
EFFECTS OF HIGH ENERGY DIETS ON SERUM BIOCHEMICAL INDICES OF GRASS CARP (CTENOPHARYNGODON IDELLA)
-
摘要: 为研究草鱼(Ctenopharyngodon idella) 在摄食高能饲料后血清生化指标的动态变化, 选择均重为(55.0±2.5) g 的健康草鱼320 尾, 随机分成4 组, 每组4个重复, 分别以3组高能纯化饲料(分别以额外的蛋白质、脂肪、糊精供能, 高能饲料组较对照组总能量提高11%)及对照组饲料喂养草鱼11周后, 将鱼体禁食48h后采样(对照), 再饱食投喂, 在摄食后的2h、8h和24h取样, 测定实验鱼8种血清生化指标, 并对采样时间点(T)和不同饲料(D)进行双因素方差分析。为确保实验结果的准确性, 需要保证不同处理组的采样时间同步性, 即均在餐后同一时间点采样。结果显示: 除血清总胆固醇(TCHO)含量外, 血清碱性磷酸酶(ALP)、谷草转氨酶(AST)、谷丙转氨酶(ALT)活性, 以及血清甘油三酯(TAG)、葡萄糖(GLU)、低密度脂蛋白胆固醇(LDLC)、高密度脂蛋白胆固醇(HDLC)含量均受到采样时间点(T)即餐后时间的显著影响, 饲料(D)对上述8个指标亦均有显著影响, 且两者存在交互作用。草鱼血清GLU含量和血清AST活性均在餐后2h达到峰值, 血清ALT和ALP活性以及LDLC含量均在餐后24h达到峰值, 血清TAG含量在餐后8h达到峰值, 而血清TCHO和HDLC含量则无明显峰值。在餐后24h, 3个高能组的ALT活性和GLU含量较对照组显著升高, 血清AST和ALP活性以及TAG、TCHO和HDLC含量较对照组无显著差异。上述结果表明, 草鱼血清生化指标受到餐后采样时间点和饲料的显著影响。Abstract: To investigate effects of high energy diets on serum biochemical indices of grass carp (Ctenopharyngodon idella), grass carp [N= 320, average initial body weight=(55.0±2.5) g] were randomly assigned into 4 groups with 4 replicates for each group (N=20 fish/replicate) to fed three times daily to satiation with the control diet (CON) and three high energy diets for 11 weeks. The 3 high energy diets were 11% increased protein, 11% increased fat or 11% increased Carbohydrate compared to the control diet, respectively. The fish were fasted for 48h and then refed a single meal for 0, 2, 8, or 24 hours before collection for analyzing eight serum biochemical indices. Statistical analysis using two-way ANOVA showed that seven serum biochemical indices except total cholesterol (TCHO) were significantly affected by refeeding time, and all 8 biochemical indices were significantly affected by the different diets. Peak values of glucose (GLU) contents and aspartate transaminase (AST) activities presented at 2h after refeeding, and peak values of alanine transaminase (ALT), alkaline phosphatase (ALP) activities and low density lipoprotein-cholesterol (LDLC) contents appeared at 24h after refeeding. The peak value of triacylglycerol (TAG) content occurred at 8h after refeeding, but there was no obvious peak value for serum TCHO or high density lipoprotein-cholesterol (HDLC) contents. At 24h after refeeding, compared with the CON diet, the higher energy diets had higher GLU contents and ALT activities but with similar activities of AST and ALP, and the contents of TAG, TCHO and HDLC. Our results demonstrated that both refeeding time and high energy diets regulated the serum biochemical indices, indicating that it is necessary to make sampling time synchronize in different groups for the accuracy of serum biochemical indices.
-
Keywords:
- Serum biochemical indices /
- Sample time /
- High energy diets /
- Ctenopharyngodon idella
-
鱼类早期资源指精卵结合至早期稚鱼阶段, 是鱼类个体生活史的开端[1]。通过对鱼类早期资源的研究, 可以了解鱼类的繁殖活动、产卵场位置分布和繁殖时间等早期生活史特征[1]。同时, 早期资源也是影响种群数量变化的主要原因之一[2]。然而, 捕捞、水利工程建设和江湖阻隔等人类活动造成了鱼类早期资源的急剧减少[3, 4]。例如, Sanches等[5]研究结果显示, 水利工程导致巴拉那河的鱼苗密度有显著变化。过度捕捞造成地中海沿海区域鱼类的产卵活动明显减少[6]。
长江中游是多种鱼类的产卵场和栖息地。刘明典等[7]2014—2015年在宜昌江段调查到27种鱼类卵苗。李世健等[8]在监利江段调查到51种鱼类的鱼卵和仔稚鱼。黎明政等[9]在武穴江段调查到27种鱼类的卵苗。同时, 长江中游也是重要经济鱼类四大家鱼的主要产卵场之一。20世纪60年代, 长江中游四大家鱼的产卵场有25处, 产卵量占长江总量的四分之三。其中, 宜昌江段是四大家鱼产卵规模最大的产卵场, 平均鱼卵径流量可达119.48×108粒, 占长江干流总量的9.7%[10]。20世纪80年代, 在葛洲坝修建之后, 长江中游宜昌至城陵矶江段仍有11处四大家鱼产卵场, 产卵规模为11.14×108粒, 占长江干流总规模的25.8%[11]。但是, 在三峡水库蓄水之后, 长江中游四大家鱼的繁殖活动受到了不利的影响[12]。同时, 过度捕捞, 尤其是对仔幼鱼的捕捞, 造成了鱼类资源的急剧减少[13]。此外, 三峡水库蓄水之后造成长江中游宜昌江段四大家鱼产卵时间推迟约20d, 监利江段宜昌江段四大家鱼的产卵规模减少约75%[14]。为了有效地开展长江鱼类资源的保护工作, 有必要掌握长江中游鱼类早期资源的现状及其变化规律。
本研究通过在宜都断面开展产漂流性卵鱼类早期资源调查, 了解宜昌江段产漂流性卵鱼类的繁殖情况; 结合历史资料, 分析四大家鱼及其他鱼类的繁殖规模和时间变化; 分析水文因子对产漂流性卵鱼类优势种类繁殖的影响; 为长江中游鱼类早期资源保护工作的开展提供科学依据。
1. 材料与方法
1.1 样本的采集
2017年5月12日至7月25日和2018年5月10日至7月24日, 在葛洲坝下游宜都市清江河口上约1.5—3.0 km处设置采样断面(112°26′E, 30°24′N)(图 1)。采用产漂流性卵鱼类早期资源的调查方法。采样网具包括弶网和圆锥网, 采集方法参照曹文宣等[1]和黎明政等[14]的方法。弶网用于每日采样, 记录种类和数量。圆锥网用于采集断面数据, 计算断面系数, 每年采集一次。弶网网口呈半圆形, 网口直径1 m, 网目尺寸500 μm, 网口面积0.393 m2, 网长2 m。网后部连接集苗箱, 集苗箱长40 cm, 宽和高均为30 cm, 上部敞口, 用于收集鱼卵和仔鱼。弶网采集在每日上午和下午各一次, 采集时间为7:30—9:00和16:30—18:00, 每次采集持续30—60min。同时, 测量采样点水温、透明度、流速等环境参数, 水位和流量数据来自于全国水雨情网站(http://xxfb.mwr.cn/)。圆锥网用于断面定量采样。网口呈圆形, 网口直径0.5 m, 网口面积0.196 m2, 网长2 m, 网目尺寸500 μm。圆锥网设置在断面左岸近岸点、左岸至江中心1/2处、江中心、江中心至右岸1/2处以及右岸近岸处5个采样点, 每个采样点设置表层、中层和底层3个采样水层(表层、中层和底层距水面的距离分别占水深的20%、50%和80%), 每个水层采集15min。
![]() 图 1 2017年和2018年长江中游宜都断面鱼类早期资源采样图Figure 1. Samling map of fish eggs and larvae in the Yidu section of the Yangtze River in 2017 and 2018
图 1 2017年和2018年长江中游宜都断面鱼类早期资源采样图Figure 1. Samling map of fish eggs and larvae in the Yidu section of the Yangtze River in 2017 and 20181.2 种类鉴定
鱼卵按照卵膜性质(漂流性、黏性和浮性)分类, 并依据卵径大小、胚体长等标准将大小不同的鱼卵区分培养, 培养水温维持在20—25℃, 直至孵出发育至分类特征明显的阶段, 按照形态、色素分布、肌节数目等进行种类鉴定, 具体方法参照曹文宣等[1]; 仔鱼通过显微镜观察进行种类鉴定, 之后样本用95%的乙醇保存。形态观测不能确定种类的样本用95%的乙醇固定保存在−20℃冰箱, 带回分子实验室, 通过线粒体DNA细胞色素b基因进行种类鉴定, 具体实验方法参照Xiao等[15]的方法进行。
1.3 数据处理
产卵规模计算 鱼卵和仔鱼通过断面径流量的估算方法参照易伯鲁等[10]、黎明政等[14]和母红霞等[16]的方法进行, 具体计算公式如下:
$A = M + M'\qquad\qquad\qquad\qquad\qquad\qquad$

$M = \sum\nolimits_{i = 1}^n {{T_i}} {D_i}{Q_i}C\qquad\qquad\qquad\qquad\quad$

$\begin{aligned}{M' } =& \sum\limits_{i = 1}^{n - 1} {T_{\left( {i,i + 1} \right)}'} \times C \times \left[ {\left( {{Q_i} + {Q_{i + 1}}} \right)/2} \right] \times \;\;\;\\& \left[ {\left( {{D_i} + {D_{i + 1}}} \right)/2} \right]\end{aligned}$

${D_{{i}}} = {N_i}/\left( {S\;{V_i}\;{T_i}} \right)\qquad\qquad\qquad\qquad\qquad\;\;$

$\;C = \left( {\sum\limits_{{{j}} = 1}^m {{D_j}} } \right)/\left( {m\;{D_r}} \right)\qquad\qquad\qquad\qquad$

式中, A为整个采样期内流经采集点江段面的鱼卵、仔鱼总流量; M为所有定时采集时间内鱼卵、仔鱼流量之和; M'为前后两次相邻采集之间非采集时间内计算出的鱼卵、仔鱼流量之和; i为某一特定的常规采样; i+1为第i次采样之后的一次采样; Qi为第i次采样时流经采样断面的水流量(m3/s); Di 为第i次采样时流经采样断面的仔鱼和鱼卵密度(ind./m3); C 为采样断面上的鱼卵和仔鱼的分布系数; T (i,i+1) 为第i次采样和第i+1次采样之间的时间间隔; Ni为第i次采样中采集到的鱼卵和仔鱼数量; S为网口面积(m2); Vi为第i次采样中网口的水流速(m/s); Ti为第i次采样的持续时间(s); Dj为断面采样每个采样点的鱼卵和仔鱼密度(ind./m3); j为第j个断面采样位点; m为断面采样的采样点数量; Dr为断面采样中流经靠近右岸的采样点表层的鱼卵和仔鱼密度(ind./m3)。
确定产漂流性卵鱼类优势种类 物种优势度利用Pinkas相对重要性指数(IRI, Index of relative abundance)[17]确定, 计算公式如下:
$IRI = \left( {N{\text{%}} + W{\text{%}} } \right) \times F{\text{%}} $

式中, N为某一种类鱼卵数量占采获鱼卵总数量的百分比; W为某种鱼卵生物量比例; F为某一种类出现的次数占调查总次数的百分比。由于鱼卵、鱼苗很小, 生物量难以确定, 因此只考虑数量这一因素。将IRI大于1000的种类定义为优势种。
四大家鱼产卵场位置推算 四大家鱼产卵场位置的计算公式如下:
$L = V \times T$

式中, L为四大家鱼卵、苗的漂流距离(m); V为调查江段水流速度(m/s); T为胚胎发育所需要的时间(s)。其中, 四大家鱼胚胎发育所需要的时间参照易伯鲁等[18]。
繁殖活动的水文条件分析 根据相关研究文献资料, 近些年一段断面所采集的仔鱼大多来源于三峡大坝以上江段, 鱼卵来自三峡大坝下游产卵场[14, 19]。因此,鱼类繁殖水文因子影响分析仅针对通过鱼卵鉴定的产漂流性卵鱼类的优势种类。首先, 对物种的密度及水文因子数据进行除趋势对应分析(Detrended correspondence analysis, DCA), 根据每个轴的梯度长度(Lengths of gradient, LGA)选择适合的分析方法。当LGA<3时, 采用冗余分析(Redundancy Analysis, RDA); 当LGA>4时, 采用典型对应分析(Canonical Correlation Analysis, CCA); 当4>LGA>3时, 两种分析方法皆可[20]。DCA分析结果轴梯度长度为1.8, 故本研究利用冗余分析(Redundancy Analysis,RDA)分析鱼卵密度与环境因子(水温、流量、流量日变化、透明度)之间的相关关系。
DCA和RDA分析利用Canoco 5.0[21]软件完成, 其余数据分析与作图用Excel 2010软件中完成。
2. 结果
2.1 种类组成及优势种
2017年和2018年共采集鱼卵21120粒和仔鱼2123尾。物种鉴定结果显示, 鱼卵和仔鱼共5目9科37种。其中, 鲤科(Cyprinidae) 23种, 鳅科(Cobitidae) 5种, 鲿科(Bagridae)及科(Sisoridae)各2种, 银鱼科(Salangidae)、平鳍鳅科(Homalopteridae)、鱵科(Hemiramphidae)、鮨科(Serranidae)和鰕虎鱼科(Gobiidae)各1种, 鲤科鱼类资源数量为20788粒, 占总数量的89.44%,其中鲌亚科为优势类群, 占鲤科鱼类总数量的39.45%。根据卵的性质, 漂流性卵24种, 黏性卵5种, 浮性卵1种, 沉性卵7种(表 1)。
表 1 2017年和2018年调查期间宜都断面鱼类早期资源种类组成Table 1. Species composition of eggs and larvae in the Yidu section in 2017 and 2018目Order 科Family 种Species 卵类型Type of egg 卵Eggs 苗Larvae 目Order 科Family 种Species 卵类型Type of egg 卵Eggs 苗Larvae 鲑形目Salmoniforme
银鱼科Salangidae太湖新银鱼Neosalanx taihuensis 沉性 + 圆筒吻Rhinogobio cylindricus 漂流性 + + 鲤形目Cypriniformes
鲤科Cyprinidae青鱼Mylopharyngodon piceus 漂流性 + + 蛇Saurogobio dabryi 漂流性 + + 草鱼Ctenopharyngodon idellus 漂流性 + + 光唇蛇Saurogobio gymnocheilus 漂流性 + 赤眼鳟Squaliobarbus curriculus 漂流性 + + 宜昌鳅鮀Gobiobotia filifer 漂流性 + 鳡Elopichthys bambusa 漂流性 + + 鲤Cyprinus capio 黏性 + 飘鱼Pseudolaubuca sinensis 黏性 + 鲫Carassius auratus 黏性 + 寡鳞飘鱼Pseudolaubuca engraulis 漂流性 + + 平鳍鳅科Homalopteridae 犁头鳅Lepturichthys fimbriata 漂流性 + + 贝氏䱗Hemiculter bleekeri 漂流性 + + 鳅科Cobitidae 花斑副沙鳅Parabotia fasciata 漂流性 + + 翘嘴鲌Culter alburnus 黏性 + + 武昌副沙鳅Parabotia banarescui 漂流性 + 鳊Parabramis pekinensis 漂流性 + + 紫薄鳅Leptobotia taeniops 漂流性 + 银鲴Xenocypris argentea 漂流性 + + 红唇薄鳅Leptobotia rubrilabris 漂流性 + 黄尾鲴Xenocypris davidi 黏性 + 鲇形目Siluriformes
鲿科Bagridae瓦氏黄颡鱼Pelteobagrus vachelli 沉性 + 似鳊Pseudobrama simoni 漂流性 + 光泽黄颡鱼Pelteobaggrus nitidus 沉性 + 鳙Hypophthalmichthys nobilis 漂流性 + + 科Sisoridae 中华纹胸Glyptothorax sinensis 沉性 + 鲢Hypophthalmichthys molitrix 漂流性 + + 福建纹胸Glyptothorax fukiensis 沉性 + 银Squalidus argentatus 漂流性 + + 颌针鱼目Beloniformes
鱵科Hemiramphidae间下鱵Hyporhamphus intermedius 沉性 + 铜鱼Coreius heterodon 漂流性 + + 鲈形目Perciformes
鮨科Serranidae鳜Siniperca chuatsi 浮性 + + 吻Rhinogobio typus 漂流性 + 鰕虎鱼科Gobiidae 子陵吻鰕虎鱼Rhinogobius giurinus 沉性 + 注: “+”代表可采集到Note: “+” means can be collected 2017年和2018年采集到鱼卵28种, 有青鱼(Mylopharyngodon piceus)、草鱼(Ctenopharyngodon idellus)、赤眼鳟(Squaliobarbus curriculus)、贝氏䱗(Hemiculter bleekeri)、鳙(Hypophthalmichthys nobilis)、鲢(Hypophthalmichthys molitrix)和银(Squalidus argentatus)等种类。采集到仔鱼27种, 有太湖新银鱼(Neosalanx taihuensis)、寡鳞飘鱼(Pseudolanbuca engraulis)、贝氏䱗、间下鱵(Hyporhamphus intermedius)和子陵吻鰕虎鱼(Rhinogobius giurinus)等种类。
2017年, 采集鱼卵10572粒, 仔鱼1221尾, 其中四大家鱼卵3172粒, 贝氏䱗卵2725粒, 分别占卵总数量的30.0%、25.8%; 仔鱼以贝氏䱗和寡鳞飘鱼为主, 分别占比54.3%、24.4%。2018年共采集鱼卵10548粒, 仔鱼902尾, 其中四大家鱼卵4249粒, 贝氏䱗卵2448粒, 分别占卵总数量的40.3%和23.2%; 仔鱼以子陵吻鰕虎鱼、寡鳞飘鱼和贝氏䱗为主, 分别占比22.7%、19.3%和17.7%。根据2017年和2018年采集卵和仔鱼数量分析, 宜都断面鱼卵优势种为四大家鱼(IRI=1433)和贝氏䱗(IRI=1319); 仔鱼优势种为贝氏䱗(IRI=1259)和寡鳞飘鱼(IRI=1169)。
2.2 产卵时间
2017年5月采集鱼卵以银和飘鱼为主, 占当月总卵数的27.13%和19.38%, 6月四大家鱼和贝氏䱗成为主要种类, 数量占比分别为38.89%和26.12%。在7月四大家鱼数量占比下降至18.50%, 而贝氏䱗上升至43.24%。2018年稍有不同, 四大家鱼数量百分比自5月的55.38%逐渐下降至7月的15.04%, 贝氏䱗相对比例依旧在6月达到最高, 占当月数量的42.77%, 7月保持稳定, 占当月数量的41.00%。2017年和2018年调查期间, 飘鱼和银所占数量比自5月至7月逐渐减少, 而花斑副沙鳅和鳊则相反, 自5月比例逐渐上升。2017年四大家鱼的产卵时间是从5月14日到7月2日。2018年则是从5月10日至7月5日。
仔鱼5月采集较少, 2017年5月仅采集仔鱼5尾, 分别为间下鱵、光泽黄颡鱼和光唇蛇, 2018年5月采集仔鱼16尾, 分别为太湖新银鱼、寡鳞飘鱼和贝氏䱗。仔鱼采集数量主要在6月和7月。2017年和2018年采集结果显示, 仔鱼中贝氏䱗和寡鳞飘鱼为优势种类(图 2)。
![]() 图 2 2017年和2018年宜都断面主要类群鱼卵(a)、仔鱼(b)的相对丰度Figure 2. Abundance percentage of eggs (a) and larvae (b) of dominant species in the Yidu section in 2017 and 2018
图 2 2017年和2018年宜都断面主要类群鱼卵(a)、仔鱼(b)的相对丰度Figure 2. Abundance percentage of eggs (a) and larvae (b) of dominant species in the Yidu section in 2017 and 20182.3 产卵规模
2017年, 通过宜都断面的鱼卵径流量为124.45×108粒, 鱼卵中四大家鱼径流量为36.13×108粒, 占总径流量的29.03%; 贝氏䱗径流量为19.96×108粒, 占总径流量的16.04%。2018年通过宜都断面的鱼卵径流量为101.07×108粒, 鱼卵中四大家鱼径流量为35.20×108粒, 占总径流量的34.83%; 贝氏䱗的鱼卵径流量为19.17×108粒, 占总径流量的18.97%。
2017年通过宜都断面的仔鱼径流量为16.43×108尾, 寡鳞飘鱼和贝氏䱗径流量分别为5.89×108尾和2.91×108尾, 占仔鱼总径流量的35.87%和17.72%。四大家鱼的总径流量为4.12×108尾, 占仔鱼总径流量的25.07%。2018年通过宜都断面的仔鱼径流量为8.29×108尾, 寡鳞飘鱼和贝氏䱗分别为0.97×108尾和1.22×108尾, 占仔鱼总径流量的11.72%和14.70%。四大家鱼的总径流量为0.11×108尾, 占仔鱼总径流量的1.32%。
2.4 四大家鱼的产卵场位置
通过对采集到的四大家鱼卵发育期的观测, 0.14%处于128细胞期前, 95.31%处于桑椹期至眼囊期, 4.55%处于尾芽期至心脏搏动期, 距受精1—33h。其中, 65.76%的四大家鱼卵处于原肠早期至原肠晚期, 距受精7—13h。四大家鱼仔鱼均处于鳔雏形期和鳔一室期, 距受精98—146h。根据鱼卵和仔鱼的发育期以及流速分析, 四大家鱼鱼卵来源于宜都断面2.4 km以上的江段。产卵场位置主要分布在葛洲坝下至庙咀、胭脂坝至云池江段。四大家鱼仔鱼应来源于三峡大坝以上江段(图 3)。
![]() 图 3 长江中游宜昌江段四大家鱼产卵场位置Figure 3. Spawning grounds of the four major Chinese carps in the Yichang reach in the middle Yangtze River
图 3 长江中游宜昌江段四大家鱼产卵场位置Figure 3. Spawning grounds of the four major Chinese carps in the Yichang reach in the middle Yangtze River2.5 产漂流性卵鱼类及关键环境因子
2017年最小流量在7月4日, 为6920 m3/s, 最大流量在7月11日, 为30200 m3/s。整个调查期间, 水温介于18.5—26.0℃, 5月12日达到18℃以上, 7月20日达到24℃以上。2017年出现4次产卵高峰, 分别在6月10日、6月11日、6月18日和7月10日。高峰期产卵规模总计为60.7×108粒, 占2017年总产卵规模的48.7%。
2018年6月12日流量最小, 为10200 m3/s, 最大流量在7月14日, 为43800 m3/s。整个调查期间, 水温为18.5—25.2℃, 5月10日达到18℃以上, 7月2日达到24℃以上。2018年出现2次显著的产卵高峰, 分别为5月19日和6月24日, 产卵规模共为35.9×108粒, 占2018年采集鱼卵总规模的35.5%(图 4)。
![]() 图 4 2017年(a)和2018年(b)调查期间宜都断面鱼卵日密度变化图Figure 4. Discharge and eggs density drifting through the Yidu section in 2017 (a) and 2018 (b)
图 4 2017年(a)和2018年(b)调查期间宜都断面鱼卵日密度变化图Figure 4. Discharge and eggs density drifting through the Yidu section in 2017 (a) and 2018 (b)应用RDA分析方法, 对鱼卵漂流密度与水文因子的关系进行分析。在前2个排序轴上, 物种与水文因子的相关性为0.426和0.345, 第一排序轴和第二排序轴的累积贡献率为92.83%。RDA分析结果显示, 四大家鱼卵总密度和花斑副沙鳅卵密度受流量日变化和流量影响较大; 贝氏䱗鱼卵密度受流量影响较大; 鳊鱼卵密度受水温影响较大; 银鱼卵密度主要受透明度影响(图 5)。
![]() 图 5 2017年和2018年宜都断面水文因子对漂流性卵密度的影响Figure 5. Effects of hydrological factors on the density of the drifting eggs in the Yidu section in 2017 and 2018
图 5 2017年和2018年宜都断面水文因子对漂流性卵密度的影响Figure 5. Effects of hydrological factors on the density of the drifting eggs in the Yidu section in 2017 and 20183. 讨论
3.1 长江中游鱼类繁殖现状及变化
本研究结果显示, 2017年和2018年5—7月通过长江中游宜都断面的鱼类早期资源较丰富。已有的研究结果显示, 母红霞[22]发现2009年和2010年5—7月通过宜都断面鱼卵径流量分别为57.26×108粒和64.45×108粒, 年均60.86×108粒。与2009年和2010年同比, 2017年和2018年通过宜都断面的鱼卵径流量明显增加, 年均径流量增加了85.3%。其中, 宜昌江段四大家鱼的产卵规模明显增大。2005—2012年5—7月通过宜都断面四大家鱼鱼卵径流量为(0.03—5.85)×108粒, 年均2.49×108粒[14], 而2017年和2018年四大家鱼卵年均35.67×108粒, 同比增加了13倍, 达到了20世纪80年代的水平[10, 11]。鱼卵径流量增加的主要原因可能有2个。第一, 2009年起, 原农业部在长江中游开展较大规模的四大家鱼亲鱼放流, 增加了繁殖群体数量[7, 23]。第二, 2011年三峡工程开始生态调度试验, 在一定程度上促进了鱼类的繁殖活动。徐薇等[24]的研究结果显示, 2012年三峡水库试验性生态调度期间长江中游沙市江段鱼卵总径流量和家鱼卵径流量均出现高峰, 繁殖种类明显增加。周雪等[25]的研究结果显示, 2013—2017年三峡水库生态调度造成长江中游监利江段涨水持续时间达到4d及以上, 期间四大家鱼的卵苗量较大, 占年总量的45%以上。
在三峡水库蓄水运行之后, 长江中游四大家鱼的产卵时间推迟。在三峡水库蓄水之前, 长江中游宜昌江段四大家鱼产卵时间从4月中下旬开始[10]。在蓄水后, 2005—2012年宜昌江段四大家鱼的产卵时间推迟到5月8日至5月28日, 平均推迟了21.8d, 导致产卵时间推迟的主要原因是下泄水温滞后, 到达18℃的时间推迟[14]。与2005—2012年相比, 2017年和2018年宜昌江段四大家鱼产卵时间没有明显变化, 产卵场位置也没有明显变化。因此, 本研究的结果表明, 在三峡水库蓄水运行之后, 长江中游四大家鱼产卵场的环境条件可能已经趋于稳定。如果亲本增加, 宜昌江段产卵场依然可以满足其完成繁殖活动, 产生较大规模的鱼卵和仔鱼。
本研究结果还显示, 2017年和2018年5—7月有一定规模的仔鱼通过长江中游宜都断面, 其中四大家鱼的仔鱼均来源于三峡大坝以上江段。《长江三峡水利枢纽环境影响报告书》认为, 在三峡水库蓄水之后, 长江上游涪陵以下的8个四大家鱼产卵场将消失, 产卵场将上移。根据母红霞[22]、王红丽等[26]、Li等[14]和马琴[27]在三峡水库库区的鱼类早期资源调查结果显示, 涪陵以下江段仍有产卵规模较小的四大家鱼的产卵场。2005—2012年, 通过宜都断面四大家鱼仔鱼径流量年均值为4.65×108尾[14]。2013年和2014年分别为1.52×108尾和1.27×108尾[27]。与2005—2014年相比, 2017年和2018年没有明显的变化。研究结果表明, 三峡水库库区内在一定水文条件下仍具备四大家鱼繁殖的条件, 且可以通过三峡大坝进入长江中游, 可以为长江中游鱼类资源提供一定的补充。
3.2 鱼类早期资源与环境因子的关系
本研究结果显示, 四大家鱼、贝氏䱗和花斑副沙鳅等种类的产卵规模与流量和流量日变化有着密切的关系。鱼类的繁殖活动受外界环境因子的影响[28]。已有的研究提出“最佳环境窗口”假说, 认为每个物种都有特定的环境条件需求, 以满足繁殖活动和保证产生最多的后代[29—31]。Li等[32]的研究结果显示, 四大家鱼的产卵活动需要18—24℃的水温和0.55 m/d的水位涨幅。Wang等[12]也发现, 四大家鱼的仔鱼丰度与流量增加持续时间显著相关。因此, 水温应该是四大家鱼等产漂流性卵鱼类的繁殖限制条件, 达到了一定水温鱼类才能开始繁殖活动[10, 33]。流量等则是刺激鱼类繁殖和促进繁殖活动的重要环境条件[10, 34]。
3.3 长江中游鱼类早期资源保护措施
鱼类早期资源减少导致种群补充量减少[35, 36], 是造成长江鱼类资源急剧减少的主要原因之一[1, 37]。Ma等[38]的研究结果显示, 长江中游对于通江湖泊中鱼类早期资源的补充, 维持鱼类多样性有重要意义。为了长江鱼类资源保护, 2019年农业农村部、财政部和人力资源社会保障部联合印发《长江流域重点水域禁捕和建立补偿制度实施方案》, 长江禁渔已经逐步实施, 2021年长江将全面禁渔。这一措施将消除捕捞对鱼类资源的影响, 尤其是对湖区等仔幼鱼资源的影响, 能够显著提高鱼类早期生活史阶段的存活率, 增加种群的补充量。长江鱼类的早期生活史阶段还面临着栖息地减少的问题。长江中下游的湖泊和河漫滩原本是仔幼鱼的主要栖息地。但是, 长江中下游的通江湖泊仅剩石臼湖、洞庭湖和鄱阳湖, 其余湖泊都已经被阻隔, 仔幼鱼无法进入湖泊生长, 其栖息地功能已经完全丧失。Liu等[39]的研究结果显示, 长江中下游江湖连通性减少已经严重影响了鱼类多样性。同时, 港口建设、防洪护坡和城市景观等建设破坏了河漫滩的自然环境条件, 减少了仔幼鱼的栖息地[40, 41]。因此, 为了更加有效地开展鱼类资源保护, 结合本研究结果, 还建议通过恢复江湖连通、灌江纳苗, 以及维持河漫滩自然环境条件, 增加仔幼鱼的栖息地面积; 开展重要经济鱼类亲本放流, 增加繁殖群体规模; 开展生态调度, 促进繁殖活动。
致谢:
感谢中国科学院水生生物研究所胡华明、李文静、杨萍、张文武和张晨在样本采集、数据处理上给予的帮助!
-
![]()
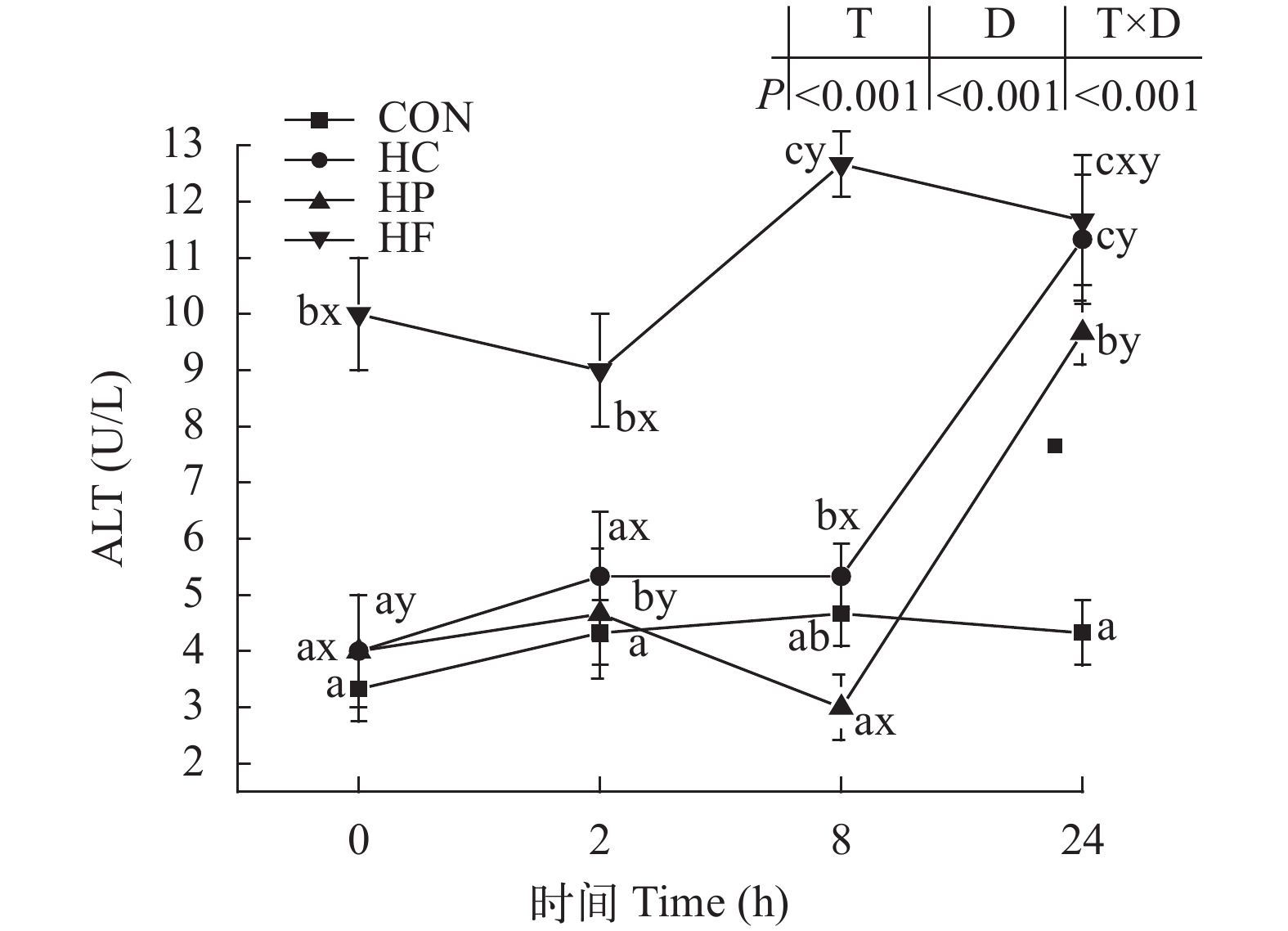
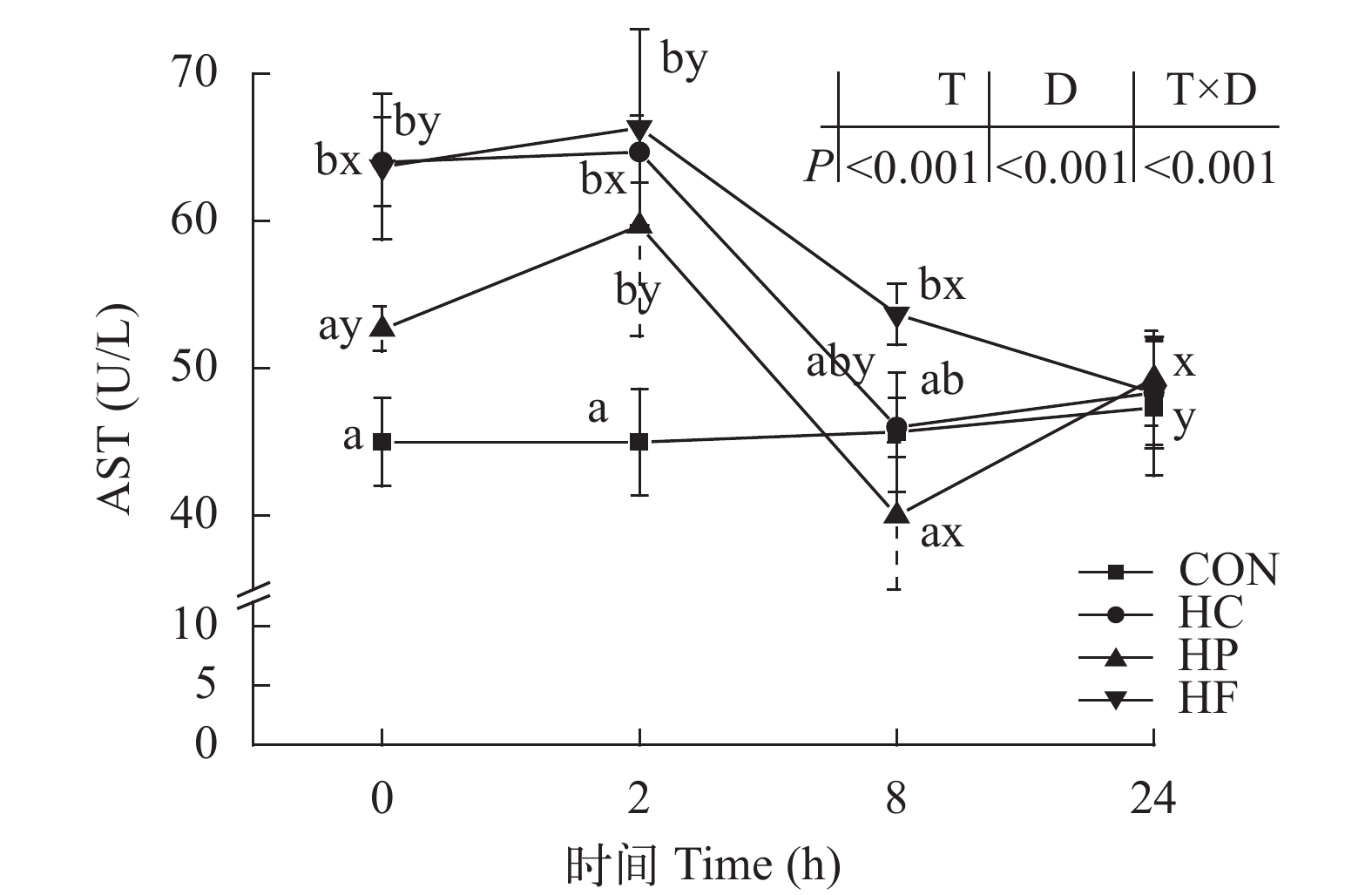
图 1 高能量饲料对草鱼餐后血清谷草转氨酶活性的影响
Figure 1. Effect of high-energy diets on postprandial AST activities in serum of grass carp
![]()
图 2 高能量饲料对草鱼餐后血清谷丙转氨酶活性的影响
Figure 2. Effect of high-energy diets on postprandial ALT activities in serum of grass carp
![]()
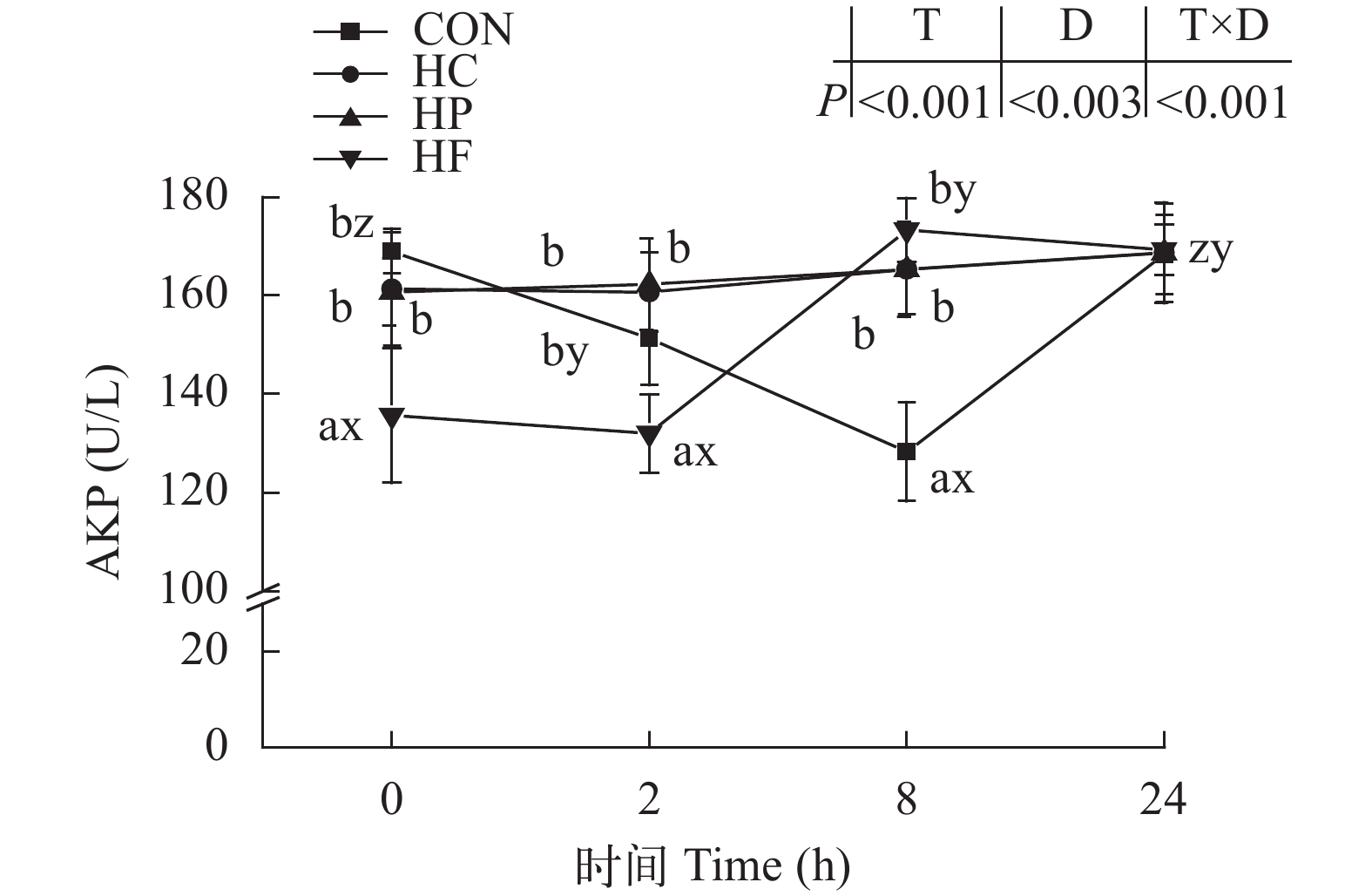
图 3 高能量饲料对草鱼餐后血清碱性磷酸酶活性的影响
Figure 3. Effect of high-energy diets on postprandial ALP activities in serum of grass carp
![]()
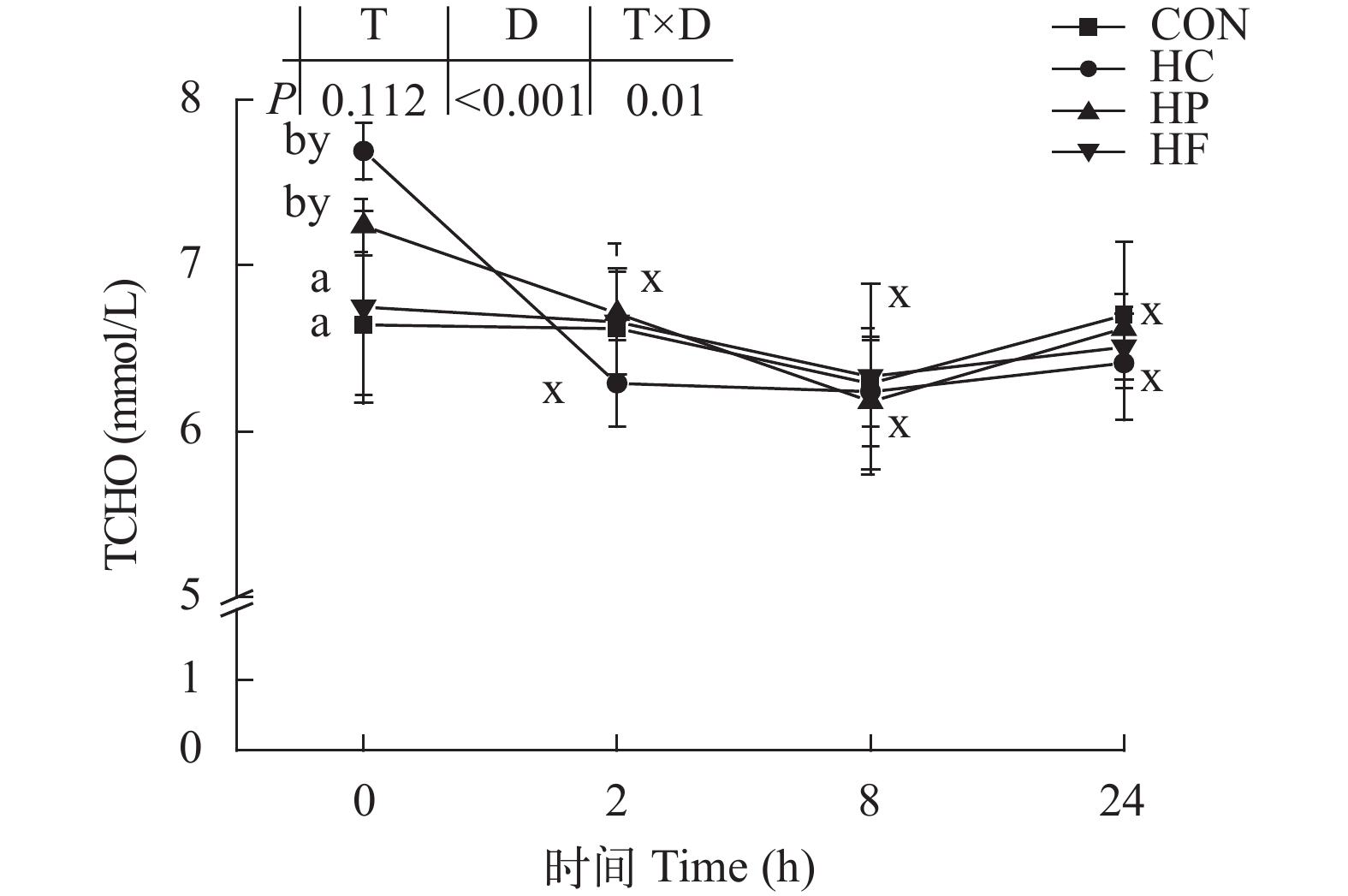
图 4 高能量饲料对草鱼餐后血清胆固醇含量的影响
Figure 4. Effect of high-energy diets on postprandial TCHO contents in serum of grass carp
![]()
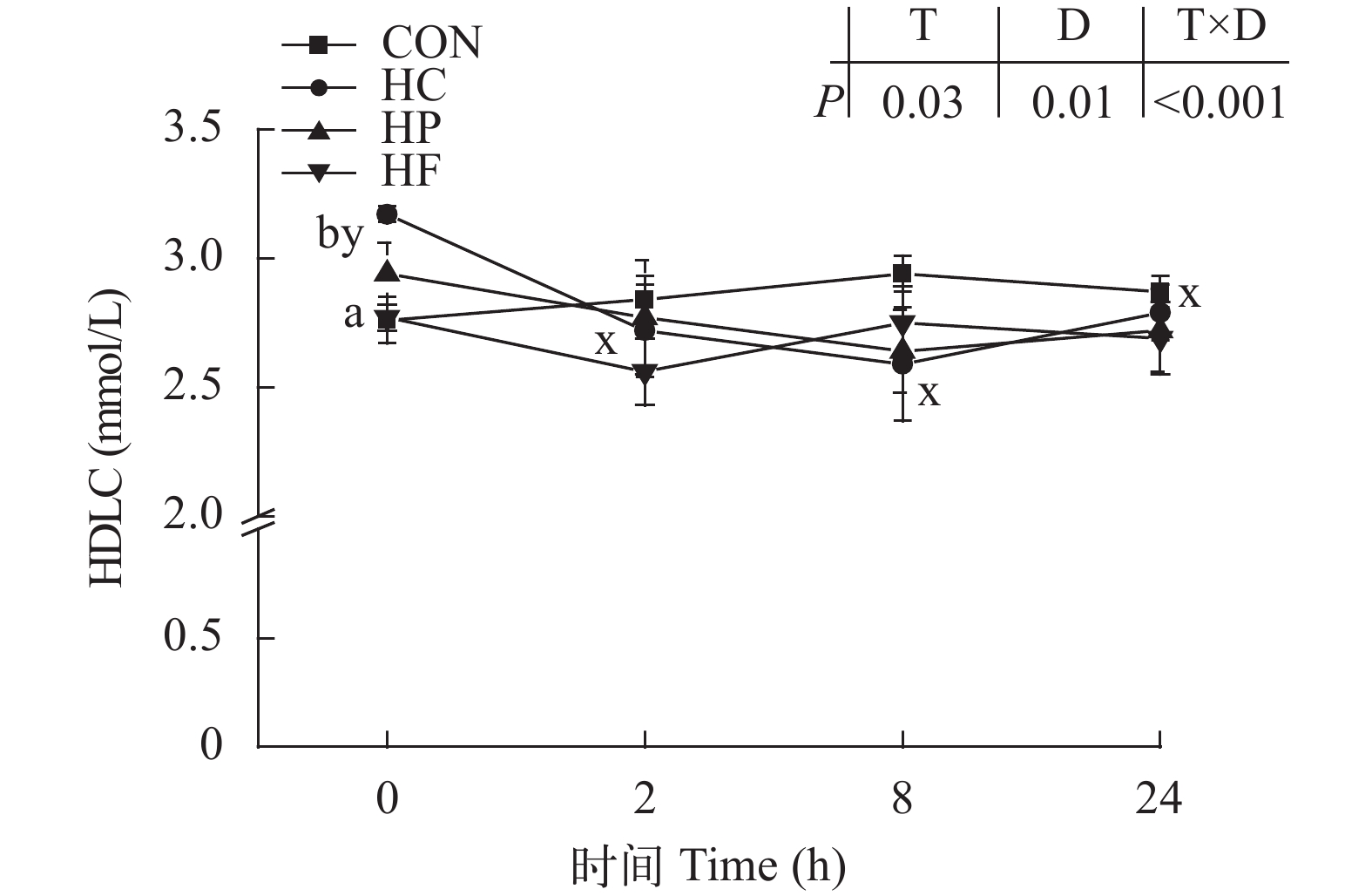
图 5 高能量饲料对草鱼餐后血清高密度脂蛋白胆固醇含量的影响
Figure 5. Effect of high-energy diets on postprandial HDLC contents in serum of grass carp
![]()
图 6 高能量饲料对草鱼餐后血清低密度脂蛋白胆固醇含量的影响
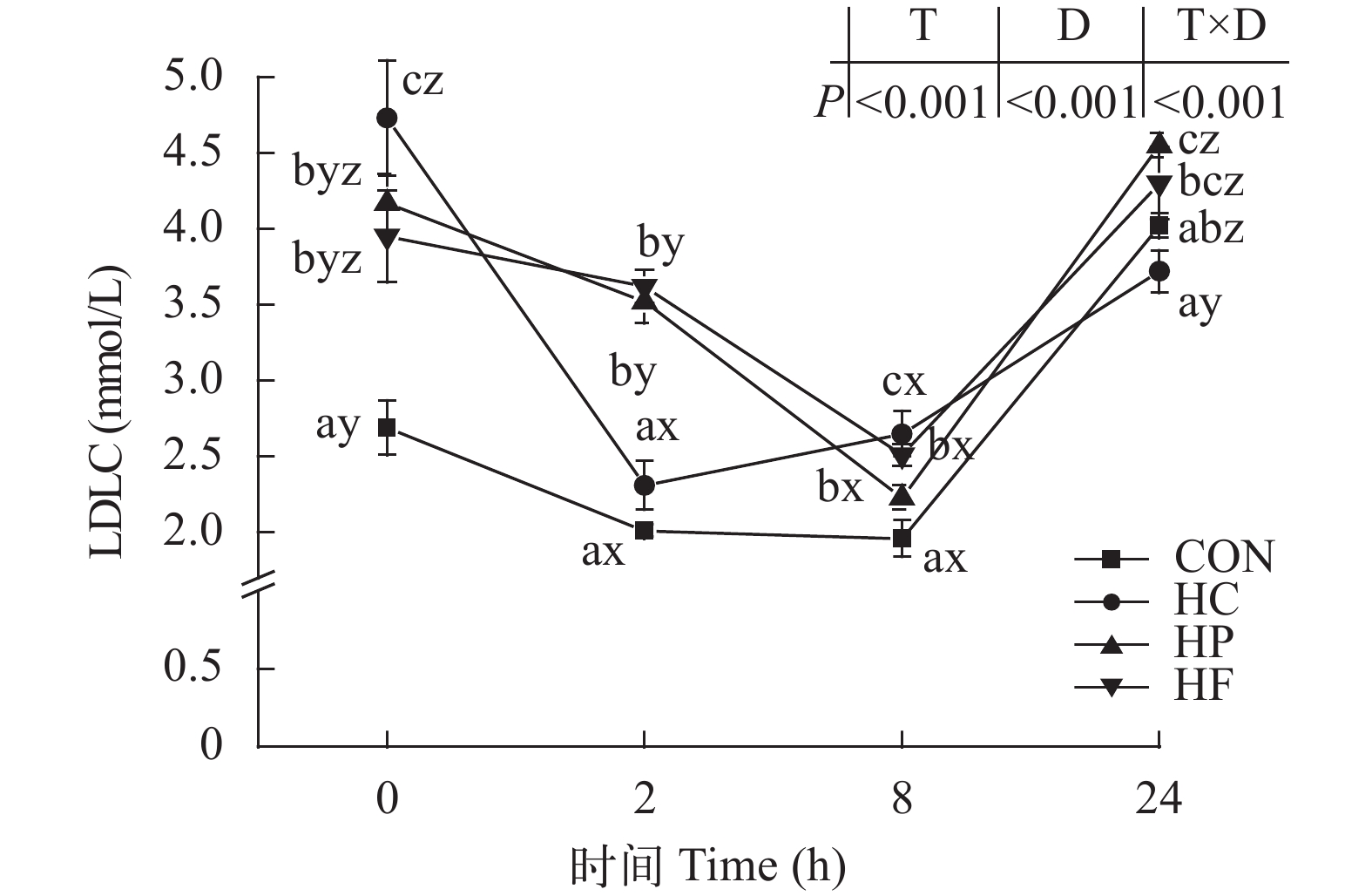
Figure 6. Effect of high-energy diets on postprandial LDLC contents in serum of grass carp
![]()
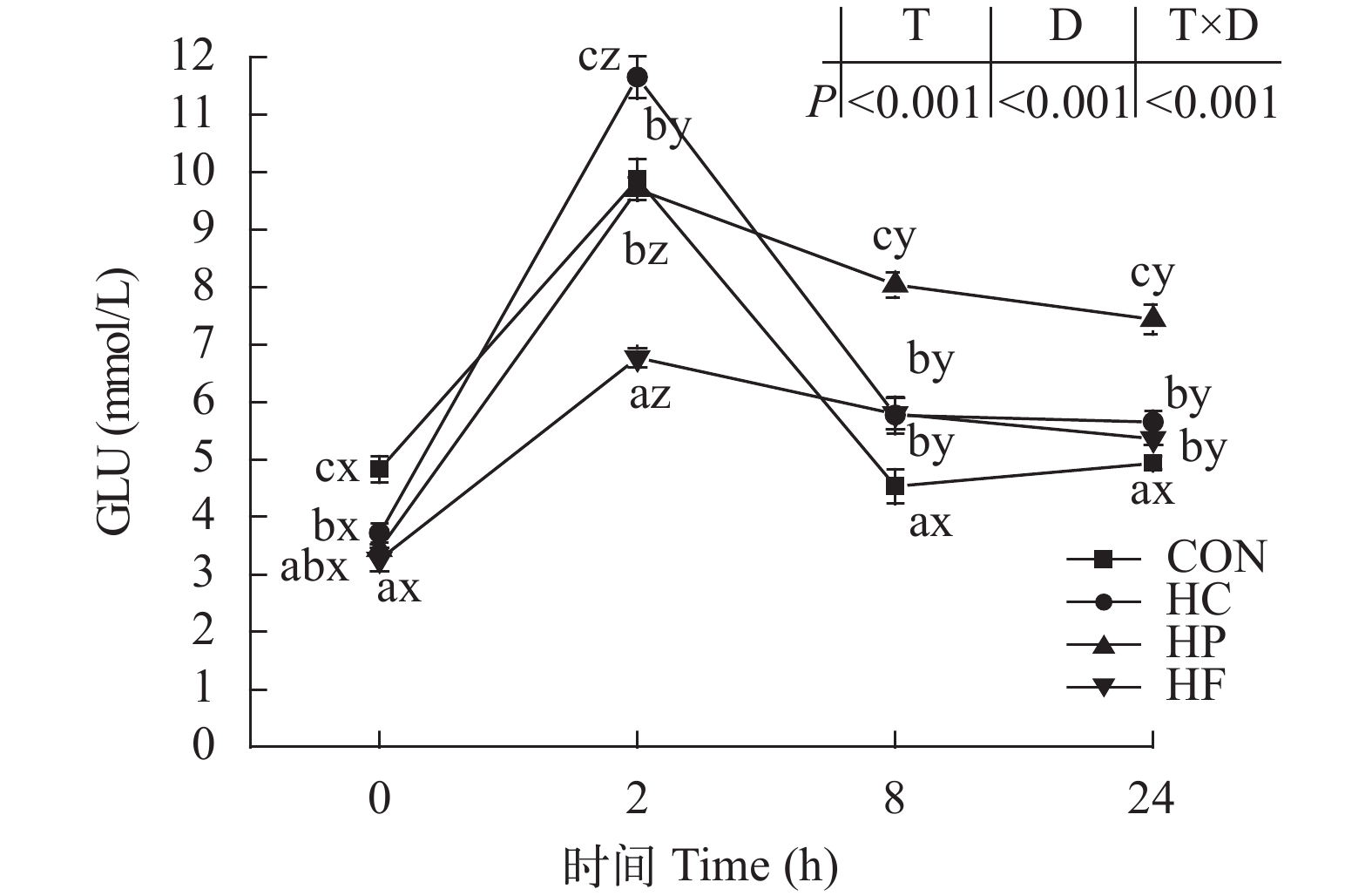
图 7 高能量饲料对草鱼餐后血清葡萄糖含量的影响
Figure 7. Effect of high-energy diets on postprandial GLU contents in serum of grass carp
![]()
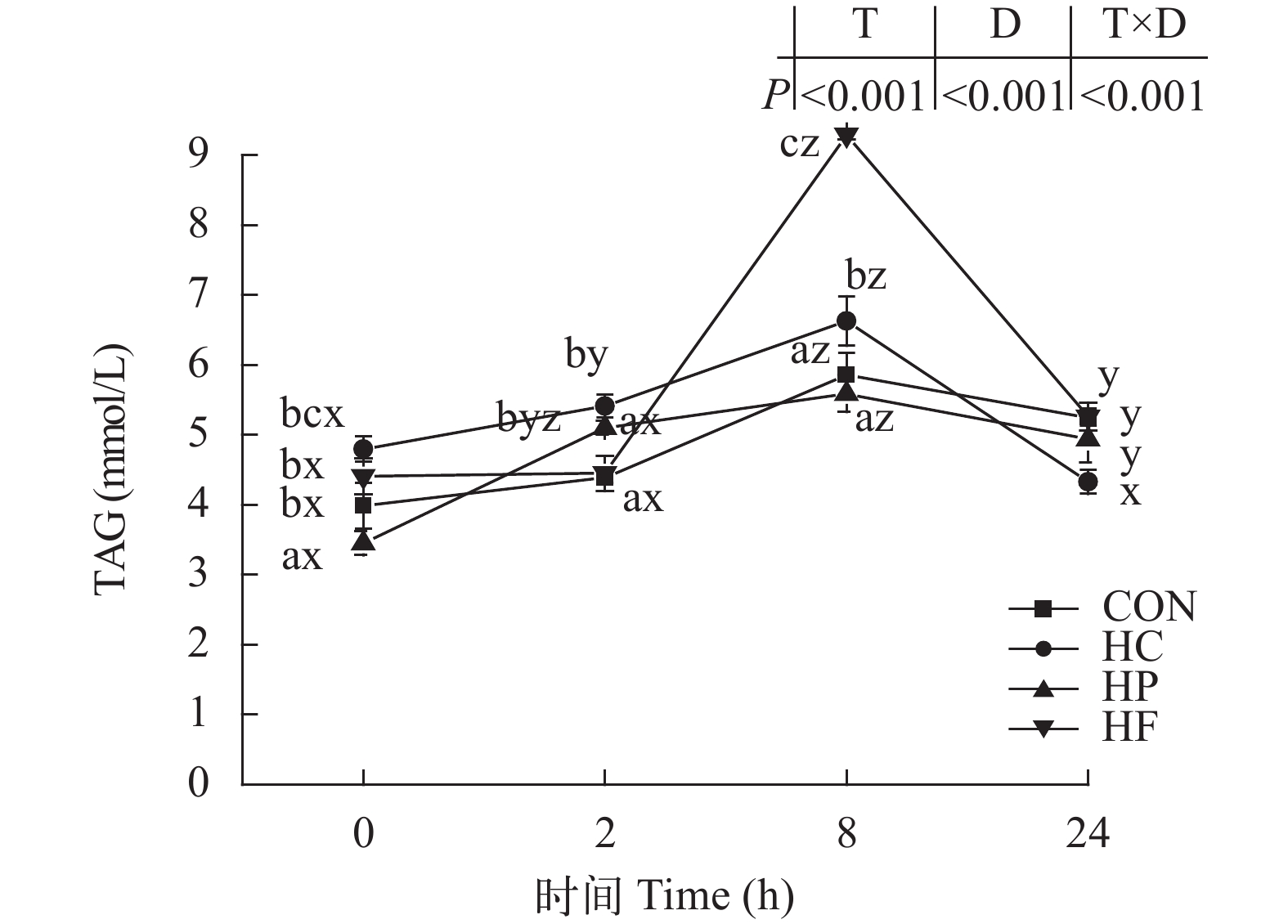
图 8 高能量饲料对草鱼餐后血清甘油三酯含量的影响
Figure 8. Effect of high-energy diets on postprandial TAG contents in serum of grass carp
表 1 实验饲料组成和基本分析
Table 1 Composition and proximate analysis of the experimental diets
原料成分Ingredient 饲料Dietary (g/kg diet) CON HP HF HC 酪蛋白Casein 280 380 280 280 明胶Gelatin 70 95 70 70 豆油和玉米油Soybean oil and corn oil (1﹕1) 80 80 140 80 糊精Dextrin 356.5 356.5 356.5 486.5 纤维素Cellulose 151 26 91 20 复合维生素Vitamin premix1 10 10 10 10 复合矿物盐Mineral premix1 50 50 50 50 氯化胆碱Choline chloride 2.5 2.5 2.5 2.5 合计Total 1000 1000 1000 1000 基本营养成分Proximate composition (g/kg diet) 干物质Dry matter 948.5 945.2 961.6 948.8 粗蛋白Crude protein 302.3 418.1 298.2 308.2 粗脂肪Crude lipid 76.7 76.1 135.4 78.1 灰分Ash 48.5 50.5 48.2 49.1 总能Gross energy (kJ/g)2 17.2 19.4 19.9 19.1 注: 1. 复合维生素和复合矿物盐配方参照田娟等[23]的实验; 2. 饲料总能用氧胆量热仪测定(SDC311, 湖南三德科技股份有限公司)
Note: 1. The vitamin and mineral premix followed previous study by Tian, et al[23]; 2. Energy was determined by direct combustion in an adiabatic bomb calorimeter (SDC311, Hunan Sundy Science and Technology Development Co., Ltd, Changsha, Hunan Province, China) 下载: 导出CSV
下载: 导出CSV
-
[1] John R P, Parkinson E A. Energy Allocation Strategy in Young Fish: Allometry and Survival [M]. 2001, 1040—1051
[2] Lin D, Mao Y, Cai F. Nutritional lipid liver disease of grass carp Ctenopharyngodon idullus (C. et V.) [J]. Chinese Journal of Oceanology and Limnology, 1990, 8(4): 363—373 doi: 10.1007/BF02849682
[3] 杜震宇. 养殖鱼类脂肪肝成因及相关思考. 水产学报, 2014, 38(9): 1628—1638 Du Z Y. Causes of fatty liver in farmed fish: a review and new perspectives [J]. Journal of Fisheries of China, 2014, 38(9): 1628—1638
[4] Watanabe T. Strategies for further development of aquatic feeds [J]. Fisheries Science, 2002, 68(2): 242—252 doi: 10.1046/j.1444-2906.2002.00418.x
[5] Tocher D R. Metabolism and functions of lipids and fatty acids in teleost fish [J]. Reviews in Fisheries Science, 2003, 11(2): 107—184 doi: 10.1080/713610925
[6] 陈毅坚. 血液的功能. 玉溪师范学院学报, 1993, 3: 46—50 Chen Y J. The function of blood [J]. Journal of Yuxi Normal University, 1993, 3: 46—50
[7] 朱心玲, 贾丽珠, 张明瑛. 草鱼血液学的研究Ⅰ.九项血液常数的周年变化. 水生生物学报, 1985, 9(3): 248—257 Zhu X L, Jia L Z, Zhang M Y. Haematological studies on the grass carp Ⅰ. year-round changes in nine haematological parameters [J]. Acta Hydrobiologica Sinica, 1985, 9(3): 248—257
[8] 陈洁, 熊邦喜, 顾钱洪, 等. 内源及外源因子对鱼类血液生理生化指标的影响. 湖北农业科学, 2011, 50(9): 1861—1865 doi: 10.3969/j.issn.0439-8114.2011.09.041 Chen J, Xiong B X, Gu Q H, et al. The effect of endogenous and environment factors on hematology physiological and biochemical indices of fish [J]. Hubei Agricultural Sciences, 2011, 50(9): 1861—1865 doi: 10.3969/j.issn.0439-8114.2011.09.041
[9] 王爱民, 韩光明, 封功能, 等. 饲料脂肪水平对吉富罗非鱼生产性能、营养物质消化及血液生化指标的影响. 水生生物学报, 2011, 35(1): 80—87 Wang A M, Han G M, Feng G N, et al. Effects of dietary lipid levels on growth performance, nutrient digestibility and blood biochemical indices of gift tilapia (Oreochromis niloticus) [J]. Acta Hydrobiologica Sinica, 2011, 35(1): 80—87
[10] 郭小泽, 梁旭方, 方刘, 等. 饲料中非蛋白能量源对草鱼血清生化指标和肝脏组织的影响. 水生生物学报, 2014, 38(3): 582—587 Guo X Z, Liang X F, Fang L, et al. Effects of non-protein energy sources on serum biochemical indices and histology of liver in grass carp (Ctenopharyngodon idella) [J]. Acta Hydrobiologica Sinica, 2014, 38(3): 582—587
[11] 梁拥军, 孙砚胜, 苏建通, 等. 饲料脂肪水平对锦鲤幼鱼生长和血液生化指标的影响. 淡水渔业, 2012, 42(5): 49—53 doi: 10.3969/j.issn.1000-6907.2012.05.010 Liang Y J, Sun Y S, Su J T, et al. Effects of different lipid levels on growth and haematological biochemistry in juvenile tilapia (Oreochromis niloticus×Oreochromis aureus) [J]. Freshwater Fisheries, 2012, 42(5): 49—53 doi: 10.3969/j.issn.1000-6907.2012.05.010
[12] 刘泓宇, 毛义波, 谭北平, 等. 饲料糖水平对不同食性鱼类生长及葡萄糖耐受能力的影响. 水产学报, 2015, 39(12): 1852—1862 Liu H Y, Mao Y B, Tan B P, et al. The effects of dietary lipid levels on growth and blood biochemical indices of Cyprinus carpio L [J]. Journal of Fisheries of China, 2015, 39(12): 1852—1862
[13] Tian J, Wen H, Zeng L B, et al. Changes in the activities and mRNA expression levels of lipoprotein lipase (LPL), hormone-sensitive lipase (HSL) and fatty acid synthetase (FAS) of Nile tilapia (Oreochromis niloticus) during fasting and re-feeding [J]. Aquaculture, 2013, 400—401: 29—35
[14] 向枭, 陈建, 周兴华, 等. 5种脂肪源对齐口裂腹鱼生长性能及血清生化指标的影响. 动物营养学报, 2010, 22(2): 498—504 doi: 10.3969/j.issn.1006-267x.2010.02.038 Xiang X, Chen J, Zhou X H, et al. Effects of five lipid sources on growth performance and serum biochemical indices of Schizothorax prenanti [J]. Chinese Journal of Animal Nutrition, 2010, 22(2): 498—504 doi: 10.3969/j.issn.1006-267x.2010.02.038
[15] 刘伟, 文华, 蒋明, 等. 2种蛋白水平的饲料循环投喂对奥尼罗非鱼幼鱼生长、体成分和血清生化指标的影响. 华南农业大学学报, 2013, 34(3): 405—410 Liu W, Wen H, Jiang M, et al. Effects of mixed feeding schedules with two dietary protein levels on growth performance, body composition and serum biochemical indexes of junvenile hybrid tilapia (Oreochromis niloticus ♀×O.aureus ♂) [J]. Journal of South China Agricultural University, 2013, 34(3): 405—410
[16] 吉红, 李杰. 草鱼脂质营养与饲料学研究进展. 饲料工业, 2010, s1: 50—54 doi: 10.3969/j.issn.1003-6202.2010.10.016 Ji H, Li J. Research progress in lipid nutrition and feed of grass carp [J]. Feed Industry, 2010, s1: 50—54 doi: 10.3969/j.issn.1003-6202.2010.10.016
[17] 刘兴旺, 张海涛. 草鱼营养生理研究进展. 饲料博览, 2013, (2): 20—24 doi: 10.3969/j.issn.1001-0084.2013.02.005 Liu X W, Zhang H T. The Advances of nutrition physiology in grass carp [J]. Feed Review, 2013, (2): 20—24 doi: 10.3969/j.issn.1001-0084.2013.02.005
[18] 冯硕恒, 米海峰, 刘迎隆, 等. 草鱼对饲料中碳水化合物利用的研究进展. 中国饲料, 2016, (5): 25—28 Feng S H, Mi H F, Liu Y L, et al. The Advances of carbohydrate utilization in grass carp [J]. China Feed, 2016, (5): 25—28
[19] Köprücü K. Effects of dietary protein and lipid levels on growth, feed utilization and body composition of juvenile grass carp (Ctenopharyngodon idella) [J]. Journal of Fisheries Sciences, 2012, 6(3): 377—380
[20] Jin Y, Tian L X, Zeng S L, et al. Dietary lipid requirement on non-specific immune responses in juvenile grass carp (Ctenopharyngodon idella) [J]. Fish & Shellfish Immunology, 2013, 34(5): 1202—1208
[21] 田丽霞, 刘永坚, 冯健, 等. 不同种类淀粉对草鱼生长、肠系膜脂肪沉积和鱼体组成的影响. 水产学报, 2002, 26(3): 247—251 Tian L X, Liu Y J, Feng J, et al. Effect of different types of starch on growth, the deposition of mesenteric fat and body composition of Ctenopharyngodon idellus [J]. Journal of Fisheries of China, 2002, 26(3): 247—251
[22] 田娟, 高攀, 蒋明, 等. 饲料蛋白能量比对草鱼幼鱼生长性能、蛋白利用和体成分的影响. 淡水渔业, 2016, 46(4): 83—90 doi: 10.3969/j.issn.1000-6907.2016.04.014 Tian J, Gao P, Jiang M, et al. Effect of dietary protein to energy ratios on growth performance, protein efficiency and body composition of juvenile Ctenopharyngodon idellus [J]. Freshwater Fisheries, 2016, 46(4): 83—90 doi: 10.3969/j.issn.1000-6907.2016.04.014
[23] Tian J, Liu W, Gao W, et al. Molecular cloning and gene/protein expression of FAT/CD36 from grass carp (Ctenopharyngodon idella) and the regulation of its expression by dietary energy [J]. Fish Physiology and Biochemistry, 2017, 43(3): 875—888 doi: 10.1007/s10695-017-0342-7
[24] Xu D, He G, Mai K, et al. Postprandial nutrient-sensing and metabolic responses after partial dietary fishmeal replacement by soyabean meal in turbot (Scophthalmus maximus L.) [J]. British Journal of Nutrition, 2016, 115(3): 379—388 doi: 10.1017/S0007114515004535
[25] Karlsson A, Eliason E J, Mydland L T, et al. Postprandial changes in plasma free amino acid levels obtained simultaneously from the hepatic portal vein and the dorsal aorta in rainbow trout (Oncorhynchus mykiss) [J]. Journal of Experimental Biology, 2006, 209(24): 4885—4894 doi: 10.1242/jeb.02597
[26] 冷向军, 田娟, 陈丙爱, 等. 罗非鱼对晶体蛋氨酸, 包膜蛋氨酸利用的比较研究. 水生生物学报, 2013, 37(2): 235—242 Leng X J, Tian J, Chen B A, et al. Comparison study on utilizing crystalline or coated methionine by tilapia (Oreochromis niloticus×O. aureus) [J]. Acta Hydrobiologica Sinica, 2013, 37(2): 235—242
[27] 王爱民, 韩光明, 吕富, 等. 饲料脂肪水平对吉富罗非鱼幼鱼摄食后血脂、血糖的影响. 盐城工学院学报(自然科学版), 2011, 24(3): 9—14 doi: 10.3969/j.issn.1671-5322.2011.03.003 Wang A M, Han G M, Lü F, et al. Effects of dietary lipid levels on blood lipid, glucose indexes of GIFT tilapia (Oreochromis niloticus) [J]. Journal of Yancheng Institute of Technology (
Natural Science Edition ) , 2011, 24(3): 9—14 doi: 10.3969/j.issn.1671-5322.2011.03.003 [28] 麻晓林, 陈伟, 金榕兵. 肝损伤后检测肝功能指标的实验研究. 创伤外科杂志, 2003, 5(1): 20—22 doi: 10.3969/j.issn.1009-4237.2003.01.008 MaX L, Chen W, Jin R B. Study of examination of liver function following liver injury [J]. Journal of Traumatic Surgery, 2003, 5(1): 20—22 doi: 10.3969/j.issn.1009-4237.2003.01.008
[29] 汪开毓, 苗常鸿, 黄锦炉, 等. 投喂高脂饲料后草鱼主要生化指标和乙酰辅酶A羧化酶1 mRNA表达的变化. 动物营养学报, 2012, 24(12): 2375—2383 doi: 10.3969/j.issn.1006-267x.2012.12.012 Wang K Y, Miao C H, Huang H L, et al. Changes in main biochemical indices and Acetyl-CoA carboxylase 1 mRNA expression of crass carp fed a high-fat diet [J]. Chinese Journal of Animal Nutrition, 2012, 24(12): 2375—2383 doi: 10.3969/j.issn.1006-267x.2012.12.012
[30] 吴凡, 文华, 蒋明, 等. 饲料碳水化合物水平对吉富罗非鱼幼鱼生长性能和血液主要生化指标的影响. 西北农林科技大学学报(自然科学版), 2012, 40(12): 8—14 Wu F, Wen H, Jiang M, et al. Effects of different dietary carbohydrate level on growth performance and blood biochemical parameters of juvenile GIFT tilapia (Oreochromis niloticus) [J]. Journal of Northwest A & F University (
Natural Science Edition ) , 2012, 40(12): 8—14 [31] 吴凡, 文华, 蒋明, 等. 饲料碳水化合物水平对奥尼罗非鱼幼鱼生长, 体成分和血清生化指标的影响. 华南农业大学学报, 2011, 32(4): 91—95 doi: 10.3969/j.issn.1001-411X.2011.04.020 Wu F, Wen H, Jiang M, et al. Effects of different dietary carbohydrate levels on growth performance, body composition and serum biochemical indices of juvenile hybrid tilapia (Oreochromis niloticus ♀×O. aureus ♂) [J]. Journal of South China Agricultural University, 2011, 32(4): 91—95 doi: 10.3969/j.issn.1001-411X.2011.04.020
[32] 丁立云, 张利民, 王际英, 等. 饲料蛋白水平对星斑川鲽幼鱼生长、体组成及血浆生化指标的影响. 中国水产科学, 2010, 17(6): 1285—1292 Ding L Y, Zhang L M, Wang J Y, et al. Effects of dietary protein level on growth performance, body composition and plasma biochemistry indices of juvenile starry flounder, Platichthys stellatus [J]. Journal of Fishery Sciences of China, 2010, 17(6): 1285—1292
[33] 张辉, 张海莲. 碱性磷酸酶在水产动物中的作用. 河北渔业, 2003, 5: 12—13—32 Zhang H, Zhang H L. The role of alkaline phosphatase in aquatic animals [J]. Hebei Fisheries, 2003, 5: 12—13—32
[34] 缪凌鸿, 刘波, 戈贤平, 等. 高碳水化合物水平日粮对异育银鲫生长、生理、免疫和肝脏超微结构的影响. 水产学报, 2011, 35(2): 221—230 Miao L H, Liu B, Ge X P, et al. Effcet of high carbohydrate levels in the dietary on growth performance,immunity and transmission electron microscopy (TEM) on hepatic cell of allogynogenetic crucian carp (Carassius auratus gibelio) [J]. Journal of Fisheries of China, 2011, 35(2): 221—230
[35] 戈贤平, 刘波, 谢骏, 等. 饲料中不同碳水化合物水平对翘嘴红鲌生长及血液指标和糖代谢酶的影响. 南京农业大学学报, 2007, 30(3): 88—93 Ge X P, Liu B, Xie J, et al. Effect of different carbohydrate levels of dietary on growth, plasma biochemical indices and hepaticpancreas carbohydrate metabolic enzymes in topmouth culter (Erythroculter ilishaeformis Bleeker) [J]. Journal of Nanjing Agricultural University, 2007, 30(3): 88—93
[36] 林小植, 罗毅平, 谢小军. 饲料碳水化合物水平对南方鲇幼鱼餐后糖酵解酶活性及血糖浓度的影响. 水生生物学报, 2006, 30(3): 304—310 doi: 10.3321/j.issn:1000-3207.2006.03.009 Lin X Z, Luo Y P, Xie X J. Effects of dietary carbohydrate level on glycolytic enzymes and serum glucose concentrations in the juvenile southern catfish after feeding [J]. Acta Hydrobiologica Sinica, 2006, 30(3): 304—310 doi: 10.3321/j.issn:1000-3207.2006.03.009
[37] 宋理平, 韩勃, 王爱英, 等. 碳水化合物水平对厚唇弱棘䱨生长和血液指标的影响. 大连海洋大学学报, 2010, 25(4): 293—297 doi: 10.3969/j.issn.1000-9957.2010.04.003 Song L P, Han B, Wang A Y, et al. The effects of dietary carbohydrate levels on growth performance and plasma biochemical indices in Hephaestus fuliginosus [J]. Journal of Dalian Fisheries University, 2010, 25(4): 293—297 doi: 10.3969/j.issn.1000-9957.2010.04.003
[38] Tian J, Wu F, Yang C G, et al. Dietary lipid levels impact lipoprotein lipase, hormone-sensitive lipase, and fatty acid synthetase gene expression in three tissues of adult GIFT strain of Nile tilapia, Oreochromis niloticus [J]. Fish Physiology and Biochemistry, 2015, 41(1): 1—18 doi: 10.1007/s10695-014-0001-1
[39] 叶坤, 王秋荣, 谢仰杰, 等. 饲料脂肪水平对黄姑鱼幼鱼生长性能、肌肉组成和血浆生化指标的影响. 动物营养学报, 2017, 29(4): 1418—1426 doi: 10.3969/j.issn.1006-267x.2017.04.041 Ye K, wang Q R, Xie Y J, et al. Effects of dietary lipid level on growth performance, muscle composition and plasma biochemical indices of juvenile Nibea albiflora [J]. Chinese Journal of Animal Nutrition, 2017, 29(4): 1418—1426 doi: 10.3969/j.issn.1006-267x.2017.04.041
[40] 杜震宇. 草鱼对脂肪的利用和对脂肪酸β-氧化的研究. 中山大学. 2005, 23—41 Du Z Y. Consequences of fat feeding on growth and body lipids in a herbivorous fish (grass carp, Ctenopharyngodon idella): mechanisms related to fatty acid oxidation [D]. Sun Yat-sen University. 2005, 23—41
[41] Riche M, Haley D I, Oetker M, et al. Effect of feeding frequency on gastric evacuation and the return of appetite in tilapia Oreochromis niloticus (L.) [J]. Aquaculture, 2004, 234(1): 657—673
[42] Sveier H, Wathne E, Lied E. Growth, feed and nutrient utilisation and gastrointestinal evacuation time in Atlantic salmon (Salmo salar L.): the effect of dietary fish meal particle size and protein concentration [J]. Aquaculture, 1999, 180(3): 265—282
[43] Fauconneau B, Choubert G, Blanc D, et al. Influence of environmental temperature on flow rate of foodstuffs through the gastrointestinal tract of rainbow trout [J]. Aquaculture, 1983, 34(1): 27—39
-
期刊类型引用(20)
1. 米玮洁,张舜,胡俊,高少波,胡菊香. 黄河内蒙古河段水文情势变化及生态效应初探. 水生态学杂志. 2024(01): 42-49 .  百度学术
百度学术
2. 班学君,樊博,刘瀚,俞立雄,林俊强,夏继红,张迪. 鱼类产卵行为与生态水文指标响应关系研究——以长江四大家鱼为例. 水生态学杂志. 2024(01): 67-74 . 百度学术
3. 李莎,姜伟,李博,党莹超,苏巍,戴凌全. 长江葛洲坝坝下江段鱼类群落结构及多样性. 水生态学杂志. 2024(03): 121-130 . 百度学术
4. 张云昌,丁洋,赵进勇. 河流伦理建构中的八个理论和实践问题探讨. 中国水利. 2024(16): 7-11 . 百度学术
5. 易燃,陶江平,杨志,刘宏高,徐薇,曹俊,张鹏. 改进的栖息地法求解多种产漂流性卵鱼类生态流量方法研究. 中国农村水利水电. 2024(09): 45-51+59 . 百度学术
6. 蒋祥龙,黎明政,杨少荣,林鹏程,常涛,王春伶,张晨,高欣. 鄱阳湖鱼类多样性的时空变化特征研究. 水生生物学报. 2023(03): 376-388 . 本站查看
7. 周月娟,高雷,段辛斌,姜伟,黄安阳,王珂,陈大庆,汪登强. 基于耳石微结构的长江监利断面草鱼和鲢仔鱼群体日龄结构研究. 长江流域资源与环境. 2023(02): 324-330 . 百度学术
8. 徐薇,金瑶,陈桂亚,董纯,易燃,赵娜,陶江平. 三峡水库十年生态调度(2011—2020年)期间下游沙市江段产漂流性卵鱼类自然繁殖变化. 湖泊科学. 2023(05): 1729-1741 . 百度学术
9. 高雷,郭国忠,杨浩,陈大庆,王珂,段辛斌. 长江中游洪湖江段四大家鱼早期资源及生态调度影响. 中国水产科学. 2023(12): 1556-1565 . 百度学术
10. 高欣,赵亚辉,田菲,王晓爱,黎明政,林鹏程,常涛,俞丹,刘焕章. 中国内陆水体鱼类多样性监测专项网的监测和研究进展. 生物多样性. 2023(12): 199-210 . 百度学术
11. 霍堂斌,宋聃,刘伟,唐富江,王继隆,张澜澜,李喆,赵荣伟. 松花江下游富锦江段鱼类早期资源状况. 中国水产科学. 2022(01): 91-101 . 百度学术
12. 彭云鑫,方弟安,徐东坡,丁隆强,黎加胜,任鹏,何晓辉,罗宇婷. 长江安庆段草鱼仔稚鱼资源时空动态特征与水文因子的相关性. 中国水产科学. 2022(02): 314-324 . 百度学术
13. 蒋祥龙,黎明政,杨少荣,林鹏程,王春伶,张晨,高欣. 鄱阳湖鱼类集合群落结构特征及其时间变化研究. 长江流域资源与环境. 2022(03): 588-601 . 百度学术
14. 李鹏程,荣义峰,杜浩,王普渊,刘文成,刁亚芹. 基于深度学习的青海湖裸鲤产卵场遥感识别方法. 中国水产科学. 2022(03): 398-407 . 百度学术
15. 王小豪,方弟安,孙海博,何苗,薛向平. 长江如皋江段仔稚鱼资源现状调查分析. 南方农业学报. 2022(06): 1742-1751 . 百度学术
16. 向浩,林枫,伍遇普,田波,蔡志宇,吴益平,黄盼君,刘千凡,周志国. 长江中游宜昌段浮游动物群落结构特征与环境因子相关性分析. 黑龙江水产. 2022(05): 3-12 . 百度学术
17. 汤正阳,蒋爱萍,林俊强,任玉峰,张迪,刘新波,杨靖,赵庆绪. 长江流域水工程生态环境保护研究进展. 水利水电技术(中英文). 2022(S2): 297-309 . 百度学术
18. 祁涛,班璇,杜浩,郭文献,杜耘,龙奥运,南璐艺,石小涛,郑楚洋. 长江中游鱼类敏感水文期内水文情势对其资源量的影响评估. 长江流域资源与环境. 2022(12): 2621-2632 . 百度学术
19. 胡春明,董熙,尤立,李曜. 红花尔基水利枢纽对下游河道水文情势影响研究. 干旱区资源与环境. 2021(06): 89-95 . 百度学术
20. 王慧,张建新,田翊霏,陈君,耿硕,杨元昊,王立新. 湑水河国家级水产种质资源保护区鱼类资源调查与评价. 家畜生态学报. 2021(11): 54-59 . 百度学术
其他类型引用(6)





计量
- 文章访问数: 1942
- HTML全文浏览量: 659
- PDF下载量: 79
- 被引次数: 26