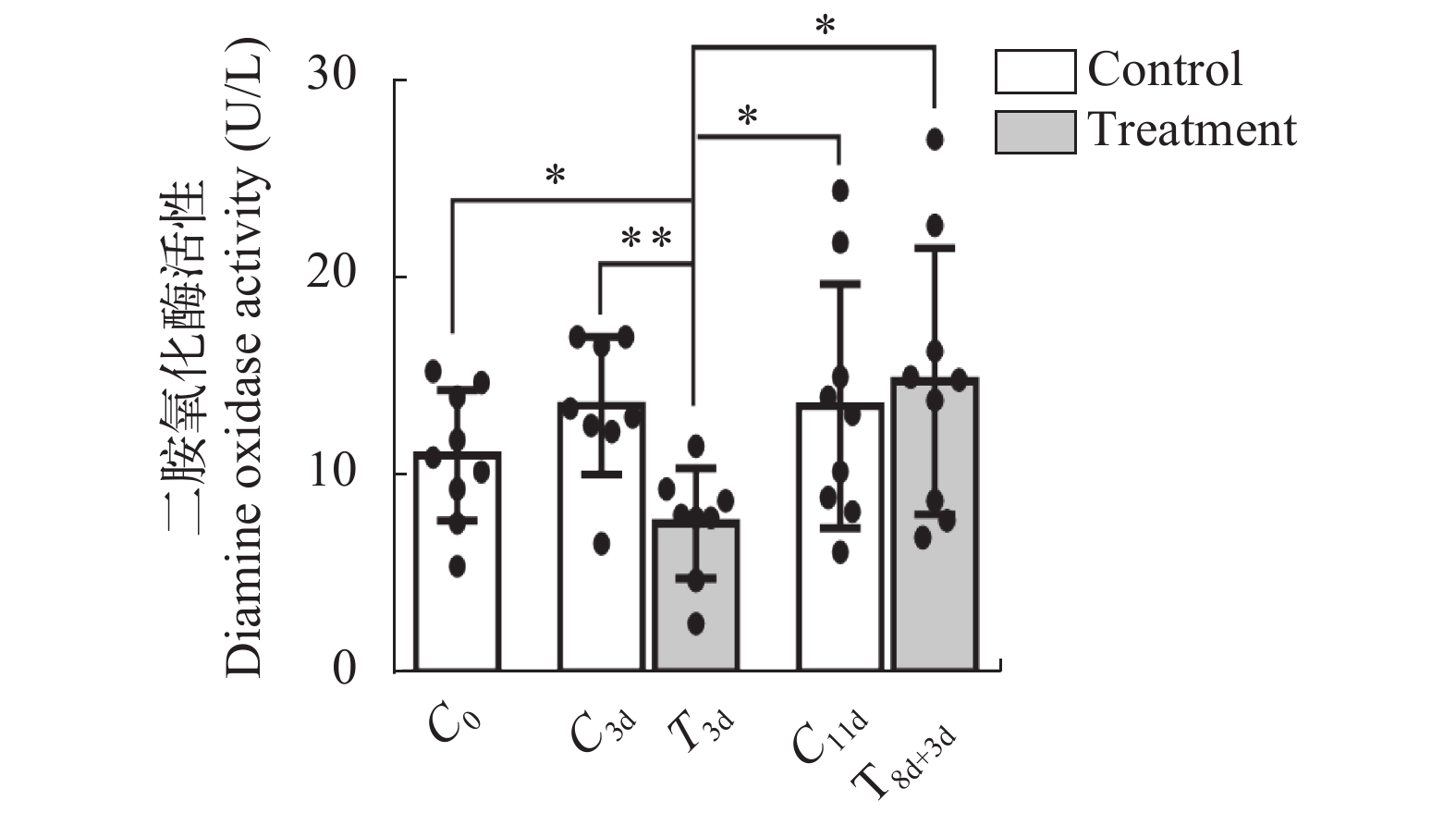
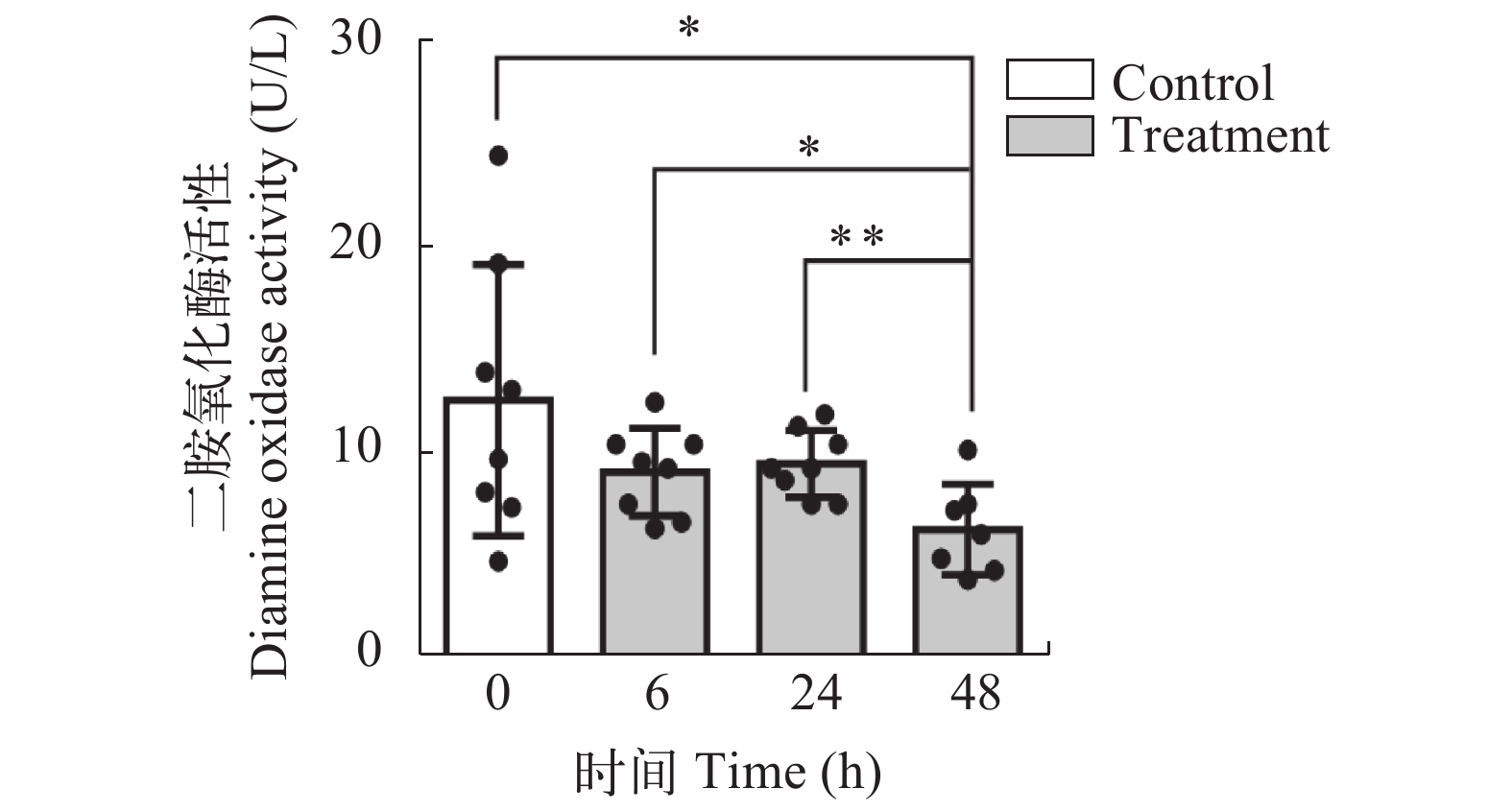
图 1 第一次急性冷胁迫后中华鳖血清DAO活性
*表示48h组与其他各组之间血清DAO活性的显著性分析, *P<0.05, **P<0.01
Figure 1. The DAO activity of Chinese soft-shelled turtle during the first acute cold stress experiment
* Indicates a significant difference between the 48h group and the other groups, * P<0.05, ** P<0.01
![]()
图 2 急性冷胁迫及复温实验中各组中华鳖血清DAO活性
C0. 急性冷胁迫前对照组; C3d. 急性冷胁迫3d对照组; T3d. 急性冷胁迫3d组; C11d. 急性冷胁迫11d对照组; T8d+3d. 急性冷胁迫8d后迅速升温至25℃并维持3d组; *表示T3d组与其他各组之间血清DAO活性的显著性分析, *P<0.05, **P<0.01; 下同
Figure 2. The DAO activity of Chinese soft-shelled turtle in acute cold stress and rewarming experiments
C0. control group before acute cold stress; C3d. 3 days of control group; T3d. 3 days of acute cold stress group; C11d. 11 days of control group; T8d+3d. 8 days of acute cold stress at 15℃ and maintained at 25℃ for 3 days; * Indicates a significant difference between T3d group and the other groups, * P<0.05, ** P<0.01; the same applies below
![]()
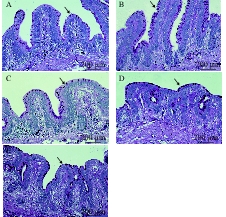
图 3 急性冷胁迫对中华鳖回肠后段黏膜上皮组织形态影响的显微照片(PAS染色×400)
A. 冷胁迫前对照组; B. 冷胁迫3d对照组; C. 冷胁迫3d组; D. 冷胁迫11d对照组; E. 冷胁迫8d后迅速升温至25℃并维持3d组; 箭头所指为杯状细胞
Figure 3. Micrograph of acute cold stress on the mucosal epithelial tissue morphology of the posterior ileum of Chinese soft-shelled turtle (PAS staining×400)
A. control group before acute cold stress; B. 3 days of control group; C. 3 days of acute cold stress group; D. 11 days of control group; E. 8 days of acute cold stress at 15℃ and maintained at 25℃ for 3 days. The arrows refer to goblet cells
![]()
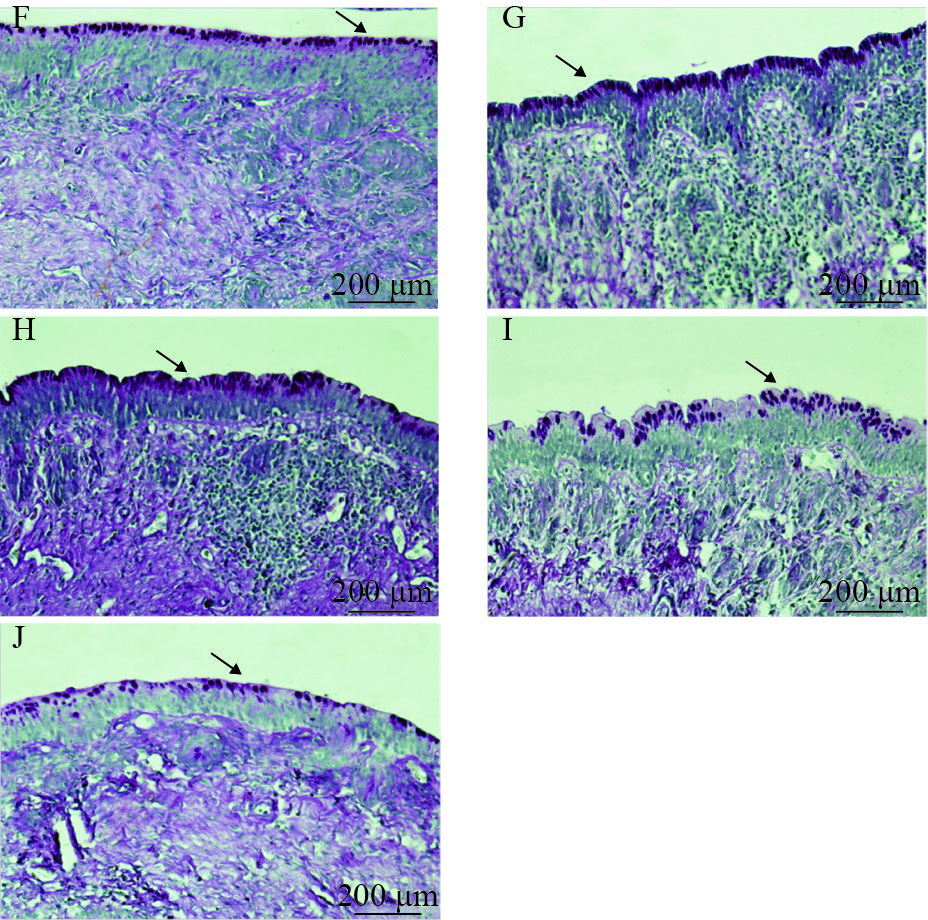
图 4 急性冷胁迫对中华鳖大肠黏膜上皮组织形态影响的显微照片(PAS染色×400)
F. 冷胁迫前对照组; G. 冷胁迫3d对照组; H. 冷胁迫3d组; I. 冷胁迫11d对照组; J. 冷胁迫8d后迅速升温至25℃并维持3d组; 箭头所指为杯状细胞
Figure 4. Micrograph of acute cold stress on the mucosal epithelial tissue morphology of the large intestine of Chinese soft-shelled turtle (PAS staining×400)
F. control group before cold stress; G. control group at day 3 after acute cold stress; H. cold stress group at day 3 day; I. control group at day 11 after acute cold stress; J. cold stress group at day 8 and maintained at 25℃ for 3 days. The arrows refer to goblet cells
![]()
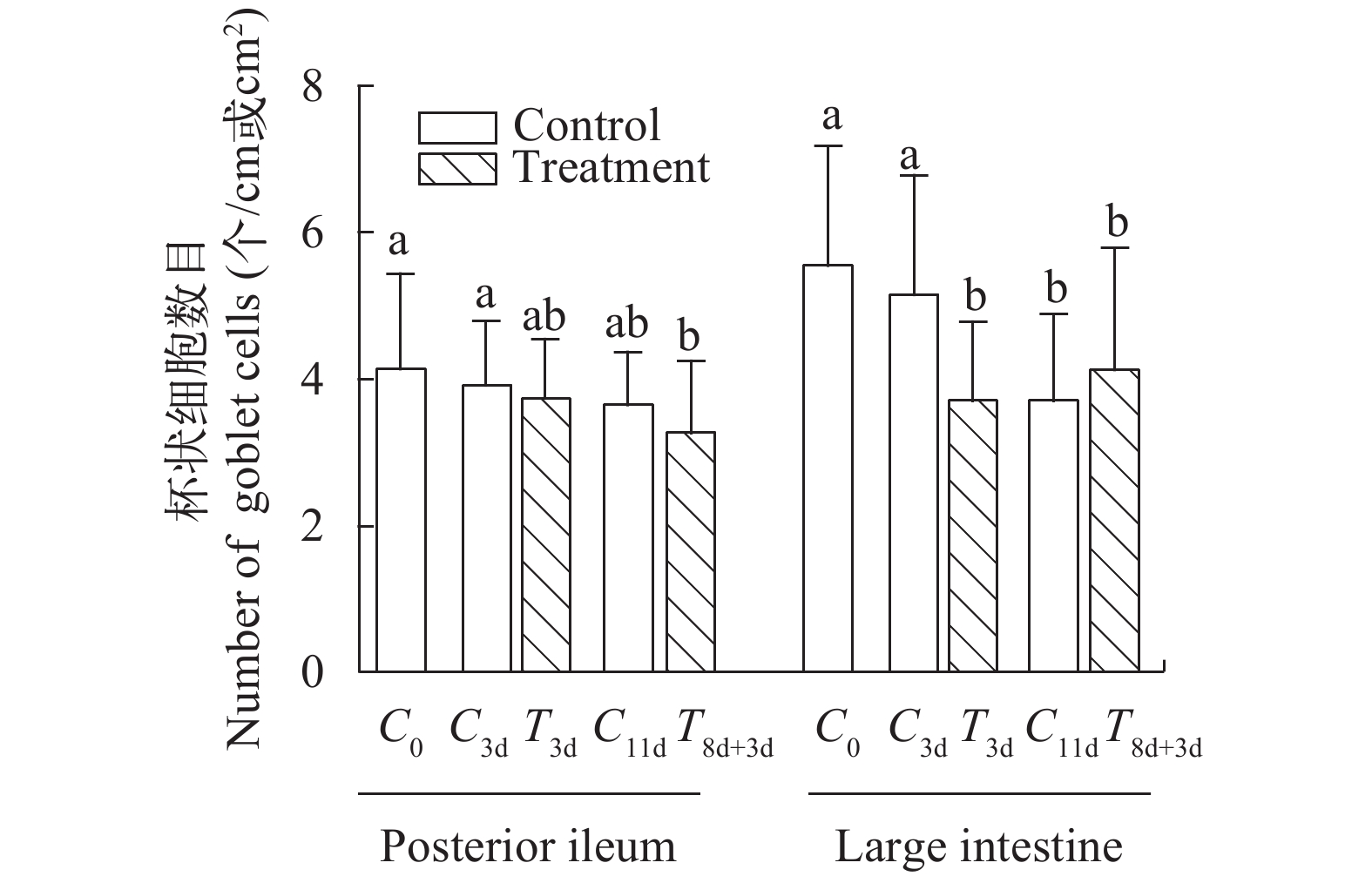
图 5 急性冷胁迫及复温实验中回肠后段/大肠杯状细胞数目变化
a与ab/b与ab表示组间无显著性差异(P>0.05), a与b表示组间差异显著(P<0.05)
Figure 5. The number of goblet cells in the posterior segment of ileum/the large intestine in acute cold stress and rewarming experiments
a and ab/b and ab showed no significant difference between the groups (P> 0.05), a and b showed significant differences between the groups (P<0.05)
![]()
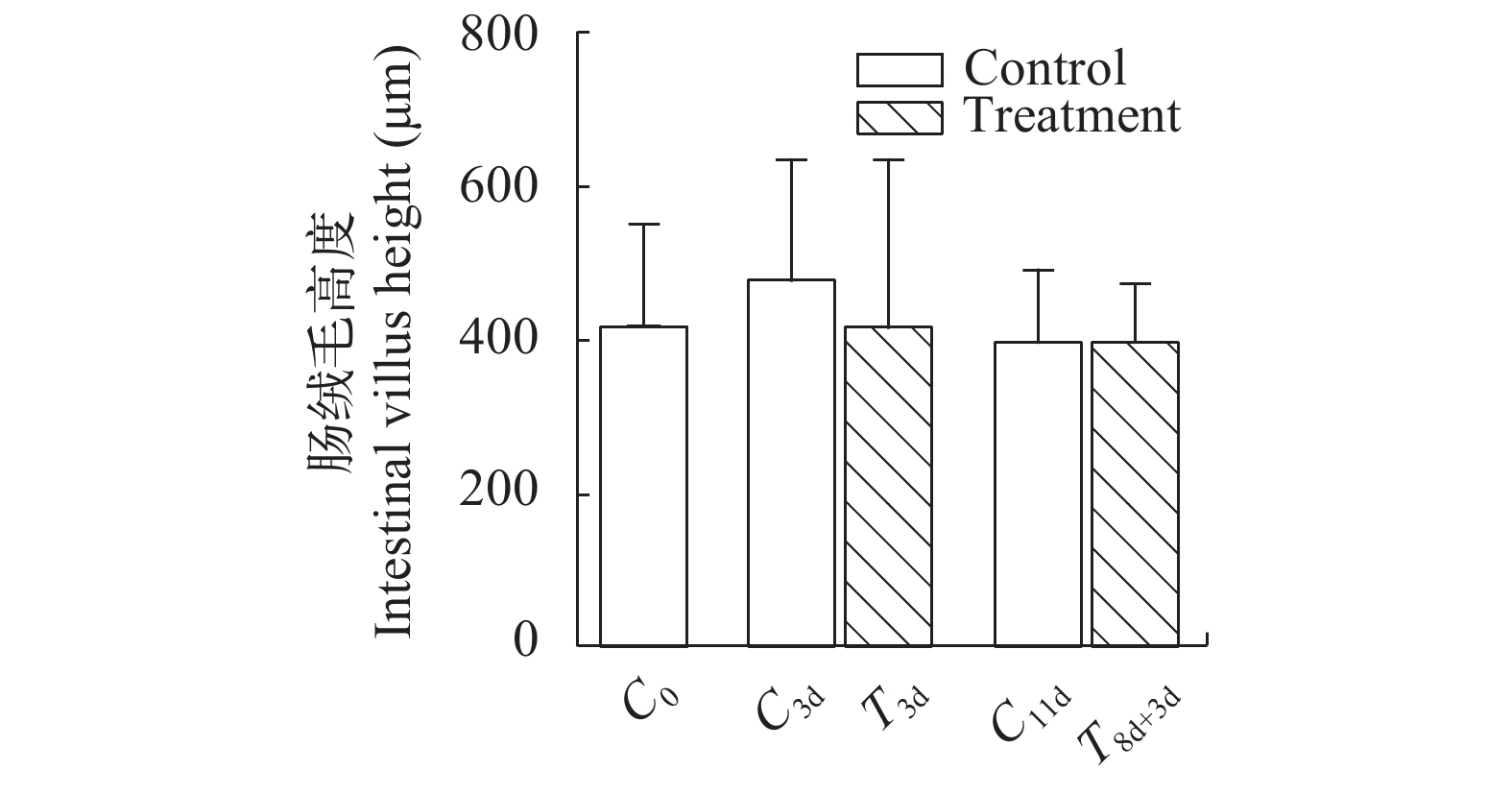
图 6 急性冷胁迫及复温实验中回肠后段肠绒毛高度变化
Figure 6. Intestinal villus height in the posterior segment of ileum during acute cold stress and rewarming experiments
![]()
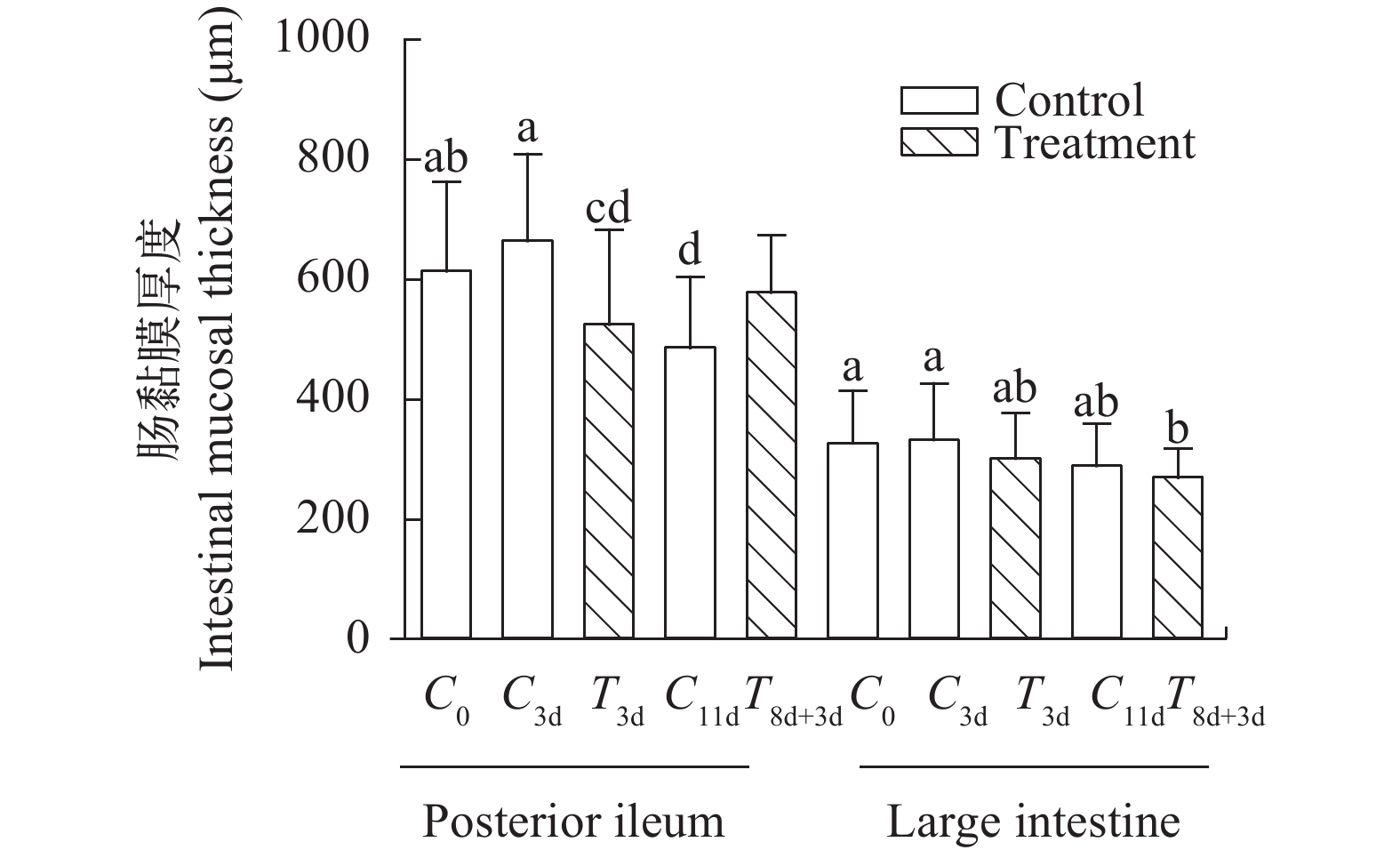
图 7 急性冷胁迫及复温实验中回肠后段/大肠黏膜厚度变化
Figure 7. Changes of colorectal mucosa in the posterior segment of ileum/large intestine during acute cold stress and rewarming experiment
![]()
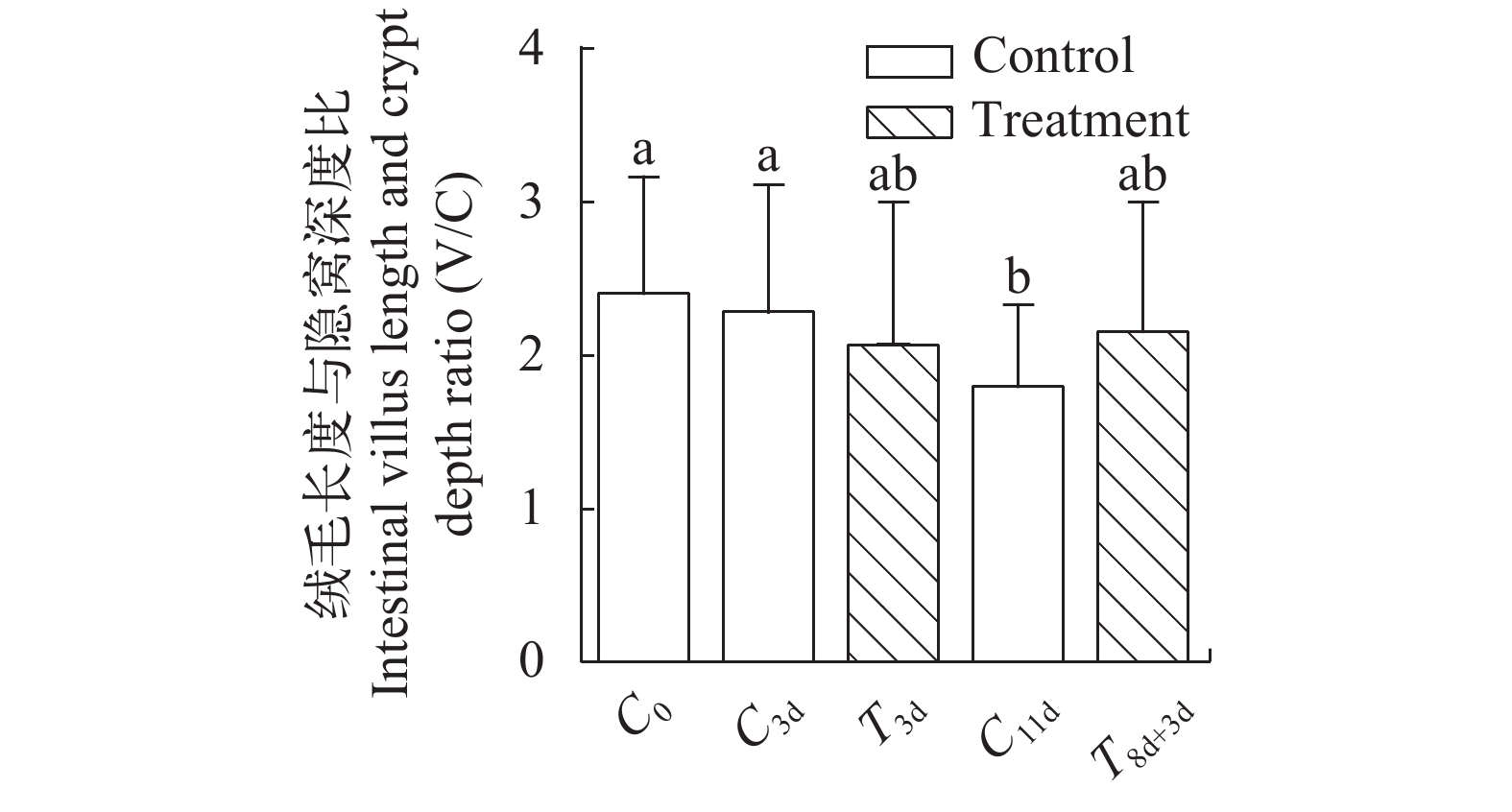
图 8 急性冷胁迫及复温实验中回肠后段绒毛长度/隐窝深度(V/C)变化
Figure 8. Ratios of villus length/crypt depth (V/C) in posterior segment of ileum during acute cold stress and rewarming experiment
表 1 急性冷胁迫对中华鳖回肠后段/大肠杯状细胞数的影响
Table 1 Effects of acute cold stress on the number of goblet cells in the posterior segment of ileum/the large intestine in Pelodiscus sinensis
组别
Group回肠后段杯状
细胞数(个/cm2)
The number of goblet
cells in the posterior
segment of ileum大肠杯状细
胞数(个/cm)
The number of goblet
cells in the large
intestineC0 4.1±1.3a 5.5±1.6a C3d 3.9±0.8a 5.1±1.6a T3d 3.7±0.8ab 3.7±1.0b C11d 3.6±0.7ab 3.7±1.1b T8d+11d 3.3±1.0b 4.1±1.6b F值 3.038 8.602  下载: 导出CSV
下载: 导出CSV
表 2 急性冷胁迫及复温对中华鳖回肠后段绒毛长度、肠壁厚度、大肠肠壁厚度及绒毛长度隐窝深度的影响
Table 2 Effects of acute cold stress and rewarming on villus length, intestinal wall thickness in the posterior segment of ileum, intestinal wall thickness in the large intestine and the villi length/crypt depth of Chinese soft-shelled turtle
组别
Group回肠后段绒毛长度
The villus length in
the posterior segment
of ileum (μm)回肠后段黏膜厚度
The intestinal mucosa
thickness in the posterior
segment of ileum (μm)大肠黏膜厚度
The intestinal mucosa
thickness in the large
intestine (μm)绒毛长度/隐窝深度
The villi length/
crypt depth (V/C)C0 416.8±132.0 617.1±142.1ab 331.0±79.3a 2.4±0.8a C3d 478.7±153.7 667.9±141.9a 330.2±96.4a 2.3±0.8a T3d 417.2±211.4 524.2±152.7cd 299.2±72.1ab 2.1±0.9ab C11d 397.6±92.2 490.6±110.9d 293.7±59.3ab 1.8±0.5b T8d+3d 402.4±68.7 574.5±93.4bc 272.3±41.1b 2.1±0.8ab F值 1.287 7.126 2.934 2.150
下载: 导出CSV
-
[1] 黄丽英, 何中央, 丁诗华, 等. 中华鳖种质资源的研究现状及保护利用对策. 宁波大学学报(理工版), 2005, 18(2): 183—186 doi: 10.3969/j.issn.1001-5132.2005.02.011 Huang L Y, He Z Y, Ding S H, et al. Current status and countermeasures for protecting and utilizing on soft-shelled turtle (Pelodiscus sinensis) germplasm resources in China [J]. Journal of Ningbo University(
Natural Science and Engineering Edition ) , 2005, 18(2): 183—186 doi: 10.3969/j.issn.1001-5132.2005.02.011 [2] Somfai-Relle S, Schauss A G, Financsek I, et al. Acute and subchronic toxicity studies of cryogenically-frozen, cryomilled, Pelodiscus sinensis (Japanese soft-shelled turtle-suppon) powder administered to the rat [J]. Food & Chemical Toxicology, 2005, 43(4): 575—580
[3] 李士泽, 袁学军, 杨玉英, 等. 浸水冷应激对雏鸡某些酶活性及消化道黏膜充血的影响. 应用与环境生物学报, 2001, 7(3): 267—270 doi: 10.3321/j.issn:1006-687X.2001.03.016 Li S Z, Yuan X J, Yang Y Y, et al. Effect of cold stress by immersing in water on some enzyme activities and digestive tract mucosal hyperemia in chicken [J]. Applied and Environmental Biology, 2001, 7(3): 267—270 doi: 10.3321/j.issn:1006-687X.2001.03.016
[4] Kaushik S, Kaur J. Effect of chronic cold stress on intestinal epithelial cell proliferation and inflammation in rats [J]. Stress-the International Journal on the Biology of Stress, 2005, 8(3): 191—197 doi: 10.1080/10253890500245953
[5] Walser E M, Nealon W H, Marroquin S, et al. Sterile fluid collections in acute pancreatitis: catheter drainage versus simple aspiration [J]. Cardiovascular & Interventional Radiology, 2006, 29(1): 102—107
[6] 牛海静, 汪邦茂. 肠黏膜屏障与功能. 解剖与临床, 2007, 12(2): 138—140 doi: 10.3969/j.issn.1671-7163.2007.02.024 Niu H J, Wang B M. Intestinal mucosal barrier and its function [J]. Anatomy and Clinics, 2007, 12(2): 138—140 doi: 10.3969/j.issn.1671-7163.2007.02.024
[7] 成令忠. 组织学(第2版). 北京: 人民卫生出版社. 1994, 1102—1125 Cheng L Z. Histology (second edition) [M]. Beijing: Peoples Medical Press. 1994, 1102—1125
[8] Reynolds J V. Gut barrier function in the surgical patients [J]. British Journal of Surgery, 1996, 83(12): 1668—1671 doi: 10.1002/(ISSN)1365-2168
[9] Fukudome I, Kobayashi M, Dabanaka K, et al. Diamine oxidase as a maker of intestinal mucosal injury and the effect of soluble dietary fiber on gastrointestinal tract toxicity after intravenous 5-fluourouracil treatment in rats [J]. Medical Molecular Morphology, 2014, 47(2): 100—107 doi: 10.1007/s00795-013-0055-7
[10] Karabulut K U, Narci H, Gul M, et al. Diamine oxidase in diagnosis of acute mesenteric ischemia [J]. American Journal of Emergency Medicine, 2013, 31(2): 309—312 doi: 10.1016/j.ajem.2012.07.029
[11] 吴文明, 张方信, 张盼, 等. 高原缺氧条件对大鼠肠黏膜组织及缺氧诱导因子一1α、诱导型-一氧化氮合酶表达的影响. 解放军医学杂志, 2010, 35(5): 592—594 Wu W M, Zhang F X, Zhang P, et al. Influence of plateau hypoxia on the tissue injury and expression of HIF-1α and iNOS in intestinal mucosa of rats [J]. Medical Journal of Chinese People’s Liberation Army, 2010, 35(5): 592—594
[12] Bonous G, Echave V, Vobecky S J, et al. Acute necrosis of the intestinal muesoa and high serum levels of diamine oxidase [J]. Digestive Diseases and Sciences, 1984, 29(9): 872—874 doi: 10.1007/BF01318436
[13] Luk G D, Bayless T M, Baylin S B. Diamine oxidase (DAO) a circulating marker for rat mucosal maturation and integrity [J]. Journal of Clinical Investigation, 1980, 66(1): 66—70 doi: 10.1172/JCI109836
[14] Nakao M, Ogura Y, Satake S, et al. Usefulness of soluble dietary fiber for the treatment of diarrhea during enteral nutrition in elderly patients [J]. Nutrition, 2002, 18(1): 35—39 doi: 10.1016/S0899-9007(01)00715-8
[15] Fu X B, Xing F, Yang Y H, et al. Activation of phosphorylating-p38 mitogen-activated protein kinase and its relationship with localization of intestinal stem cells in rats after ischemiareperfusion injury [J]. World Journal of Gastroenterology, 2003, 9(9): 2036—2039
[16] 黎君友, 盛志勇, 吕艺, 等. 严重创伤后肠屏障功能损伤及谷氨酰胺的保护. 世界华人消化杂志, 2000, 8(10): 1093—1096 doi: 10.3969/j.issn.1009-3079.2000.10.004 Li J Y, Sheng Z Y, Lü Y, et al. Severe trauma induced intestinal barrier function injury and protection [J]. World Chinese Journal of Digestology, 2000, 8(10): 1093—1096 doi: 10.3969/j.issn.1009-3079.2000.10.004
[17] Mantis N J, Cheung M C, Chintalcharuvu K R, et al. Selective adherence of IgA to murine Peyer's patch M cells: evidence for a novel IgA receptor [J]. Journal of Immunology, 2002, 169(4): 1844—1851 doi: 10.4049/jimmunol.169.4.1844
[18] 王自力, 于同泉, 朱晓宇, 等. 中药复方对热应激下猪肠道组织IL-2 IL-10和黏液IgA含量影响. 中国兽药杂志, 2007, 43(9): 83—85 Wang Z L, Yu T Q, Zhu X Y, et al. Effects of complex prescriptions of Chinese herbs on the concentration of IL-2, IL-10 and IgA in porcine intestines after heat stress [J]. Chinese Journal of Veterinary Medicine, 2007, 43(9): 83—85
[19] 白晓东. 肠道黏液层研究进展. 国外医学: 创伤与外科基本问题分册, 1997, (4): 200—202 Bai X D. Intestinal mucous layer research progress [J]. Foreign Medicine: Volumes of Trauma and Surgical Problems, 1997, (4): 200—202
[20] Olsen R E, Sundell K, Mayhew T M, et al. Acute stress alters intestinal function of rainbow trout, Oncorhynchus mykiss [J]. Aquaculture, 2005, 250(1-2): 480—495 doi: 10.1016/j.aquaculture.2005.03.014
[21] Fu J, Liu C P, Zhang Z W, et al. Influence of inflammatory pathway markers on oxidative stress induced by cold stress in intestine of quails [J]. Research in Veterinary Science, 2013, 95(2): 495—501 doi: 10.1016/j.rvsc.2013.05.006
[22] Glaser R, Kiecoltglaser J K. Stress-induced immune dysfunction: implications for health [J]. Nature Reviews Immunology, 2005, 5(3): 243—251 doi: 10.1038/nri1571
[23] Zhu G F, Chancellor-Freeland C, Berman A S, et al. Endogenous substance P mediates cold water stress-induced increase in interleukin-6 secretion from peritoneal macrophages [J]. Journal of Neuroscience the Official Journal of Society for Neuroscience, 1996, 16(11): 3745—3752 doi: 10.1523/JNEUROSCI.16-11-03745.1996
[24] 张雯. 冷应激对雏鸡肠黏膜屏障功能的影响. 东北农业大学. 2010 Zhang W. Effects of cold stress on chicken’s barrier function of intestinal mucosa [D]. The Northeast Agricultural University. 2010
[25] 韩正康. 家畜营养生理学. 北京: 农业出版社. 1991, 16—17 Han Z K. Livestock Nutrition Physiology [M]. Beijing: Agriculture Press. 1991, 16—17
-
期刊类型引用(5)
1. 郭祉宾,杨德国,陈亮,颜文斌,朱挺兵. 雅鲁藏布江中游仔稚鱼种类组成与分布特征. 淡水渔业. 2024(02): 13-22 .  百度学术
百度学术
2. 李文康,骆小年,段友健,李姣,吴晨. 鸭绿沙塘鳢仔稚鱼发育观察及饥饿不可逆点的确定. 大连海洋大学学报. 2023(01): 32-42 . 百度学术
3. 石叶忠,朱双全,潘月,申屠琰,冯彬彬,程鑫,张克鑫,朱卫东,夏荣兴,张玉明,竺俊全. 宽鳍鱲胚胎发育研究. 浙江海洋大学学报(自然科学版). 2022(04): 308-314 . 百度学术
4. 韩军军,贺疆滔,陈朋,胡江伟,祁峰,封永辉,蔡林钢,时春明,张人铭. 扁吻鱼胚胎发育及仔稚鱼形态与生长观察研究. 南方水产科学. 2021(01): 59-66 . 百度学术
5. 陈军平,沈方方,武慧慧,张佳鑫,于若梦,付永杰,谢国强,赵道全. 我国鱼类胚胎发育研究进展. 江苏农业科学. 2021(17): 45-52 . 百度学术
其他类型引用(2)




计量
- 文章访问数: 1957
- HTML全文浏览量: 633
- PDF下载量: 52
- 被引次数: 7