
EFFECTS OF ENZYMATIC HYDROLYZED SOYBEAN MEAL ON GROWTH PERFORMANCE, LIVER FUNCTION AND METABOLISM OF LARGEMOUTH BASS (MICROPTERUS SALMOIDES)
-
摘要: 研究旨在探究酶解豆粕替代鱼粉对大口黑鲈(Micropterus salmoides)的生长性能、消化酶活力、肝脏抗氧化能力以及代谢率的影响, 以植物蛋白复合物(酶解豆粕 鲶玉米蛋白粉=10 鲶1)替代部分鱼粉, 酶解豆粕的添加量分别为0 (E0组)、15% (E15组)、20% (E20组)、25% (E25组)和30% (E30组), 各组替代鱼粉的量分别为0、23.64%、30.91%、40%和47.27%, 并在E20组(替代30.91%的鱼粉)的基础上, 将添加量为20%的酶解豆粕改用豆粕(SBM组)或发酵豆粕(FSBM组)等蛋白替代鱼粉, 配制等氮等能的试验饲料。将初始体重为(17.17±0.14) g的大口黑鲈随机分为7组, 分别投喂7种不同的试验饲料, 养殖周期为67d。结果显示: (1) E25和E30的特定生长率和增重率显著高于其他组(P<0.05), 各组的饲料系数无显著性差异。E25和E30组的存活率较低, 可能与摄食不均衡, 小个体被残杀有关, 这也可能造成E25和E30组的生长性能显著高于其他组。随着酶解豆粕添加量的增加, 脏体比、肝体比、全鱼脂肪含量显著降低(P<0.05); 豆粕、酶解豆粕、发酵豆粕分别替代30.91%鱼粉时, FSBM组的特定生长率显著低于E20组(P<0.05), 脏体比、肝体比显著高于其他两组(P<0.05)。(2) 随着酶解豆粕替代鱼粉的比例不断增大, 肠道淀粉酶和脂肪酶活力呈先上升后下降的趋势, 均高于对照组; E20和E30组的胃蛋白酶活力显著高于对照组(P<0.05); 3种豆粕分别替代30.91%的鱼粉时, FSBM组肠道淀粉酶活力显著高于其他两组(P<0.05), 肠道脂肪酶活力的变化趋势则相反。(3) 肝脏总抗氧化能力(T-AOC)、谷草转氨酶(AST)、谷丙转氨酶(ALT)显著高于对照组(P<0.05, 除E30组肝脏ALT外), 肝脏丙二醛(MDA)呈下降趋势; E20、SBM与FSBM三组中, FSBM组的肝脏MDA、ALT显著低于其他两组(P<0.05), 肝脏AST活力依次为E20>FSBM>SBM; (4) 在酶解豆粕替代鱼粉后, 各组的耗氧率有显著升高的趋势(P<0.05), E20、E25和E30组的排氨率显著高于对照组(P<0.05), E20和E30组肌肉氮的保留率显著高于其他组(P<0.05); 酶解豆粕替代鱼粉对大口黑鲈血清的游离脂肪酸(NEFA)、总胆固醇(T-CHO)、甘油三酯(TG)和低密度脂蛋白胆固醇(LDL-C)都有显著的影响。3种不同豆粕分别替代30.91%鱼粉时, FSBM组的耗氧率显著低于E20和SBM组 (P<0.05), 排氨率显著高于其他两组(P<0.05); 豆粕组(SBM组)的血清T-CHO显著高于其他两组, 而血清LDL-C与肌肉脂肪含量呈相反的结果; E20组的血清TG显著低于其他两组。综上所述, 饲料中酶解豆粕的添加量小于30%时, 对大口黑鲈的生长无不利影响, 但有利于减轻肝脏氧化应激负担, 并提高代谢水平。普通豆粕、酶解豆粕与发酵豆粕均可以替代30.91%鱼粉, 且酶解豆粕的替代效果最好。Abstract: To investigate the effects of enzymatic hydrolyzed soybean meal (ESBM) on growth performance, liver function and metabolism of largemouth bass (Micropterus salmoides), five isonitrogenous and isoenergetic diets were formulated by replacing fish meal with a plant-based protein source compound (enzymatic hydrolyzed soybean meal: corn gluten meal=10 鲶1) for a 67-days trial. The additions of ESBM in the feed were 0 (E0), 15% (E15), 20% (E20), 25% (E25), 30% (E30) to replace 0, 23.64%, 30.91%, 40%, 47.27% of the fish meal, respectively. In additional, two diets of soybean meal (SBM) and fermented soybean meal (FSBM) were used as the substitute for 20% ESBM, respectively. The results showed that the specific growth rate and weight gain rate of E25 and E30 were significantly higher than those of other groups (P<0.05), and that the feed coefficient ratio of each group had no significant difference (P>0.05). The survival rates of the E25 and E30 groups were lower than other groups. With the increase of ESBM, the viscerosomatic index, hepatic index and the lipid content of body decreased significantly (P<0.05). The specific growth rate of FSBM group was significantly lower than that of E20 group (P<0.05), and viscerosomatic index and hepatic index of FBSM were significantly higher than those of SBM and E20 (P<0.05). The activities of amylase and lipase of intestines increased first and then decreased with the increased ESBM, which were significantly higher than that of the control group (P<0.05). The activity of pepsin in E20 and E30 group was significantly higher than that of the control group (P<0.05). The activity of amylase in FSBM group was significantly higher than SBM and E20 (P<0.05), while the change of intestinal lipase activity was opposite. The activities of liver total-antioxidant capacity (T-AOC), glutamic oxaloacetic transaminase (AST) and alanine aminotransferase (ALT) in all group except liver ALT in E30 group were significantly induced by ESBM (P<0.05), and ESBM significantly reduced liver malondialdehyde (MDA) (P<0.05). The liver MDA content and ALT activities of FSBM group was significantly lower than SBM and E20 group (P<0.05), and the liver AST activity was in the order of E20>FSBM>SBM. The oxygen consumption increased significantly with the increased ESBM, and the nitrogen excretion in E20, E25 and E30 were significantly higher than E0 group (P<0.05). The muscle nitrogen retention rates of E20 and E30 were higher than other groups. ESBM had significant effects on serum free fatty acid (NEFA), total cholesterol (T-CHO), triglyceride (TG), and low density lipoprotein cholesterol (LDL-C) of largemouth bass. Among E20, FSBM and SBM groups, the oxygen consumption in FSBM group was significantly lower than other two groups (P<0.05), while nitrogen excretion was opposite. The serum T-CHO of SBM group was significantly higher than other two groups (P<0.05), while serum LDL-C and muscle lipid were the contrary trend. The serum TG of E20 group was significantly lower than other two groups (P<0.05). These results indicated that the addition of ESBM up to 30% did not harm the growth, and reduced liver oxidative stress to enhance nutrients metabolism. All FSBM, SBM and ESBM have benefits to replace 30.91% fish meal with the best effects by ESBM.
-
鱼粉作为水产饲料中非常重要的蛋白源, 其蛋白含量高, 氨基酸的种类齐全且成分接近全鱼鱼体氨基酸组成, 抗营养因子少, 具有未知的促生长因子[1]。由于水产养殖规模的不断扩大, 水产饲料工业的快速发展, 对鱼粉的需求量越来越大, 造成鱼粉供不应求, 价格持续上涨[2]。面对这一严峻现状, 解决鱼粉的供需矛盾, 降低饲料成本, 已经成为水产饲料行业亟待解决的问题。豆粕粗蛋白含量较高, 必需氨基酸含量丰富, 来源广泛, 因此成为了水产饲料中最重要的植物性蛋白源之一, 但是由于存在抗营养因子, 使得限制了豆粕的使用量[3]。具体表现为鱼类生长缓慢, 摄食率降低, 饲料系数升高, 饲料利用率降低[4], 并且高替代水平对鱼类的肝脏组织和肠道组织结构有破坏作用[5, 6]、影响鱼类对营养物质的消化吸收等。因此, 将豆粕处理成适合动物的优质蛋白质饲料具有非常重要的意义。酶解豆粕即是利用现代生物技术将大豆蛋白通过蛋白酶酶解降解为可溶性蛋白和小分子多肽的混合物。经过酶解处理的蛋白比传统大豆中蛋白质更易于吸收、低抗原等特点, 是水产动物饲料的理想植物蛋白[7]。研究发现, 酶解豆粕适宜的替代量, 可以促进星斑川鲽(Platichthys stellatus Pallas)幼鱼的生长, 提高饲料利用并对体组织脂肪的蓄积有一定的改善作用[8]。
大口黑鲈(Micropterus salmoides), 俗名加州鲈, 隶属鲈形目(Perciformes), 太阳鱼科(Cehtrachidae), 黑鲈属(Micropterus), 为肉食性温水鱼类, 原产于美国加利福尼亚州。大口黑鲈因具有适应性强、生长快、病害少、适温性广和肉质鲜美等优点, 已成为我国淡水鱼养殖的主要品种之一。目前, 大口黑鲈全程专用人工配合饲料在我国尚未广泛应用[9], 多以冰鲜杂鱼和其他动物性饲料为主, 因而饲料利用率较低, 这对海洋资源是一种浪费, 同时造成养殖环境的污染, 也容易引起各种疾病的爆发。研究表明, 大口黑鲈对饲料中蛋白质的需求较高, 一般超过 40%, 并对饲料中鱼粉依赖性较强, 鱼粉对蛋白的贡献率应不低于50%[10]。在鱼粉供需矛盾日渐突出的形势下, 探究鱼粉的替代对大口黑鲈生长及健康的影响有利于大口黑鲈饲料的持久发展。为此, 本研究用不同水平的酶解豆粕部分替代鱼粉, 探究大口黑鲈生长性能、体组成、消化酶活性、抗氧化能力以及蛋白质、脂类、糖类代谢的变化, 并与豆粕和发酵豆粕相比较, 以此确定酶解豆粕对鱼粉的适宜替代量, 为大口黑鲈人工配合饲料的配方优化提供依据。
1. 材料与方法
1.1 试验饲料
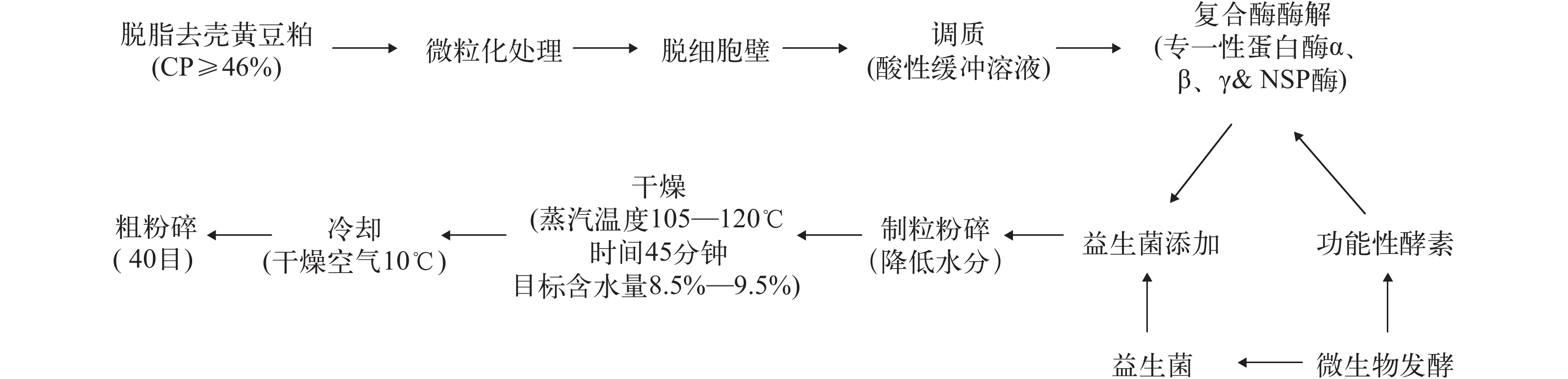
试验所用酶解豆粕由泰州福益生物有限公司提供, 其具体加工工艺如图 1所示。
酶解豆粕水溶性氮的分子量委托江南大学分析测试中心测定, 分子量分布见表 1。
表 1 酶解豆粕中水溶性氮的分子量分布Table 1. Molecular mass distribution of water-soluble nitrogen in enzymatic hydrolyzed soybean meal分子量范围Molecular mass 百分比Percentage (%) >5000 10.21 5000—3000 4.43 3000—2000 4.22 2000—1000 8.32 1000—500 11.49 500—180 16.63 <180 44.71 根据大口黑鲈的营养需求[11]配制试验饲料。以优质鱼粉、血粉、豆粕为蛋白源, 鱼油与菜籽油为脂肪源, 以微晶纤维素和沸石粉作为填充剂, 补充晶体蛋氨酸、赖氨酸和牛磺酸, 配制等蛋等脂的配合饲料。在饲料中, 以复合物(酶解豆粕 鲶玉米蛋白粉=10 鲶1)替代部分鱼粉。在五组试验饲料中酶解豆粕的添加量分别为0、15%、20%、25%和30%(组别分别为E0、E15、E20、E25和E30), 各组替代鱼粉的量分别为0、23.64%、30.91%、40%和47.27%, 并在E20组(替代30.91%的鱼粉)的基础上, 将添加量为20%的酶解豆粕改用豆粕(SBM组)或发酵豆粕(FSBM组)等蛋白替代鱼粉。饲料原料过60目筛, 在充分混匀后, 经饲料机制成直径为2 mm的颗粒, 50℃烘干至水分含量低于10%, 密封–20℃保存。试验饲料组成及营养成分见表 2。
表 2 饲料组成及营养水平(%干物质基础)Table 2. Composition and nutrient levels of diets (% DM basis)指标Index 组别Group E0 E15 E20 E25 E30 SBM FSBM 鱼粉Fish meal 55 42 38 33 29 38 38 酶解豆粕Enzymatic hydrolyzed soybean meal 0 15 20 25 30 0 0 玉米蛋白粉Corn powder 3 4.5 5 5.5 6 5 5 豆粕Soybean meal 9 9 9 9 9 29.9 9 发酵豆粕Fermented soybean meal 0 0 0 0 0 0 19 血粉Dried blood 4 4 4 4 4 4 4 生粉Cornstarch 9 9 9 9 9 9 9 鱼油Fish oil 3 4 4.2 4.4 4.7 4.2 4.2 菜籽油Rap oil 3 3 3 3 3 3 3 多维Vitamins premix* 1 1 1 1 1 1 1 多矿Minerals premix** 0.5 0.5 0.5 0.5 0.5 0.5 0.5 磷酸二氢钙Ca(H2PO4)2 1 1 1 1 1 1 1 乌贼膏Squid paste 1 1 1 1 1 1 1 胆碱Choline 0.5 0.5 0.5 0.5 0.5 0.5 0.5 羧甲基纤维素CMC 6.67 3.54 2.25 1.69 0.41 1.65 2.35 沸石粉Zeolite powder 3.33 1.77 1.13 0.85 0.2 0.83 2.03 晶体蛋氨酸Crystal methionine 0 0 0.1 0.1 0.1 0.1 0.1 晶体赖氨酸Crystal lysine 0 0.1 0.2 0.3 0.4 0.2 0.2 牛磺酸Taurine 0 0.09 0.12 0.16 0.19 0.12 0.12 合计Total 100 100 100 100 100 100 100 营养水平Nutrient level 粗蛋白Crude protein (%) 46.74 46.48 46.62 46.08 46.22 46.63 46.12 粗脂肪Crude lipid (%) 11.03 11.22 11.18 11.06 11.11 11.20 11.46 总能Gross energy (MJ/kg) 18.70 19.13 19.25 19.29 19.45 19.21 18.96 替代鱼粉比例Replacement ratio of fish meal (%) 0 23.64 30.91 40 47.27 30.91 30.91 注: 总能计算公式[12]为: 总能(MJ/kg)=蛋白质×23.6+脂肪×39.5+无氮浸出物×17.6; *多维(IU或mg/kg干饲料): 维生素A, 16000 IU; 维生素D3, 800 IU; 维生素K3, 14.72; 维生素B1, 17.8; 维生素B2, 48; 维生素B6, 29.52; 维生素Bl2, 0.24; 维生素E, 160; 维生素C (35%), 800; 烟酸胺, 79.2; 泛酸钙, 73.6; 叶酸, 6.4; 生物素, 0.64; 肌醇, 320; 氯化胆碱, 1500; L-肉碱, 100; **多矿(mg/kg干饲料): 铜(CuSO4), 2.0; 锌(ZnSO4), 34.4; 锰(MnSO4), 6.2; 铁(FeSO4), 21.1; 碘(Ca(IO3)2), 1.63; 硒(Na2SeO3), 0.18; 钴(CoCl2), 0.24; 镁(MgSO4·H2O), 52.7
Note: Gross energy was calculated using the following formula: Gross energy (MJ/kg)= protein×23.6+ lipid×39.5+NFE×17.6; *Vitamin premix (IU or mg/kg dry diet): vitamin A, 16000 IU; vitamin D3, 8000 IU; vitamin K3, 14.72; thiamin, 17.8; ribo flavine, 48; pyridoxine, 29.52; cyanocobalamine, 0.24; tocopherol acetate, 160; ascorbic acid (35%), 800; niacinamide, 79.2; calcium-D-pantothenate, 73.6; folic acid, 6.4; biotin, 0.64; inositol, 320; choline chloride, 1500; L-carnitine, 100; **Mineral premix (mg/kg dry diet): Cu (CuSO4), 2.0; Zn (ZnSO4), 34.4; Mn (MnSO4), 6.2; Fe (FeSO4), 21.1; I (Ca (IO3)2), 1.63; Se (Na2SeO3), 0.18; Co (CoCl2), 0.24; Mg (MgSO4·H2O), 52.71.2 试验对象及养殖
试验在上海海洋大学滨海养殖基地进行, 养殖对象为大口黑鲈。在正式试验前, 试验鱼在养殖系统中暂养 2 周, 期间投喂商品饲料。在试验开始时, 饥饿处理24h, 挑选体质健康、规格均匀的大口黑鲈(17.17±0.14) g, 每组设置4个网箱, 每个网箱35尾鱼, 养殖周期为67d, 每天限量投喂2次(8: 00和16: 00), 日投喂量为体重的2%—3%且保持组间相等。定期检测水质, 水质条件保持在: 溶氧浓度>5.0 mg/L, 总氨氮浓度<0.5 mg/L, pH7.5—8.5, 水温23—26℃, 光照时间为自然光照, 7d左右吸污换水1次, 换水量约为1/3。
1.3 样品采集
在试验开始前, 称量大口黑鲈的初始体重, 并挑选体重与试验用鱼接近的大口黑鲈 10 尾, 置于–20℃冰箱保存, 用于测定初始肌肉样本中的含氮量。在养殖试验结束后, 饥饿处理24h, 分别统计每个平行大口黑鲈的数量以及体重。每平行随机捞取10尾鱼, 使用丁香酚麻醉。其中3尾–20℃冰箱保存, 用作全鱼的体成分分析, 剩余7尾分别称量其体重, 测量体长、全长, 之后进行尾静脉取血, 置于用肝素钠抗凝剂润洗的离心管中, 并以3500 r/min离心15min后取上清液即为血清, 置于–20℃冰箱保存, 用于血清生理生化指标分析。取血完毕后进行鱼体解剖, 称量内脏团重、肝脏重, 用于计算鱼体的形体指标。分别取其肝脏、胃、肠道、幽门盲囊置于–20℃冰箱, 用于肝脏中粗脂肪含量的测定以及消化酶活力测定。取侧线以上肌肉, 冷冻干燥至恒重后, 粉碎密封保存, 用于测定肌肉样本中粗蛋白、粗脂肪的含量。
1.4 测定指标与方法
营养常规的测定 试验饲料以及鱼体水分测定采用105℃烘箱干燥恒重法(GB/T6435-1986), 粗蛋白质含量测定采用凯氏定氮法(GB/T6432-1994), 粗脂肪含量采用氯仿-甲醇法测定, 粗灰分含量采用马弗炉550℃灼烧法(GB/T 6438-1992)。
消化酶活性的测定 蛋白酶活力单位定义: 每毫克组织蛋白一定温度和pH条件下, 1min水解酪蛋白产生1 μg酪氨酸, 即为1个酶活力单位, 以U/mg prot表示。
淀粉酶活力单位定义: 组织中1 mg蛋白在37℃与底物作用30min, 水解10 mg淀粉定义为1个淀粉酶活力单位, 以U/mg prot表示。
组织脂肪酶单位定义: 在37℃条件下, 每毫克组织蛋白在本反应体系中与底物反应1min每消耗1 μmol底物为1个酶活力单位, 以U/mg prot表示。
淀粉酶、脂肪酶活力均采用南京建成生物工程研究所提供的试剂盒进行测定, 蛋白酶活力使用福林法测定(GB/T23527-2009), 组织匀浆液的蛋白质浓度采用考马斯亮蓝染色法进行测定。
肝脏功能测定 过氧化氢酶 (CAT)、总抗氧化能力 (T-AOC)、丙二醛(MDA)、总超氧化歧化酶 (T-SOD)、谷草转氨酶(AST)和谷丙转氨酶(ALT)均采用南京建成生物工程研究所提供的试剂盒进行测定, 肝脏匀浆液的蛋白质浓度采用考马斯亮蓝染色法进行测定。
耗氧率、排氨率的测定 采用静水密闭式呼吸实验方法[13, 14]。将体积为30 L的满水水桶进行曝气, 在养殖试验结束并饥饿处理24h后, 每组随机捞取16尾大口黑鲈, 平均放入4个曝气的水桶中, 并以不放入鱼的满水水桶作为对照组。在试验前, 先让试验鱼在水桶中适应2—3h, 待呼吸平稳后再进行试验。在试验进行过程中, 桶口密封, 温度保持在25℃左右, 持续进行2h。在试验结束后, 分别用溶氧仪、纳氏试剂比色法测定水体中溶解氧含量和氨氮含量, 并用分析天平称量鱼体湿重(精确至0.001 g)。在试验开始时, 水中的溶解氧为4.9—6.4 mg/L, 氨氮浓度为0.2—0.5 mg/L; 在试验结束时, 水中的溶解氧范围为0.9—3.5 mg/L, 氨氮浓度为0.7—1.8 mg/L。根据以下公式计算耗氧率以及排氨率。
耗氧率=(C0–C1)×V/(W×T)
式中: C0为试验结束时对照组水中的溶解氧浓度(mg/L); C1为试验结束时试验组水中的溶解氧浓度(mg/L); V为水样的体积(L); W为试验鱼的体重(g); T为试验持续的时间(h)。
排氨率=(N1–N0)×V/(W×T)
式中: N0为试验结束时对照组水样中氨氮的浓度(mg/L); N1为试验结束时试验组水样中氨氮的浓度(mg/L); V为水样的体积(L); W为试验鱼的体重(g); T为试验持续的时间(h)。
脂质代谢指标的测定 血清游离脂肪酸(NEFA)、甘油三酯(TG)、总胆固醇(T-CHO)、低密度脂蛋白胆固醇(LDL-C)含量的测定所使用的试剂盒购于南京建成生物工程研究所, 按试剂盒说明书测定脂质代谢指标。
分别取0.5 g冷冻干燥后的肝脏、肌肉样品, 使用氯仿-甲醇提取法测定肝脏、肌肉中的粗脂肪含量。
1.5 指标的计算
成活率(Survival Rate, SR, %)=100%×试验结束鱼尾数/试验初始鱼尾数;
增重率(Weight Gain Rate, WGR, %)=100%×(鱼末均重–鱼初均重)/初体均重;
特定生长率(Specific Growth Rate, SGR, %/d)=100%×(ln鱼末均重–ln鱼初均重)/试验天数;
饲料系数(Feed Coefficient Ratio, FCR)=总投饵量/(终末总重–初始总重+死亡总重)
鱼体肥满度(Condition Factor, CF, g/cm3)=100 g/cm3×鱼末均重/体长3
脏体比(Viscerosomatic Index, VSI)=100×内脏重(g)/体重(g)
肝体比(Hepatosomatic index, HSI)=100%×肝脏重(g)/体重(g)
肌肉氮的沉积率(%)=100%×(终末鱼体重×终末肌肉干物质含量×终末肌肉氮的含量–初始鱼体重×初始肌肉干物质含量×初始肌肉氮的含量)/(饲料消耗量×饲料干物质含量×饲料中氮的含量)
1.6 数据统计与分析
采用SPSS Statistics17.0软件对数据进行单因素方差分析(One-way ANOVA), 若组间差异显著, 用Duncan氏法进行多重差异显著性比较, 显著水平P<0.05。试验数据以平均值±标准差(Mean ± SD)表示。
2. 结果
2.1 酶解豆粕替代鱼粉对大口黑鲈生长性能、饲料利用及体成分的影响
由表 3可知, 酶解豆粕替代鱼粉时, E0、E15和E20三组的增重率无显著性差异(P>0.05), E25组和E30组的增重率显著高于其他组(P<0.05), 其中增重率最高组为E30。FSBM、SBM和E20三组增重率呈依次增高趋势, 但无显著性组间差异(P>0.05)。随着酶解豆粕对鱼粉替代水平的升高, 特定生长率呈上升趋势, E25组和E30组的特定生长率显著高于其他组(P<0.05)。不同豆粕替代30.91%的鱼粉时, E20组的特定生长率显著高于FSBM组(P<0.05)。随着酶解豆粕对鱼粉替代量的增加, 存活率显著降低, E25组和E30组存活率显著低于其他组(P<0.05), 分别为84.29%和86.43%。E20组的存活率显著低于FSBM组(P<0.05)。在酶解豆粕替代鱼粉时, E20和E30组的饲料系数最低, 但是各组之间没有显著性差异(P>0.05)。不同豆粕替代30.91%鱼粉对大口黑鲈的饲料系数无显著影响(P>0.05)。
表 3 酶解豆粕替代鱼粉对大口黑鲈生长、存活率及饲料系数的影响Table 3. Effects of enzymatic hydrolysis of soybean meal on growth, survival rate and feed coefficient ratio of largemouth bass (n=4)组别Group 初始均重Initial weight (g) 终末均重Final weight (g) 增重率WGR
(%)特定生长率 SGR
(%/d)存活率SR
(%)饲料系数FCR E0 17.23±0.16 73.32±0.84abc 325.58±2.83ab 2.13±0.01ab 100.00±0.00c 0.87±0.01ab E15 17.09±0.12 73.40±2.46abc 334.70±11.04b 2.16±0.04b 95.00±4.29bc 0.89±0.02ab E20 17.21±0.12 75.14±1.06bc 336.14±8.12b 2.17±0.03b 88.57±7.74ab 0.84±0.06a E25 17.13±0.13 76.79±2.83c 355.72±6.99c 2.23±0.02c 84.29±4.95a 0.87±0.03ab E30 17.20±0.12 76.08±3.36bc 362.42±19.34c 2.22±0.02c 86.43±6.34a 0.85±0.04a SBM 17.16±0.21 73.21±1.02ab 326.56±8.74ab 2.13±0.03ab 95.00±1.43bc 0.89±0.01ab FSBM 17.19±0.15 70.91±1.51a 312.60±8.31a 2.10±0.01a 98.57±1.65c 0.91±0.02b 注: 同列无字母或数据肩标相同字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05),下表同
Note: In the same column, values with no letter or the same letter superscripts mean no significant difference (P>0.05), while different small letter superscripts mean significant differences (P<0.05). The same applies below由表 4可知, 对照组E0的肥满度、脏体比显著高于其他组(P<0.05), 发酵豆粕组(FSBM)的脏体比显著高于E20组。随着酶解豆粕对鱼粉替代水平的升高, 各组的肝体比显著降低(P<0.05), 其中, 肝体比在E30组最低。
表 4 酶解豆粕替代鱼粉对大口黑鲈形体指标(n=28)及体成分(n=4)的影响Table 4. Effects of enzymatic hydrolysis of soybean meal on body index (n=28) and body composition (n=4) of largemouth bass (%)组别Group 肥满度CF (g/cm3) 脏体比VSI 肝体比HSI 水分Moisture 灰分Ash 粗蛋白Crude protein 粗脂肪Crude lipid E0 2.04±0.12c 6.91±0.55c 2.48±0.22f 69.78±0.39a 15.68±0.24d 58.55±0.67a 21.21±0.14d E15 1.98±0.15ab 5.73±0.43a 1.19±0.11d 70.72±0.22cd 14.99±0.51bc 60.55±0.01b 19.71±0.56c E20 1.95±0.11ab 5.54±0.48a 0.96±0.08b 69.90±0.22ab 13.95±0.12a 58.95±0.50a 20.87±0.41cd E25 2.00±0.14ab 5.78±0.48a 1.07±0.10c 71.06±0.22d 15.05±0.23bc 60.78±0.88b 17.33±1.02b E30 1.99±0.13ab 5.56±0.46a 0.81±0.09a 70.84±0.27cd 14.89±0.14b 61.08±0.62b 16.10±0.82a SBM 1.91±0.09a 5.60±0.48a 1.01±0.07b 70.34±0.67bc 15.41±0.15c 59.17±0.82a 17.86±0.26b FSBM 1.91±0.11a 6.13±0.49b 1.55±0.13e 70.80±0.23cd 15.12±0.35bc 58.16±0.84a 17.83±1.06b 当酶解豆粕替代鱼粉时, 除E20组外, 各替代组的全鱼水分、粗蛋白含量均显著高于对照组(E0组)(P<0.05), E20组的全鱼灰分含量显著低于其他组(P<0.05), 脂肪含量随着替代水平的增加显著降低(P<0.05)。
不同豆粕替代30.91%的鱼粉时, E20组的全鱼水分含量显著高于FSBM组(P<0.05), 全鱼灰分含量显著低于SBM和FSBM组(P<0.05), 粗脂肪含量则相反, 即显著高于其他两组(P<0.05), 三组的全鱼粗蛋白含量无显著差异(P>0.05)。
2.2 酶解豆粕替代鱼粉对大口黑鲈消化酶活性的影响
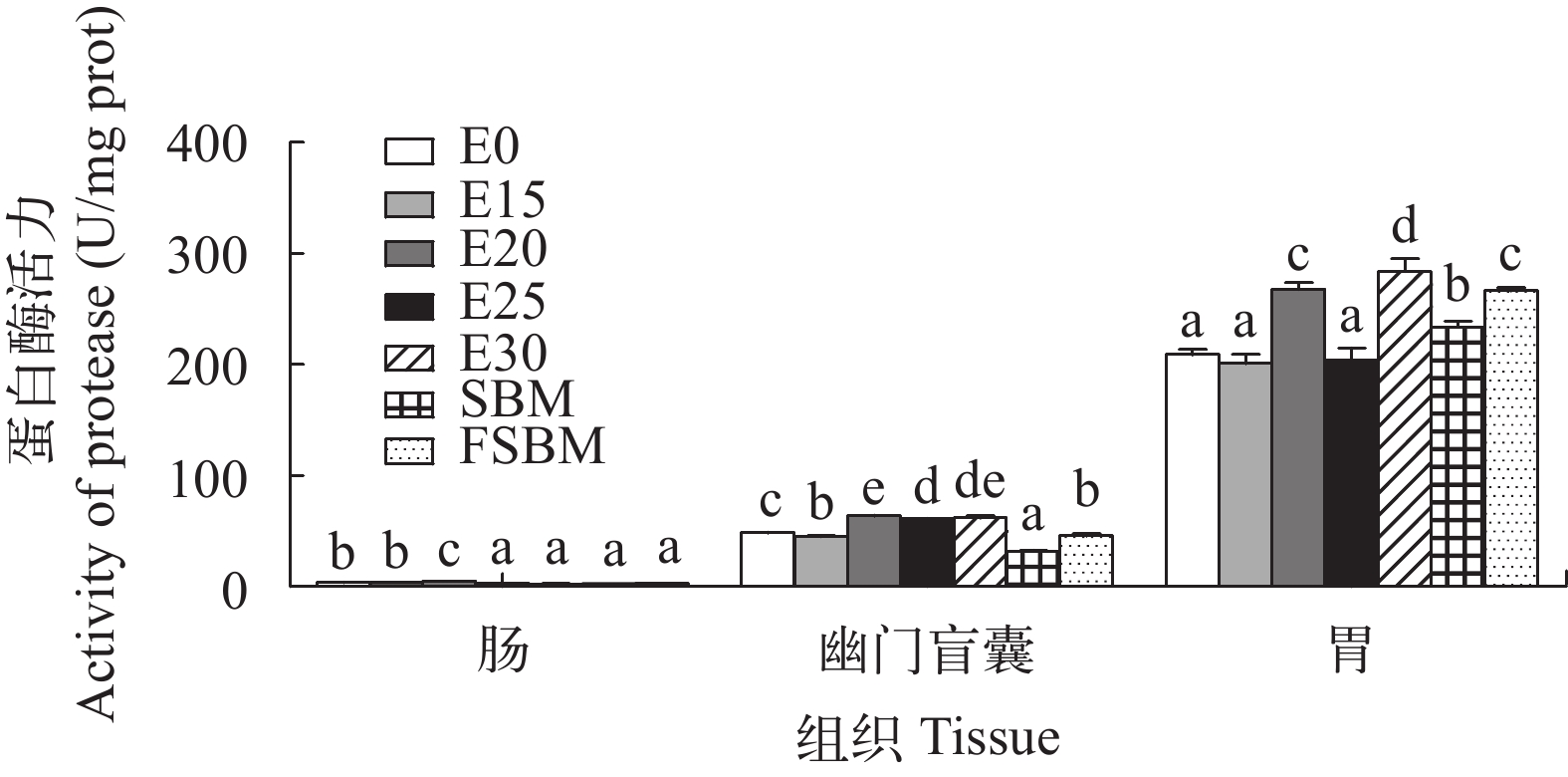
由图 2可知, 随着酶解豆粕添加量的增加, 替代组肠道的蛋白酶活力先上升后下降, 在E20组 达到最大值; 幽门盲囊的蛋白酶活力呈上升趋势; E20和E30两组的胃蛋白酶活力显著高于其他组(P<0.05)。在3种豆粕替代效果对比中, SBM组的幽门盲囊、胃蛋白酶的活力显著低于其他两组(P<0.05)。
![]() 图 2 酶解豆粕替代鱼粉对大口黑鲈蛋白酶活力的影响图中无字母或相同字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05); 下图同Figure 2. Effects of enzymatic hydrolyzed soybean meal on protease activity of largemouth bass (n=4)Values with the same superscripts or none superscripts mean no significant difference (P>0.05), while different superscripts mean significant difference (P<0.05). The same applies below
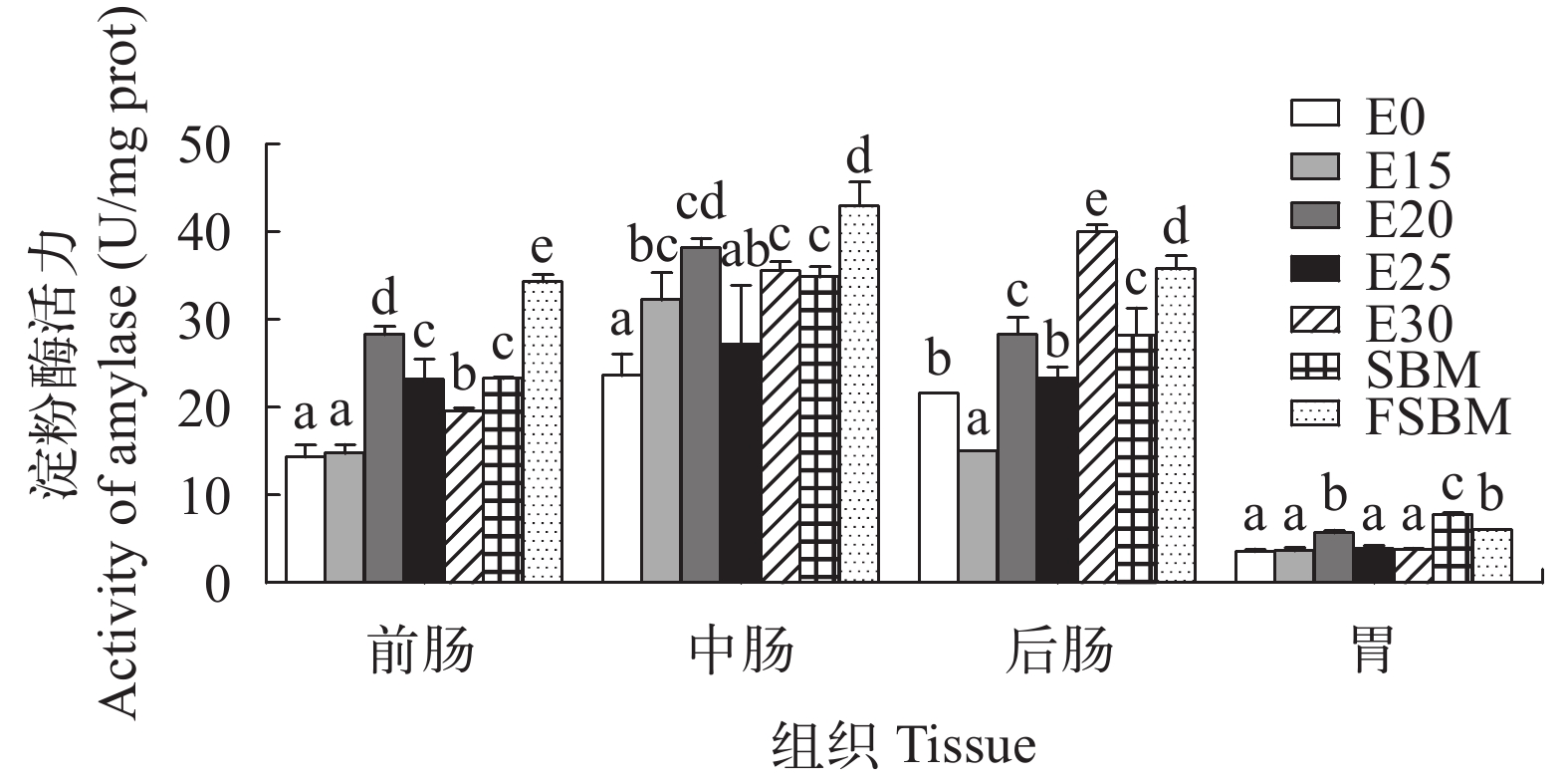
图 2 酶解豆粕替代鱼粉对大口黑鲈蛋白酶活力的影响图中无字母或相同字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05); 下图同Figure 2. Effects of enzymatic hydrolyzed soybean meal on protease activity of largemouth bass (n=4)Values with the same superscripts or none superscripts mean no significant difference (P>0.05), while different superscripts mean significant difference (P<0.05). The same applies below由图 3可知, 在酶解豆粕替代鱼粉时, 前肠淀粉酶呈先上升后下降的趋势, 但是替代组显著高于对照组(P<0.05); 中肠淀粉酶活力除E25组和对照组无显著性差异外(P>0.05), 其他替代组均显著高于对照组(P<0.05); 后肠E15组的淀粉酶活力显著低于对照组(P<0.05), E20、E30组显著高于对照组(P<0.05)。在同一替代水平下, FSBM组的肠道淀粉酶活力显著高于其他组(P<0.05)。
![]() 图 3 酶解豆粕替代鱼粉对大口黑鲈淀粉酶活力的影响Figure 3. Effects of enzymatic hydrolyzed soybean meal on amylase activity of largemouth bass (n=4)
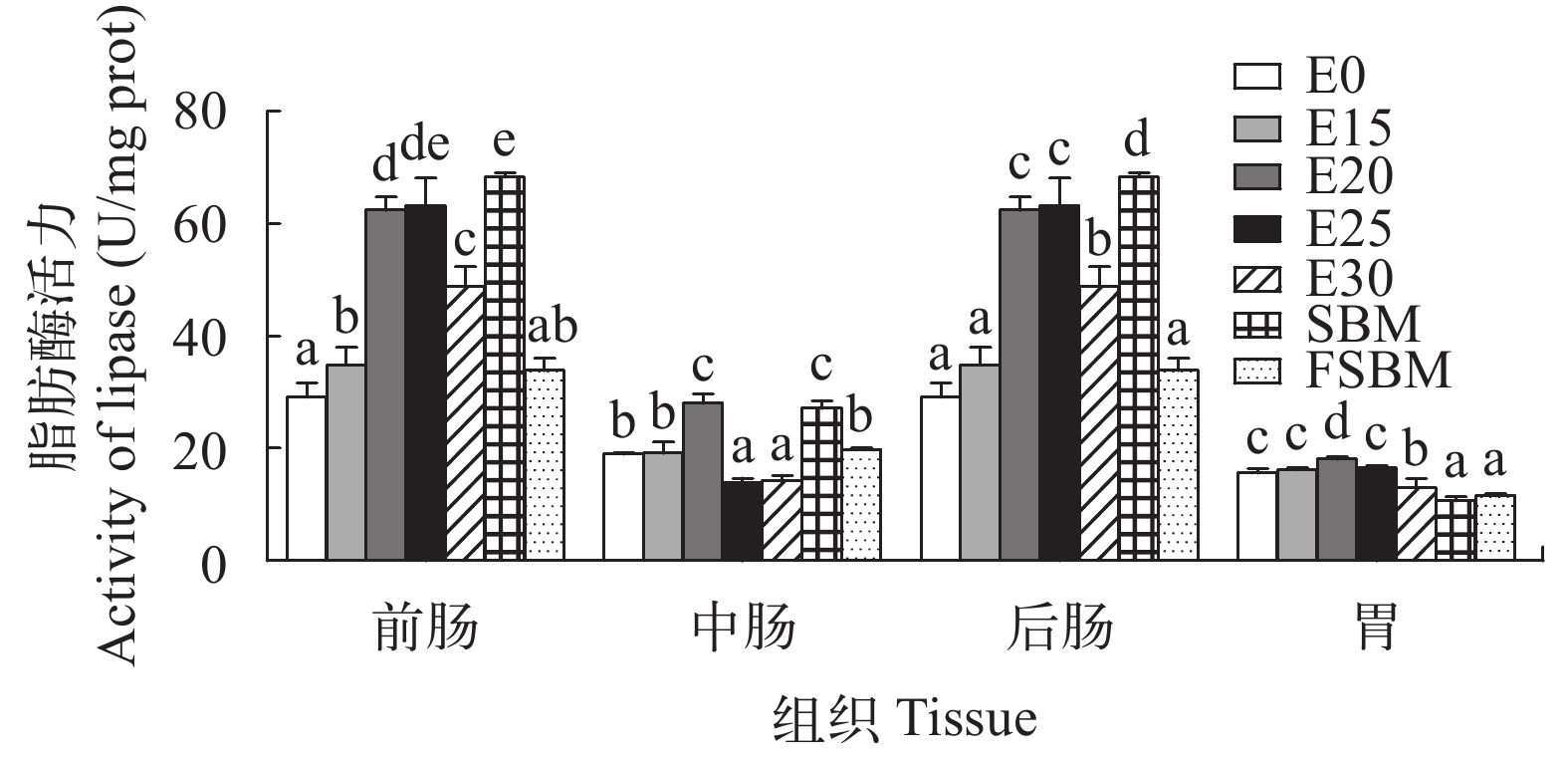
图 3 酶解豆粕替代鱼粉对大口黑鲈淀粉酶活力的影响Figure 3. Effects of enzymatic hydrolyzed soybean meal on amylase activity of largemouth bass (n=4)由图 4可知, 随着酶解豆粕替代水平的增加, 前肠、后肠的脂肪酶活力先上升后下降, 但是显著高于E0组(P<0.05); 中肠脂肪酶活力呈相同趋势, E20组达到最大值, E25和E30组显著低于其他组(P<0.05)。在不同豆粕替代鱼粉时, FSBM的肠道脂肪酶活力显著低于其他两组(P<0.05)。
![]() 图 4 酶解豆粕替代鱼粉对大口黑鲈脂肪酶活力的影响Figure 4. Effects of enzymatic hydrolyzed soybean meal on lipase activity of largemouth bass (n=4)
图 4 酶解豆粕替代鱼粉对大口黑鲈脂肪酶活力的影响Figure 4. Effects of enzymatic hydrolyzed soybean meal on lipase activity of largemouth bass (n=4)2.3 酶解豆粕替代鱼粉对大口黑鲈肝脏功能的影响
由表 5可知, 在酶解豆粕替代鱼粉时, 肝脏的过氧化氢酶(CAT)、总超氧化歧化酶(T-SOD)呈先下降后上升的趋势, 但都显著低于基础组(P<0.05); 肝脏的总抗氧化能力(T-AOC)显著高于对照组(P<0.05), 在E25组达到最大值; 丙二醛(MDA)均显著低于对照组, 且在E25组达到最小值; 肝脏谷草转氨酶(AST)、谷丙转氨酶(ALT)呈先上升后下降, 除E30组的ALT与对照组无显著性差异外, 其余替代组均显著高于对照组(P<0.05)。
表 5 酶解豆粕替代鱼粉对大口黑鲈肝脏功能的影响Table 5. Effects of enzymatic hydrolysis of soybean meal on liver function of largemouth bass (n=4)组别Group 过氧化氢酶CAT (U/mg prot) 总抗氧化能力T-AOC (U/mg prot) 丙二醛MDA (nmol/mg prot) 总超氧化歧化酶T-SOD (U/mg prot) 谷草转氨酶AST (U/g prot) 谷丙转氨酶ALT (U/g prot) E0 49.82±0.96f 0.37±0.02a 4.33±0.54e 506.47±16.82f 0.87±0.05a 4.77±0.18b E15 41.44±1.05e 0.61±0.02cd 1.28±0.51bc 398.01±12.31d 1.78±0.05c 5.49±0.28c E20 30.46±1.60c 0.57±0.01c 3.17±0.30d 319.79±7.44a 2.74±0.20f 5.44±0.41c E25 35.95±0.63d 0.86±0.03e 0.73±0.03ab 340.46±8.33b 2.14±0.13d 6.16±0.22d E30 30.66±0.16c 0.65±0.02d 1.32±0.06c 345.18±5.49b 1.77±0.03c 4.39±0.20ab SBM 27.56±0.87b 0.48±0.02b 1.56±0.17c 371.55±6.28c 0.99±0.01a 5.55±0.12c FSBM 23.95±3.59a 0.65±0.05d 0.68±0.05a 453.83±23.75e 1.41±0.03b 4.28±0.08a 在替代30.91%鱼粉时, FSBM组的肝脏CAT和MDA显著低于其他两组(P<0.05); 肝脏T-AOC活性依次为SBM<E20<FSBM, 且三组之间有显著性差异(P<0.05); FSBM组的肝脏T-SOD显著高于其他两组(P<0.05); E20组的AST显著高于其他两组(P<0.05), ALT显著高于FSBM组(P<0.05)。
2.4 酶解豆粕替代鱼粉对大口黑鲈代谢的影响
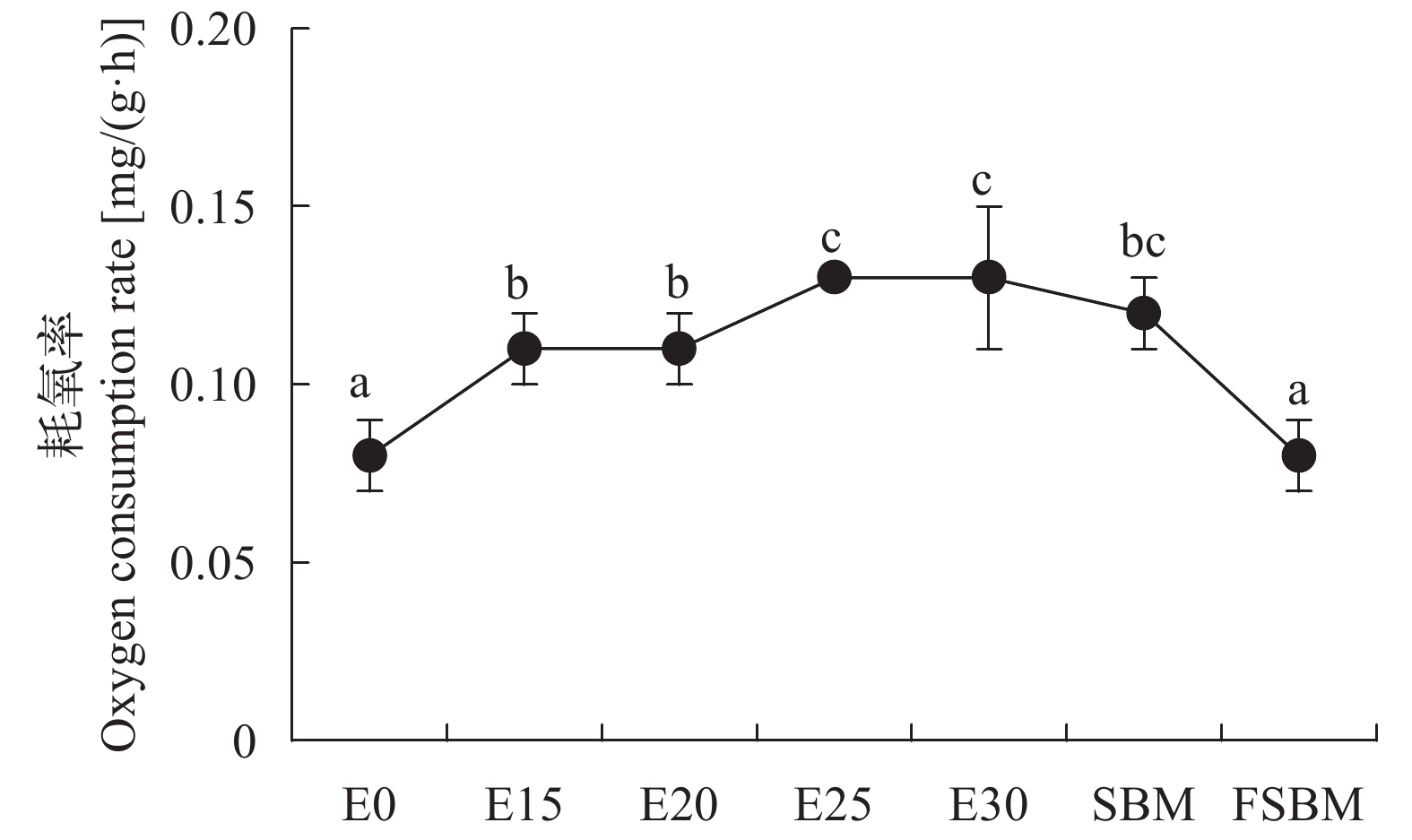
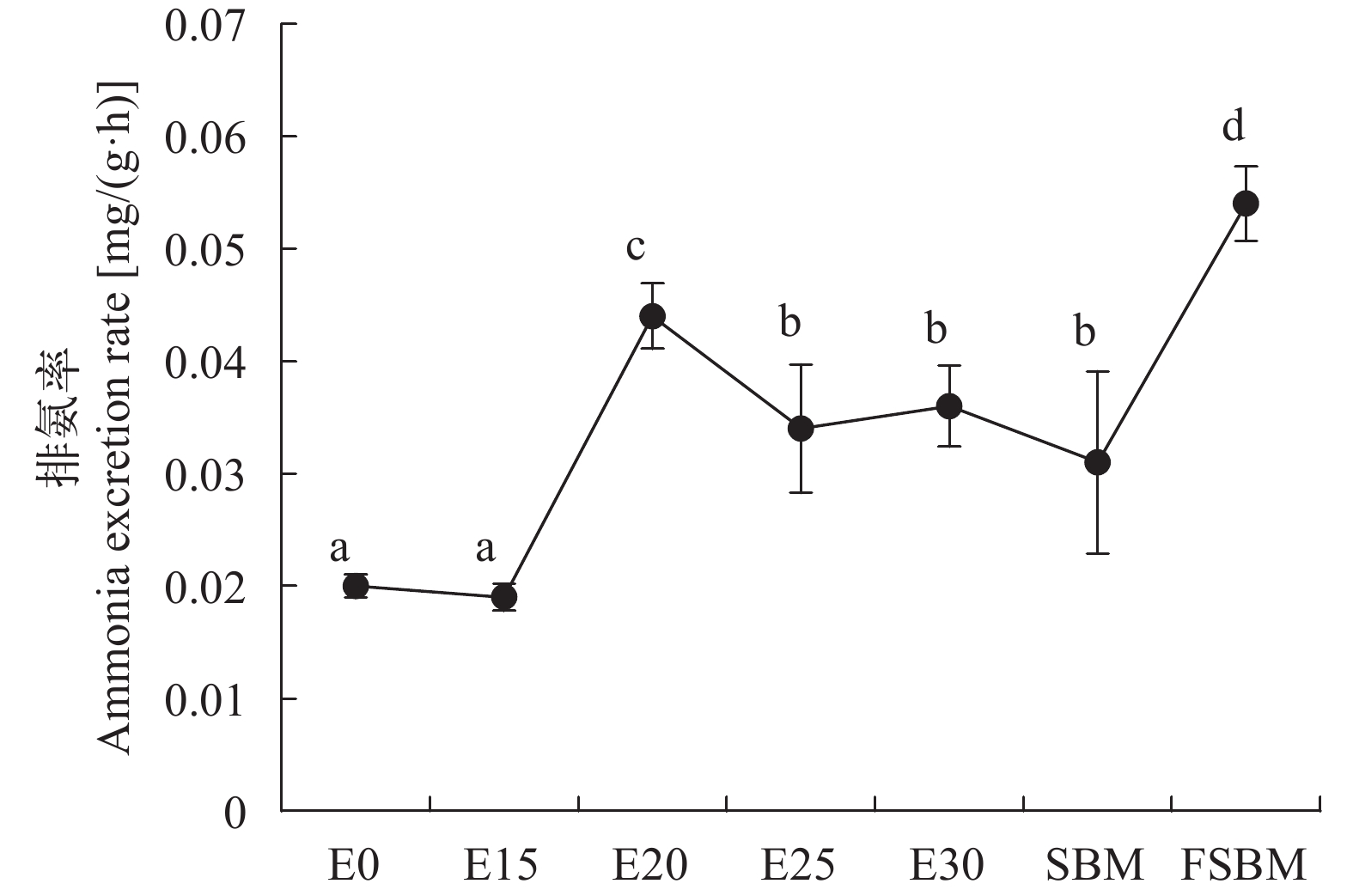
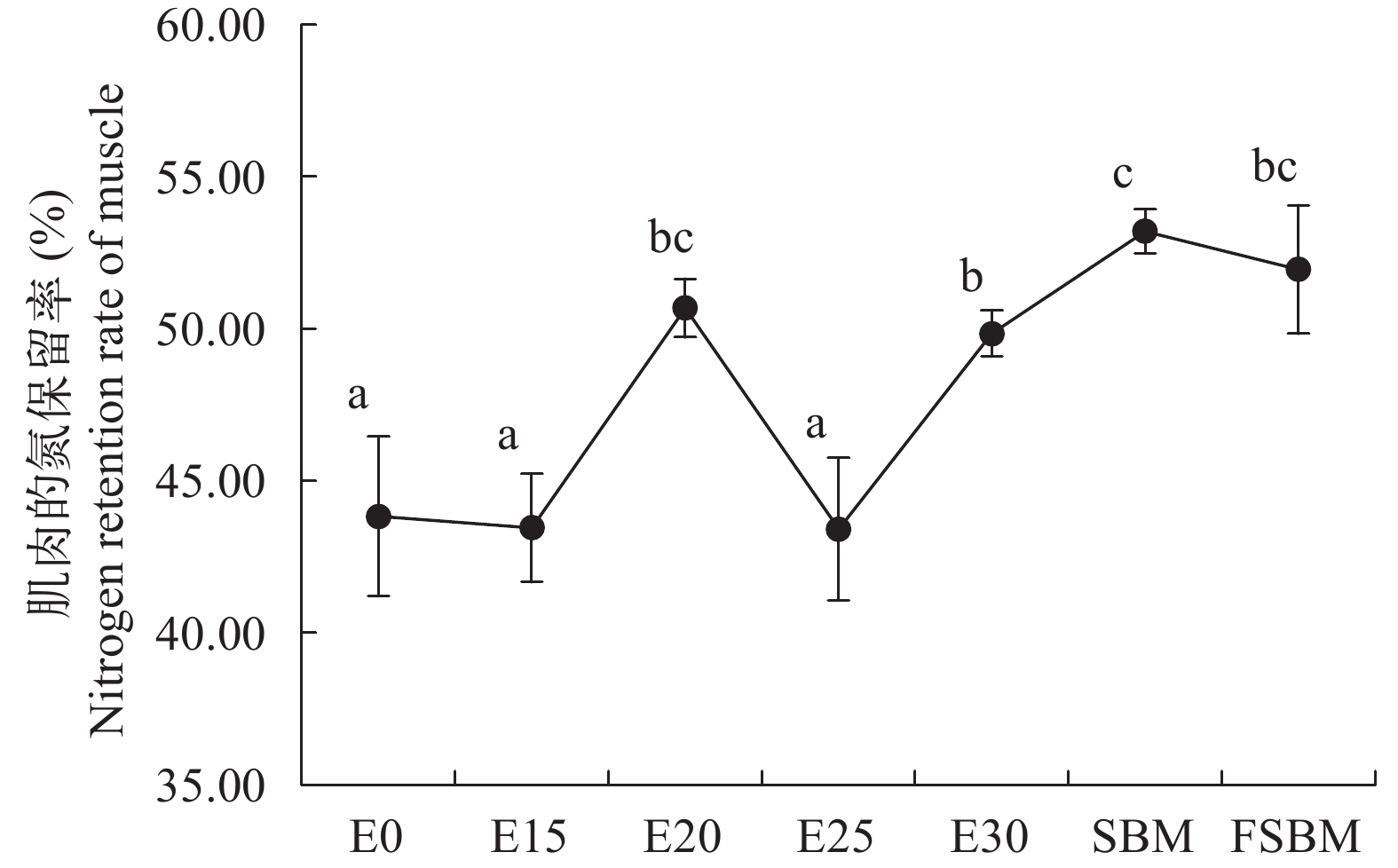
酶解豆粕替代鱼粉对大口黑鲈耗氧率、排氨率的影响 由图 5可知, 随着酶解豆粕替代水平增加, 耗氧率显著增高(P<0.05); 不同豆粕替代30.91%鱼粉时, FSBM组的耗氧率显著低于其他两组(P<0.05)。由图 6可知, 在酶解豆粕替代鱼粉时, E20、E25和E30组的排氨率显著高于对照组(P<0.05), 其中E20组值最高, E15组与对照组无显著性差异(P>0.05); 在不同豆粕替代30.91%鱼粉时, FSBM组的排氨率显著高于其他两组(P<0.05)。由图 7可知, 在酶解豆粕不同水平替代鱼粉时, E20和E30两组的氮的保留率显著高于其他组(P>0.05); 在不同豆粕替代30.91%鱼粉时, 酶解豆粕组、豆粕组和发酵豆粕组的氮的保留率无显著性差异(P>0.05)。
![]() 图 5 酶解豆粕替代鱼粉对大口黑鲈的耗氧率的影响Figure 5. Effects of enzymatic hydrolysis of soybean meal on oxygen consumption rate of largemouth bass (n=4)
图 5 酶解豆粕替代鱼粉对大口黑鲈的耗氧率的影响Figure 5. Effects of enzymatic hydrolysis of soybean meal on oxygen consumption rate of largemouth bass (n=4)![]() 图 6 酶解豆粕替代鱼粉对大口黑鲈的排氨率的影响Figure 6. Effects of enzymatic hydrolysis of soybean meal on ammonia excretion rate of largemouth bass (n=4)
图 6 酶解豆粕替代鱼粉对大口黑鲈的排氨率的影响Figure 6. Effects of enzymatic hydrolysis of soybean meal on ammonia excretion rate of largemouth bass (n=4)![]() 图 7 酶解豆粕替代鱼粉对大口黑鲈肌肉氮保留率的影响Figure 7. Effects of enzymatic hydrolysis of soybean meal on muscle nitrogen retention rate of largemouth bass (n=4)
图 7 酶解豆粕替代鱼粉对大口黑鲈肌肉氮保留率的影响Figure 7. Effects of enzymatic hydrolysis of soybean meal on muscle nitrogen retention rate of largemouth bass (n=4)酶解豆粕替代鱼粉对大口黑鲈脂质代谢的影响 由表 6可知, 随着酶解豆粕替代量的增加, 血清游离脂肪酸(NEFA)、血清低密度脂蛋白胆固醇(LDL-C)和肌肉脂肪含量呈现先升高后降低的趋势; 血清总胆固醇(T-CHO)及肝脏的脂肪含量显著升高(P<0.05); E25组的血清甘油三酯(TG)显著高于其他组(P<0.05)。
表 6 酶解豆粕替代鱼粉对大口黑鲈脂质代谢指标的影响Table 6. Effects of enzymatic hydrolysis of soybean meal on lipid metabolism of largemouth bass (n=4)组别Group 血清游离脂肪酸Serum NEFA (μmol/L) 血清总胆固醇Serum T-CHO (mmol/L) 血清甘油三酯Serum TG
(mmol/L)血清低密度脂蛋白胆固醇Serum LDL-C (mmol/L) 肝脏脂肪含量Liver lipid (%) 肌肉脂肪含量Muscle lipid (%) E0 268.69±6.06a 4.01±0.32a 2.71±0.10a 1.39±0.05a 7.72±0.27a 3.86±0.17d E15 305.72±20.34b 4.66±0.31b 2.62±0.18a 1.77±0.04b 8.57±0.23b 4.50±0.16e E20 366.33±27.50cd 4.71±0.31b 2.87±0.17a 2.10±0.03c 10.82±0.06de 3.80±0.06cd E25 396.77±11.12d 5.28±0.06c 3.29±0.18b 2.40±0.07d 10.88±0.64de 3.50±0.16bc E30 354.21±24.35c 6.52±0.39d 2.79±0.14a 1.43±0.25a 11.47±0.70e 3.34±0.19ab SBM 369.02±16.33cd 5.60±0.28c 3.41±0.18b 1.76±0.05b 10.43±0.54d 3.13±0.27a FSBM 359.93±6.85c 4.32±0.06ab 3.37±0.25b 1.95±0.01c 9.47±0.22c 4.53±0.13e 当替代30.91%鱼粉时, 三组的血清游离脂肪酸(NEFA)无显著性变化(P>0.05); SBM组的血清总胆固醇(T-CHO)显著高于其他两组, 而血清低密度脂蛋白胆固醇(LDL-C)与肌肉脂肪含量呈相反的结果; E20组的血清TG显著低于其他两组; E20和15FSBM的肝脏脂肪含量显著高于SBM组。
3. 讨论
3.1 酶解豆粕替代鱼粉对大口黑鲈生长性能、饲料利用及体成分的影响
已有不少研究表明, 酶解大豆蛋白对水产动物的生长有促进作用。李学丽等[15]在珍珠龙胆石斑鱼(Epinephelus fuscoguttatus♀× Epinephelus lanceolatus ♂)幼鱼饲料中, 用酶解豆粕替代40%的鱼粉时珍珠龙胆石斑鱼幼鱼的增重率和特定生长率都有显著的提高。Rep等[16]研究表明牙鲆(Paralichthys olivaceus)幼鱼饲料中酶解大豆多肽替代鱼粉的添加量为20%时促进试验鱼的生长。本研究结果表明, 当酶解豆粕添加量为25%—30%, 即替代40%—47.27%鱼粉时, 对大口黑鲈的生长有显著的促进作用。酶解豆粕促进水产动物的生长性能与其富含小肽有关。与游离氨基酸吸收相比, 小肽的转运和吸收具有速度快、耗能低、不易饱和等特点[17]。Newey等[18] 发现小肽能完整地被吸收并以二、三肽形式进入血液循环, 直接参与组织蛋白质的合成, 此外肝脏、肾脏、皮肤和其他组织也能完整地利用小肽。小肽与游离氨基酸具有相互独立的吸收机制, 二者互不干扰[19]。除了直接的营养作用外, 酶解豆粕中的大豆肽可能通过调节信号转导途径中的关键因子而有利于细胞生长[20]。但是, 本试验结果显示在E25和E30组的存活率较低, 可能是因为限量投喂造成摄食不均衡, 从而导致大口黑鲈生长出现差异, 小个体被大个体残杀所致, 这可以通过增大投喂量的办法加以弥补, 这也可能是造成E25 和E30组的生长性能显著高于其他组的原因之一。在酶解豆粕、豆粕、发酵豆粕替代相同水平的鱼粉时, 三组之间的增重率无显著性差异, 可能是因为在这一替代水平下, 小肽的有效吸收以及添加的晶体蛋氨酸、赖氨酸、牛磺酸对鱼体所需氨基酸的平衡, 有效改善了植物蛋白替代鱼粉时所带来的负面影响[21]。本试验结果显示, 在酶解豆粕替代鱼粉时, 大口黑鲈的脏体比和肝体比都有显著性的降低, 这说明在高鱼粉饲料中, 适当降低鱼粉含量, 可以改善肝脏状况[22]。
饲料给鱼类提供营养, 鱼类通过摄食饲料对其体成分产生影响。在本试验中, 酶解豆粕替代鱼粉时, 大口黑鲈全鱼的粗蛋白含量有所提高, 脂肪含量呈下降趋势。这是由于酶解豆粕中的小肽可以直接被吸收利用, 参与蛋白质合成, 从而促进蛋白质的沉积[23]。在星斑川鲽的研究中, 全鱼粗脂肪含量有相似的变化[8]。研究发现, 大豆肽既能刺激交感神经, 阻止脂肪的吸收和促进脂质的分解代谢, 也能刺激产生热能的褐色脂肪组织(BAT)的活性, 提高甲状腺素在血液中的浓度, 从而提高基础代谢水平, 促进体内多余脂肪的消耗[24]。
3.2 酶解豆粕替代鱼粉对大口黑鲈消化酶活性的影响
不少研究表明, 水产动物饲料中大豆蛋白源替代鱼粉会对鱼体的消化酶活性产生影响。Lin等[25]研究发现, 当豆粕替代鱼粉比例大于 75% 时, 奥尼罗非鱼(Oreochromis niloticus×O. aureus)幼鱼肠道蛋白酶活性显著低于对照组。钱曦等[26]对翘嘴红鲌(Erythroculter Ilishaeformis Bleeker)饲料的研究表明, 当豆粕对鱼粉的替代比例达到 40.5% 和 54% 时, 肠道和肝胰脏的蛋白酶活力显著低于对照组。Xu等[27]的研究结果显示, 当饲料中大豆分离蛋白对鱼粉的替代水平为62.50%或更高, 中华鲟(Acipenser schrenckii)幼鱼前肠、中肠和后肠的脂肪酶和淀粉酶活性显著降低。本试验表明酶解豆粕替代组的前中后肠的淀粉酶活力相较于对照组都有不同程度的提高(E20组后肠除外), 替代组的脂肪酶活力均高于对照组(E25和E30组中肠除外), 这与关胜军等[28]研究一致。研究表明食物中的蛋白质主要在大口黑鲈的胃中消化, 很少在肠道中消化[29]。在本试验中, E20和E30的胃蛋白酶活力显著高于其他组, 说明这两组对蛋白质的消化效率较高。本试验结果与Zhao等[30]在黄颡鱼(Pelteobagrus fulvidraco)饲料中添加不同比例大豆肽替代鱼粉的研究结果相似。可能的原因是酶解豆粕中的一些生理活性小肽可直接作为神经递质间接刺激肠道激素受体或促进酶的分泌而发挥生理调节作用[31], 同时, 酶解蛋白肽不仅是诱导消化酶分泌的最适底物, 又能给机体消化酶的快速合成提供完整的氮架[32]。因此, 在饲料中添加小肽类物质能显著提高胃蛋白酶、脂肪酶、淀粉酶的活性, 促进蛋白质的合成, 防止肠黏膜萎缩和维持肠黏膜的正常结构和功能[33]。
3.3 酶解豆粕替代鱼粉对大口黑鲈肝脏抗氧化指标的影响
总抗氧化能力(T-AOC)的强弱与健康程度密切相关, 该防御体系由酶促和非酶促2个体系组成, 协同防护机体氧化; 丙二醛(MDA)是机体内自由基作用于脂肪发生过氧化反应的氧化终产物, 具有细胞毒性, 其含量间接反映了机体细胞中氧自由基含量以及受自, 由基攻击的严重程度[34]; 总超氧化歧化酶(T-SOD)是机体内天然存在的超氧自由基清除因子, 它可以把有害的超氧阴离子自由基转化为过氧化氢, 而过氧化氢酶(CAT)可以将过氧化氢分解成水和氧气, 以此来减轻活性氧自由基对机体造成的应激。刘文斌[35]的研究表明, 在饲料中添加植物蛋白酶解物对异育银鲫(Alogymogenetic crucian)的免疫能力有促进作用。本试验结果表明, 随着酶解豆粕不同水平替代鱼粉, 肝脏的T-AOC呈显著上升趋势, MDA呈相反趋势, 说明酶解豆粕有助于降低肝脏脂肪氧化, 进而保护肝脏健康。邓成萍等[36]研究表明大豆多肽具有很强的DPPH自由基清除能力, 且大多数具有抗氧化性的功能性肽主要集中在分子量小于 5000 Da的小肽中。由表 1可以看出本研究所用的酶解豆粕中89.79%的水溶性氮分子量<5000 Da, 为其抗氧化功能的发挥奠定了良好的分子基础。
谷 草 转 氨 酶(AST/GOT) 和谷丙 转 氨 酶 (ALT/GPT)是动物体内蛋白质代谢过程中的两种关键的代谢酶, ALT主要参与谷氨酸和丙酮酸间的转氨作用, 而AST则主要与谷氨酸和草酰乙酸之间的转氨作用相关。2种酶皆存在于鱼的肝脏中, 其活性反映了机体对蛋白质合成和分解代谢的状况[37]。在本试验中, 各替代组的AST活性显著高于对照组, 说明酶解豆粕替代鱼粉在一定程度上促进了机体的蛋白质合成代谢, 降低分解代谢。肝脏ALT活性的升高, 说明了机体尿素生成加快, 减少了氨基酸代谢废物对机体的毒害[38]。
3.4 酶解豆粕替代鱼粉对大口黑鲈代谢的影响
呼吸和排泄是鱼类进行能量代谢的基本生理活动, 耗氧率和排氨率是衡量鱼体能量消耗和代谢水平的重要指标。在本试验中, 随着酶解豆粕体替代量的增加, 大口黑鲈的耗氧率显著上升, 说明酶解豆粕的增加使其体内的生理活动增强, 代谢率提高, 可能是因为蛋白质作为能量的消耗增加, 也可能是由于脂肪和糖类作为能量消耗的量增加。
氨氮是水生动物的主要代谢产物之一, 是氨基酸脱氨基作用后的终产物, 排氨率可用来评估水生动物机体内氨基酸分解代谢的强弱以及利用蛋白质供能的情况[39]。一般来说, 随着饲料中植物蛋白源添加比例升高, 水生动物排氨率呈现逐渐升高的趋势, 这在欧洲舌齿鲈(Dicentrarchus labrax)[40]、虹鳟(Oncorhynchus mykiss)[41]、金头鲷(Sparus aurata)[42]等的研究结果中均有体现。本试验的结果显示, 酶解豆粕添加量高于20%时, 排氨率显著高于对照组, 与上述研究结果一致。代谢底物中氮排泄物的增加表示蛋白质作为能源物质消耗的比例增大[43]。除此之外, E20和E30两组肌肉的氮保留率显著高于其他组。在添加晶体蛋氨酸和赖氨酸保证饲料氨基酸平衡的前提下, 鱼体内的蛋白质合成速率和分解速率均提高, 使合成与分解差值增大, 促进生长, 即氮保留率增大[44]。当排氨率、肌肉氮的保留率同时升高, 间接反映出被消化吸收的蛋白质升高。
血清中TG水平是肝脏脂肪代谢的重要指示指标[45, 46], 是机体重要的供能物质, 也是机体主要的储能物质, 其水平的变化通常与T-CHO水平的变化趋势一致; 低密度脂蛋白(LDL)则携带胆固醇形成低密度脂蛋白携带胆固醇(LDL-C), 由肝脏向血液运输。在本试验中, 血清T-CHO和LDL-L含量随着酶解豆粕替代水平的增加而显著升高, 血清中TG也有上升的趋势, 说明随着酶解豆粕添加量的增加, 大口黑鲈的脂肪代谢加强, 机体转运清除外源性脂肪的能力增强, 更多的LDL-C从肝脏运往血液中, 缓解肝脏负担。同时, 随着酶解豆粕替代水平的增加, 肝体比显著降低, 这都说明了减少鱼粉使用量可以降低鱼粉氧化对肝脏的负作用[22]。鱼体内的脂肪代谢是一个复杂的过程, 其中包括脂肪的合成、氧化、代谢和转运。饲料中蛋白源的改变对鱼体脂肪代谢影响的分子机制还需进一步探究。
4. 结论
在本试验条件下, 大口黑鲈幼鱼饲料中酶解豆粕用量不超过30%, 即替代鱼粉的量不超过47.27%时, 对其生长无不利影响, 并且有利于减轻大口黑鲈肝脏负担, 提高肝脏的抗氧化功能及机体代谢水平。在添加晶体蛋氨酸、赖氨酸和牛磺酸满足鱼体所需氨基酸平衡的条件下, 普通豆粕、酶解豆粕与发酵豆粕均可以替代30.91%鱼粉, 且酶解豆粕的替代效果最好。
-
![]()
图 2 酶解豆粕替代鱼粉对大口黑鲈蛋白酶活力的影响
图中无字母或相同字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05); 下图同
Figure 2. Effects of enzymatic hydrolyzed soybean meal on protease activity of largemouth bass (n=4)
Values with the same superscripts or none superscripts mean no significant difference (P>0.05), while different superscripts mean significant difference (P<0.05). The same applies below
![]()
图 3 酶解豆粕替代鱼粉对大口黑鲈淀粉酶活力的影响
Figure 3. Effects of enzymatic hydrolyzed soybean meal on amylase activity of largemouth bass (n=4)
![]()
图 4 酶解豆粕替代鱼粉对大口黑鲈脂肪酶活力的影响
Figure 4. Effects of enzymatic hydrolyzed soybean meal on lipase activity of largemouth bass (n=4)
![]()
图 5 酶解豆粕替代鱼粉对大口黑鲈的耗氧率的影响
Figure 5. Effects of enzymatic hydrolysis of soybean meal on oxygen consumption rate of largemouth bass (n=4)
![]()
图 6 酶解豆粕替代鱼粉对大口黑鲈的排氨率的影响
Figure 6. Effects of enzymatic hydrolysis of soybean meal on ammonia excretion rate of largemouth bass (n=4)
![]()
图 7 酶解豆粕替代鱼粉对大口黑鲈肌肉氮保留率的影响
Figure 7. Effects of enzymatic hydrolysis of soybean meal on muscle nitrogen retention rate of largemouth bass (n=4)
表 1 酶解豆粕中水溶性氮的分子量分布
Table 1 Molecular mass distribution of water-soluble nitrogen in enzymatic hydrolyzed soybean meal
分子量范围Molecular mass 百分比Percentage (%) >5000 10.21 5000—3000 4.43 3000—2000 4.22 2000—1000 8.32 1000—500 11.49 500—180 16.63 <180 44.71  下载: 导出CSV
下载: 导出CSV
表 2 饲料组成及营养水平(%干物质基础)
Table 2 Composition and nutrient levels of diets (% DM basis)
指标Index 组别Group E0 E15 E20 E25 E30 SBM FSBM 鱼粉Fish meal 55 42 38 33 29 38 38 酶解豆粕Enzymatic hydrolyzed soybean meal 0 15 20 25 30 0 0 玉米蛋白粉Corn powder 3 4.5 5 5.5 6 5 5 豆粕Soybean meal 9 9 9 9 9 29.9 9 发酵豆粕Fermented soybean meal 0 0 0 0 0 0 19 血粉Dried blood 4 4 4 4 4 4 4 生粉Cornstarch 9 9 9 9 9 9 9 鱼油Fish oil 3 4 4.2 4.4 4.7 4.2 4.2 菜籽油Rap oil 3 3 3 3 3 3 3 多维Vitamins premix* 1 1 1 1 1 1 1 多矿Minerals premix** 0.5 0.5 0.5 0.5 0.5 0.5 0.5 磷酸二氢钙Ca(H2PO4)2 1 1 1 1 1 1 1 乌贼膏Squid paste 1 1 1 1 1 1 1 胆碱Choline 0.5 0.5 0.5 0.5 0.5 0.5 0.5 羧甲基纤维素CMC 6.67 3.54 2.25 1.69 0.41 1.65 2.35 沸石粉Zeolite powder 3.33 1.77 1.13 0.85 0.2 0.83 2.03 晶体蛋氨酸Crystal methionine 0 0 0.1 0.1 0.1 0.1 0.1 晶体赖氨酸Crystal lysine 0 0.1 0.2 0.3 0.4 0.2 0.2 牛磺酸Taurine 0 0.09 0.12 0.16 0.19 0.12 0.12 合计Total 100 100 100 100 100 100 100 营养水平Nutrient level 粗蛋白Crude protein (%) 46.74 46.48 46.62 46.08 46.22 46.63 46.12 粗脂肪Crude lipid (%) 11.03 11.22 11.18 11.06 11.11 11.20 11.46 总能Gross energy (MJ/kg) 18.70 19.13 19.25 19.29 19.45 19.21 18.96 替代鱼粉比例Replacement ratio of fish meal (%) 0 23.64 30.91 40 47.27 30.91 30.91 注: 总能计算公式[12]为: 总能(MJ/kg)=蛋白质×23.6+脂肪×39.5+无氮浸出物×17.6; *多维(IU或mg/kg干饲料): 维生素A, 16000 IU; 维生素D3, 800 IU; 维生素K3, 14.72; 维生素B1, 17.8; 维生素B2, 48; 维生素B6, 29.52; 维生素Bl2, 0.24; 维生素E, 160; 维生素C (35%), 800; 烟酸胺, 79.2; 泛酸钙, 73.6; 叶酸, 6.4; 生物素, 0.64; 肌醇, 320; 氯化胆碱, 1500; L-肉碱, 100; **多矿(mg/kg干饲料): 铜(CuSO4), 2.0; 锌(ZnSO4), 34.4; 锰(MnSO4), 6.2; 铁(FeSO4), 21.1; 碘(Ca(IO3)2), 1.63; 硒(Na2SeO3), 0.18; 钴(CoCl2), 0.24; 镁(MgSO4·H2O), 52.7
Note: Gross energy was calculated using the following formula: Gross energy (MJ/kg)= protein×23.6+ lipid×39.5+NFE×17.6; *Vitamin premix (IU or mg/kg dry diet): vitamin A, 16000 IU; vitamin D3, 8000 IU; vitamin K3, 14.72; thiamin, 17.8; ribo flavine, 48; pyridoxine, 29.52; cyanocobalamine, 0.24; tocopherol acetate, 160; ascorbic acid (35%), 800; niacinamide, 79.2; calcium-D-pantothenate, 73.6; folic acid, 6.4; biotin, 0.64; inositol, 320; choline chloride, 1500; L-carnitine, 100; **Mineral premix (mg/kg dry diet): Cu (CuSO4), 2.0; Zn (ZnSO4), 34.4; Mn (MnSO4), 6.2; Fe (FeSO4), 21.1; I (Ca (IO3)2), 1.63; Se (Na2SeO3), 0.18; Co (CoCl2), 0.24; Mg (MgSO4·H2O), 52.7
下载: 导出CSV
表 3 酶解豆粕替代鱼粉对大口黑鲈生长、存活率及饲料系数的影响
Table 3 Effects of enzymatic hydrolysis of soybean meal on growth, survival rate and feed coefficient ratio of largemouth bass (n=4)
组别Group 初始均重Initial weight (g) 终末均重Final weight (g) 增重率WGR
(%)特定生长率 SGR
(%/d)存活率SR
(%)饲料系数FCR E0 17.23±0.16 73.32±0.84abc 325.58±2.83ab 2.13±0.01ab 100.00±0.00c 0.87±0.01ab E15 17.09±0.12 73.40±2.46abc 334.70±11.04b 2.16±0.04b 95.00±4.29bc 0.89±0.02ab E20 17.21±0.12 75.14±1.06bc 336.14±8.12b 2.17±0.03b 88.57±7.74ab 0.84±0.06a E25 17.13±0.13 76.79±2.83c 355.72±6.99c 2.23±0.02c 84.29±4.95a 0.87±0.03ab E30 17.20±0.12 76.08±3.36bc 362.42±19.34c 2.22±0.02c 86.43±6.34a 0.85±0.04a SBM 17.16±0.21 73.21±1.02ab 326.56±8.74ab 2.13±0.03ab 95.00±1.43bc 0.89±0.01ab FSBM 17.19±0.15 70.91±1.51a 312.60±8.31a 2.10±0.01a 98.57±1.65c 0.91±0.02b 注: 同列无字母或数据肩标相同字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05),下表同
Note: In the same column, values with no letter or the same letter superscripts mean no significant difference (P>0.05), while different small letter superscripts mean significant differences (P<0.05). The same applies below
下载: 导出CSV
表 4 酶解豆粕替代鱼粉对大口黑鲈形体指标(n=28)及体成分(n=4)的影响
Table 4 Effects of enzymatic hydrolysis of soybean meal on body index (n=28) and body composition (n=4) of largemouth bass (%)
组别Group 肥满度CF (g/cm3) 脏体比VSI 肝体比HSI 水分Moisture 灰分Ash 粗蛋白Crude protein 粗脂肪Crude lipid E0 2.04±0.12c 6.91±0.55c 2.48±0.22f 69.78±0.39a 15.68±0.24d 58.55±0.67a 21.21±0.14d E15 1.98±0.15ab 5.73±0.43a 1.19±0.11d 70.72±0.22cd 14.99±0.51bc 60.55±0.01b 19.71±0.56c E20 1.95±0.11ab 5.54±0.48a 0.96±0.08b 69.90±0.22ab 13.95±0.12a 58.95±0.50a 20.87±0.41cd E25 2.00±0.14ab 5.78±0.48a 1.07±0.10c 71.06±0.22d 15.05±0.23bc 60.78±0.88b 17.33±1.02b E30 1.99±0.13ab 5.56±0.46a 0.81±0.09a 70.84±0.27cd 14.89±0.14b 61.08±0.62b 16.10±0.82a SBM 1.91±0.09a 5.60±0.48a 1.01±0.07b 70.34±0.67bc 15.41±0.15c 59.17±0.82a 17.86±0.26b FSBM 1.91±0.11a 6.13±0.49b 1.55±0.13e 70.80±0.23cd 15.12±0.35bc 58.16±0.84a 17.83±1.06b
下载: 导出CSV
表 5 酶解豆粕替代鱼粉对大口黑鲈肝脏功能的影响
Table 5 Effects of enzymatic hydrolysis of soybean meal on liver function of largemouth bass (n=4)
组别Group 过氧化氢酶CAT (U/mg prot) 总抗氧化能力T-AOC (U/mg prot) 丙二醛MDA (nmol/mg prot) 总超氧化歧化酶T-SOD (U/mg prot) 谷草转氨酶AST (U/g prot) 谷丙转氨酶ALT (U/g prot) E0 49.82±0.96f 0.37±0.02a 4.33±0.54e 506.47±16.82f 0.87±0.05a 4.77±0.18b E15 41.44±1.05e 0.61±0.02cd 1.28±0.51bc 398.01±12.31d 1.78±0.05c 5.49±0.28c E20 30.46±1.60c 0.57±0.01c 3.17±0.30d 319.79±7.44a 2.74±0.20f 5.44±0.41c E25 35.95±0.63d 0.86±0.03e 0.73±0.03ab 340.46±8.33b 2.14±0.13d 6.16±0.22d E30 30.66±0.16c 0.65±0.02d 1.32±0.06c 345.18±5.49b 1.77±0.03c 4.39±0.20ab SBM 27.56±0.87b 0.48±0.02b 1.56±0.17c 371.55±6.28c 0.99±0.01a 5.55±0.12c FSBM 23.95±3.59a 0.65±0.05d 0.68±0.05a 453.83±23.75e 1.41±0.03b 4.28±0.08a
下载: 导出CSV
表 6 酶解豆粕替代鱼粉对大口黑鲈脂质代谢指标的影响
Table 6 Effects of enzymatic hydrolysis of soybean meal on lipid metabolism of largemouth bass (n=4)
组别Group 血清游离脂肪酸Serum NEFA (μmol/L) 血清总胆固醇Serum T-CHO (mmol/L) 血清甘油三酯Serum TG
(mmol/L)血清低密度脂蛋白胆固醇Serum LDL-C (mmol/L) 肝脏脂肪含量Liver lipid (%) 肌肉脂肪含量Muscle lipid (%) E0 268.69±6.06a 4.01±0.32a 2.71±0.10a 1.39±0.05a 7.72±0.27a 3.86±0.17d E15 305.72±20.34b 4.66±0.31b 2.62±0.18a 1.77±0.04b 8.57±0.23b 4.50±0.16e E20 366.33±27.50cd 4.71±0.31b 2.87±0.17a 2.10±0.03c 10.82±0.06de 3.80±0.06cd E25 396.77±11.12d 5.28±0.06c 3.29±0.18b 2.40±0.07d 10.88±0.64de 3.50±0.16bc E30 354.21±24.35c 6.52±0.39d 2.79±0.14a 1.43±0.25a 11.47±0.70e 3.34±0.19ab SBM 369.02±16.33cd 5.60±0.28c 3.41±0.18b 1.76±0.05b 10.43±0.54d 3.13±0.27a FSBM 359.93±6.85c 4.32±0.06ab 3.37±0.25b 1.95±0.01c 9.47±0.22c 4.53±0.13e
下载: 导出CSV
-
[1] 周歧存, 麦康森, 刘永坚, 等. 动植物蛋白源替代鱼粉研究进展. 水产学报, 2005, 29(3): 404—410 Zhou Q C, Mai K S, Liu Y J, et al. Advances in animal and plant protein sources in place of fish meal [J]. Journal of Fisheries of China, 2005, 29(3): 404—410
[2] Albertgj T, Marc M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects [J]. Aquaculture, 2008, 285(1): 146—158
[3] Yìldìrìm Ö, Ergün S, Yaman S, et al. Effects of two seaweeds (Ulva lactuca and Enteromorpha linza) as a feed additive in diets on growth performance, feed utilization, and body composition of rainbow trout (Oncorhynchus mykiss) [J]. Kafkas Üniversitesi Veteriner Fakültesi Dergisi, 2009, 15(3): 455—460
[4] 高荣兵, 庄平, 章龙珍, 等. 豆粕替代鱼粉对点篮子鱼生长性能的影响. 水产学报, 2010, 34(10): 1534—1540 Gao R B, Zhuang P, Zhang L Z, et al. Effects of replacement of fish meal by soybean meal on growth characters of Siganidae (Siganus guttatus) [J]. Journal of Fisheries of China, 2010, 34(10): 1534—1540
[5] Boonyaratpalin M, Suraneiranat P, Tunpibal T. Replacement of fish meal with various types of soybean products in diets for the Asian seabass, Lates calcarifer [J]. Aquaculture, 1998, 161(1–4): 67—78
[6] 张帆, 张文兵, 麦康森, 等. 饲料中豆粕替代鱼粉对大黄鱼生长、消化酶活性和消化道组织学的影响. 中国海洋大学学报(自然科学版), 2012, (s1): 75—82 Zhang F, Zhang W B, Mai K S, et al. Effects of replacement of dietary fish meal by soybean meal on growth digestive enzyme activity and digestive tract histology of Juvenile Large Yellow Croaker, Pseudosciaena crocea R [J]. Periodical of Ocean University of China, 2012, (s1): 75—82
[7] Song Z, Li H, Wang J, et al. Effects of fishmeal replacement with soy protein hydrolysates on growth performance, blood biochemistry, gastrointestinal digestion and muscle composition of juvenile starry flounder (Platichthys stellatus) [J]. Aquaculture, 2014, 426—427(1): 96—104
[8] 宋志东, 王际英, 李培玉, 等. 酶解大豆蛋白替代鱼粉对星斑川鲽幼鱼生长、血液生化和体组成的影响. 水生生物学报, 2016, 40(1): 165—172 doi: 10.7541/2016.23 Song Z D, Wang J Y, Li P Y, et al. Effects of replacement fish meal with soy protein hydrolysates on growth, blood biochemistry, body composition of juvenile starry flounder Platichthys stellatus Pallas [J]. Acta Hydrobiologica Sinica, 2016, 40(1): 165—172 doi: 10.7541/2016.23
[9] 王广军, 吴锐全, 谢骏, 等. 人工配合饲料饲养大口黑鲈效果研究. 饲料工业, 2005, 26(4): 22—23 doi: 10.3969/j.issn.1001-991X.2005.04.007 Wang G J, Wu Y Q, Xie J, et al. The study on the effects of feeding largemouth bass with artificial feed [J]. Feed Industry, 2005, 26(4): 22—23 doi: 10.3969/j.issn.1001-991X.2005.04.007
[10] 李永娟, 霍雅文, 杜瑜, 等. 大口黑鲈营养生物学及配合饲料研究进展. 饲料研究, 2016, (23): 39—44 Li Y J, Huo Y W, Du Y, et al. Research advances on nutritional biology and compound feed of largemouth bass [J]. Feed Research, 2016, (23): 39—44
[11] 李二超, 陈立侨. 大口黑鲈的营养需要研究进展. 现代农业科技, 2011, (21): 312—316 doi: 10.3969/j.issn.1007-5739.2011.21.198 Li E C, Chen L Q. Research advances on nutrient requirements of Micropterus salmoides [J]. Modern Agricultural Sciences and Technology, 2011, (21): 312—316 doi: 10.3969/j.issn.1007-5739.2011.21.198
[12] AOAC. Official methods of the analysis of the association of official analytical chemists [S]. 15th ed. Gaithersburg, Maryland, USA: Association of Official Analytical Chemists, 1990
[13] 乔德亮, 李思发, 凌去非, 等. 白斑狗鱼耗氧率和窒息点的研究. 上海水产大学学报, 2005, 14(2): 202—206 Qiao D L, Li S F, Ling Q F, et al. Study on oxygen consumption rate and suffocation point of white spot pike (Esox lucius) [J]. Journal of Shanghai Fisheries University, 2005, 14(2): 202—206
[14] 王维娜, 牛东红, 商利新, 等. 低温对日本沼虾耗氧率、排氨率和Na+/K+ ATPase比活力的影响. 应用与环境生物学报, 2004, 10(5): 602—604 doi: 10.3321/j.issn:1006-687X.2004.05.013 Wang W N, Niu D H, Shang L X, et al. Effect of low temperature on oxygen consumption, ammonia-N excretion and Na+/K+ ATPase of Macrobrachium nipponense [J]. Chinese Journal of Applied & Environmental Biology, 2004, 10(5): 602—604 doi: 10.3321/j.issn:1006-687X.2004.05.013
[15] 李学丽, 王际英, 宋志东, 等. 酶解豆粕替代鱼粉对珍珠龙胆石斑鱼幼鱼生长和主要代谢酶活力的影响. 海洋渔业, 2017, 39(5): 529—538 doi: 10.3969/j.issn.1004-2490.2017.05.006 Li X L, Wang J Y, Song Z D, et al. Effects of fishmeal replacement by hydrolyzed soybean meal on growth, body composition, digestive and metabolic enzyme activities of juvenile Epinephelus fuscoguttatus♀× Epinephelus lanceolatus ♂ [J]. Marine Fisheries, 2017, 39(5): 529—538 doi: 10.3969/j.issn.1004-2490.2017.05.006
[16] Rep M, Koshio S, Ishikawa M, et al. Soy peptide inclusion levels influence the growth performance, proteolytic enzyme activities, blood biochemical parameters and body composition of Japanese flounder, Paralichthys olivaceus [J]. Aquaculture, 2012, 321(3): 252—258
[17] 王建峰, 乐国伟. 小肽在动物营养中的应用. 饲料工业, 2006, 27(7): 9—11 doi: 10.3969/j.issn.1001-991X.2006.07.004 Wang J F, Le G W. Application of peptide in animal nutrition [J]. Feed Industry, 2006, 27(7): 9—11 doi: 10.3969/j.issn.1001-991X.2006.07.004
[18] Newey H, Smyth D H. Intracellular hydrolysis of dipeptides during intestinal absorption [J]. The Journal of Physiology, 1960, 152(2): 367—380 doi: 10.1113/jphysiol.1960.sp006493
[19] 张会平, 雷宁利. 小肽营养及应用研究. 畜禽业, 2006, (24): 20—23 doi: 10.3969/j.issn.1008-0414.2006.24.006 Zhang H P, Lei N L. The advance on the nutrition and utilization of small- peptide [J]. Livestock and Poultry Industry, 2006, (24): 20—23 doi: 10.3969/j.issn.1008-0414.2006.24.006
[20] Lee J, Roh K B, Kim S C, et al. Soy peptide-induced stem cell proliferation: involvement of ERK and TGF-β1 [J]. Journal of Nutritional Biochemistry, 2012, 23(10): 1341—1351 doi: 10.1016/j.jnutbio.2011.08.003
[21] 李昭林. 低鱼粉饲料中添加牛磺酸对黄鳝生长、免疫及代谢的影响. 硕士学位论文, 湖南农业大学, 湖南. 2016 Li Z L. Effect of taurine on growth, immune and metabolism in rice filed eel (Monopterus albus) fed low fish meal [D]. Thesis for Master of Science. Agricultural University of Hunan, Hunan. 2016
[22] 郑银桦, 彭聪, 吴秀峰, 等. 酵母酶解物对大口黑鲈生长性能、脂类代谢及肠道组织结构的影响. 动物营养学报, 2015, 27(5): 1605—1612 doi: 10.3969/j.issn.1006-267x.2015.05.033 Zheng Y H, Peng C, Wu X F, et al. Effects of hydrolyzed yeast on growth performance, lipids metabolism and intestinal structure of largemouth bass (Micropterus salmoides) [J]. Chinese Journal of Animal Nutrition, 2015, 27(5): 1605—1612 doi: 10.3969/j.issn.1006-267x.2015.05.033
[23] 陆烝, 应朝阳, 郑斌, 等. 植物肽的生理功能及其在饲料中的应用研究进展. 中国饲料, 2015, (12): 30—32, 36 Lu Z, Ying C Y, Zheng B, et al. The metabolic mechanism and physiological function of plant peptide and its application in feed [J]. China Feed, 2015, (12): 30—32, 36
[24] 侯丽. 大豆多肽与运动医学研究进展. 甘肃医药, 2012, 31(12): 907—909 doi: 10.3969/j.issn.1004-2725.2012.12.012 Hou L. Research advances on soybean polypeptide and sports medicine [J]. Gansu Medical Journal, 2012, 31(12): 907—909 doi: 10.3969/j.issn.1004-2725.2012.12.012
[25] Lin S, Luo L. Effects of different levels of soybean meal inclusion in replacement for fish meal on growth, digestive enzymes and transaminase activities in practical diets for juvenile tilapia, Oreochromis niloticus× O. aureus [J]. Animal Feed Science & Technology, 2011, 168(1): 80—87
[26] 钱曦, 王桂芹, 周洪琪, 等. 饲料蛋白水平及豆粕替代鱼粉比例对翘嘴红鲌消化酶活性的影响. 动物营养学报, 2007, 19(2): 182—187 doi: 10.3969/j.issn.1006-267X.2007.02.017 Qian X, Wang G Q, Zhou H Q, et al. Effect of dietary protein on the activities of digestive enzymes of topmouth culter (Erythroculter ilishaeformis Bleeker) [J]. Chinese Journal of Animal Nutrition, 2007, 19(2): 182—187 doi: 10.3969/j.issn.1006-267X.2007.02.017
[27] Xu Q Y. Effects of replacement of fish meal by soy protein isolate on the growth, digestive enzyme activity and serum biochemical parameters for juvenile amur sturgeon (Acipenser schrenckii) [J]. Asian-Australasian Journal of Animal Sciences, 2012, 25(11): 1588—1594 doi: 10.5713/ajas.2012.12192
[28] 关胜军, 吴锐全, 谢骏, 等. 两种饲料对大口黑鲈生长、消化道指数和消化酶活性的影响. 饲料工业, 2007, 28(2): 32—36 doi: 10.3969/j.issn.1001-991X.2007.02.010 Guan S J, Wu R Q, Xie J, et al. Effects of two feeds on growth, digestive tract index and the activities of digestive enzymes of largemouth bass (Micropterus salmoides) [J]. Feed Industry, 2007, 28(2): 32—36 doi: 10.3969/j.issn.1001-991X.2007.02.010
[29] 关胜军, 吴锐全, 谢骏, 等. 大口黑鲈主要消化器官蛋白酶活力的研究. 大连海洋大学学报, 2007, 22(1): 32—36 doi: 10.3969/j.issn.1000-9957.2007.01.007 Guan S J, Wu R Q, Xie J, et al. Protease activities in digestive organ in largemouth bass (Micropterus salmoides) [J]. Journal of Dalian Fisheries University, 2007, 22(1): 32—36 doi: 10.3969/j.issn.1000-9957.2007.01.007
[30] Zhao Z X, Song C Y, Xie J, et al. Effects of fish meal replacement by soybean peptide on growth performance, digestive enzyme activities, and immune responses of yellow catfish Pelteobagrus fulvidraco [J]. Fisheries Science, 2016, (4): 1—9
[31] 陈路, 张日俊. 生物活性肽(或寡肽)饲料添加剂的研究与应用. 动物营养学报, 2004, 16(2): 12—14 doi: 10.3969/j.issn.1006-267X.2004.02.003 Chen L, Zhang R J. Bioactive peptide or oligopeptide feed additive research and applications [J]. Acta Zoonutrimenta Sinica, 2004, 16(2): 12—14 doi: 10.3969/j.issn.1006-267X.2004.02.003
[32] 丁辉景, 刘刚, 张力. 单胃动物小肽吸收利用的研究进展. 饲料研究, 2007, (4): 29—32 doi: 10.3969/j.issn.1002-2813.2007.04.010 Ding H J, Liu G, Zhang L. Research advances on absorption and utilization of small peptide in monogastric animals [J]. Feed Research, 2007, (4): 29—32 doi: 10.3969/j.issn.1002-2813.2007.04.010
[33] 郑云峰, 许云英, 徐玉娟. 蛋白质营养中小肽的研究新进展. 饲料研究, 2006, (2): 16—18 Zheng Y F, Xu Y Y, Xu Y J. Research advances on small peptides in protein nutrition [J]. Feed Research, 2006, (2): 16—18
[34] 成艳波, 张月星, 董智勇, 等. 小麦蛋白替代鱼粉及大豆蛋白对日本黄姑鱼(Nibea japonica)和黑鲷(Sparus macrocephalus)血清生化指标及肝脏抗氧化指标的影响. 渔业科学进展, 2017, 38(3): 106—114 doi: 10.11758/yykxjz.20160426001 Cheng Y B, Zhang Y X, Dong Z Y, et al. Effects of dietary replacement of fish meal and soy protein by wheat gluten on plasma biochemical indices and liver anti-oxidative indices of Nibea japonica and Sparus macrocephalus [J]. Progress in Fishery Sciences, 2017, 38(3): 106—114 doi: 10.11758/yykxjz.20160426001
[35] 刘文斌. 饼粕蛋白酶解产物对异育银鲫生长发育影响及其生物效价分析的研究. 硕士学位论文. 南京农业大学, 南京. 2005 Liu W B. Effect of plant protein hydrolysates on the growth development of Alogymogenetic crucian and bio-active value analysis of plant protein hydrolysates [D]. Thesis for Master of Science. Agricultural University of Nanjing, Nanjing. 2005
[36] 邓成萍, 薛文通, 孙晓琳, 等. 不同分子量段大豆多肽功能特性的研究. 食品科学, 2006, 27(5): 109—112 doi: 10.3321/j.issn:1002-6630.2006.05.020 Deng C P, Xue W T, Sun X L, et al. Functional properties of different molecular weight segments of soybean peptides [J]. Food Science, 2006, 27(5): 109—112 doi: 10.3321/j.issn:1002-6630.2006.05.020
[37] Yan Q, Xie S, Zhu X, et al. Dietary methionine requirement for juvenile rockfish, Sebastes schlegeli [J]. Aquaculture Nutrition, 2007, 13(3): 163—169 doi: 10.1111/anu.2007.13.issue-3
[38] 施永海, 张根玉, 张海明, 等. 配合饲料和活饵料对刀鲚幼鱼生长、存活和消化酶、非特异性免疫酶、代谢酶及抗氧化酶活性的影响. 水产学报, 2014, 38(12): 2029—2038 Shi Y H, Zhang G Y, Zhang H M, et al. Effects of compound feed and live feed on the growth, survival, and the activities of digestive enzyme, nonspecific immunity enzyme, metabolic enzyme, and antioxidant enzyme of young fish Coilia nasus [J]. Journal of Fisheries of China, 2014, 38(12): 2029—2038
[39] 侯勇. 水温和饵料对幼蟹氮及能量代谢、能量收支的影响. 硕士学位论文, 苏州大学, 苏州. 2009 Hou Y. Effects of water temperature and food types on energy and nitrogen budget and metabolism of Chinese mitten-handed crab (Eriocheir sinensis) juveniles [D]. Thesis for Master of Science. Soochow University, Suzhou. 2009
[40] Kaushik S J, Coves D, Dutto G, et al. Almost total replacement of fish meal by plant protein sources in the diet of a marine teleost, the European seabass, Dicentrarchus labrax [J]. Aquaculture, 2004, 230(1): 391—404
[41] Yang Y H, Wang Y Y, Lu Y, et al. Effect of replacing fish meal with soybean meal on growth, feed utilization and nitrogen and phosphorus excretion on rainbow trout (Oncorhynchus mykiss) [J]. Aquaculture International, 2011, 19(3): 405—419 doi: 10.1007/s10499-010-9359-y
[42] Robaina L, Moyano F J, Izquierdo M S, et al. Corn gluten and meat and bone meals as protein sources in diets for gilthead seabream (Sparus aurata): Nutritional and histological implications [J]. Aquaculture, 1997, 157(3—4): 347—359
[43] 潘鲁青. 环境因子对甲壳动物渗透调节与免疫力的影响. 硕士学位论文, 中国海洋大学, 青岛. 2004 Pan L Q. Effects of environment factors on osmoregulation and immune parameters of crustaceans [D]. Thesis for Master of Science. Ocean University of China, Qingdao. 2004
[44] 罗莉, 叶元土, 林仕梅, 等. 日粮必需氨基酸模式对草鱼生长及蛋白质周转的影响. 水生生物学报, 2003, 27(3): 278—282 doi: 10.3321/j.issn:1000-3207.2003.03.012 Luo L, Ye Y T, Lin S M, et al. Effect of dietary essential amino acid pattern of the growth and protein turnover of grass carp (Ctenopharyngodon Idellus) [J]. Acta Hydrobiologica Sinica, 2003, 27(3): 278—282 doi: 10.3321/j.issn:1000-3207.2003.03.012
[45] 强俊, 孙意岚, 黄永, 等. 饲料中添加几丁聚糖对吉富罗非鱼幼鱼生长性能、免疫调控与抗海豚链球菌感染的影响. 动物营养学报, 2015, 27(6): 1769—1778 Qiang J, Sun Y L, Huang Y, et al. Effects of chitosan supplementation on growth performance, immunomodulatory and against Streptococcus iniae infection of juvenile GIFT tilapia (Oreochromis niloticus) [J]. Chinese Jour-nal of Animal Nutrition, 2015, 27(6): 1769—1778
[46] Krajnovic O M, Ozretic B. Detection and evaluation of hepatic intoxication in fish [J]. Journal of Hydrology, 1992, 340(3—4): 233—243
-
期刊类型引用(19)
1. 何明,贠彪,钱雪桥,肖伟伟,解绶启. 酶解棉籽蛋白替代鱼粉及添加单宁酸对斑点叉尾鮰生长性能和肠道健康的影响. 饲料工业. 2024(02): 93-101 .  百度学术
百度学术
2. 徐文杰,朱波,蔡明浪,戴济鸿,钟蕾,邵闯,胡毅,陈开健. 低鱼粉饲料中添加酶解鱼小肽对牛蛙蝌蚪生长、变态、肝脏代谢及抗氧化能力的影响. 动物营养学报. 2024(01): 478-489 . 百度学术
3. 马兴宇,唐忠林,陈树桥,周国勤,徐钢春. 转食饲料对大口黑鲈幼鱼的存活率、抗氧化酶和消化酶活性及肠道菌群的影响. 中国水产科学. 2024(04): 403-415 . 百度学术
4. 刘崇万,朱晓华,王明华,张娇,蔡青和,徐志华,任娣,李昊泽. 胰酶对大口黑鲈幼鱼生长、肝脏与肠道健康的影响. 水生生物学报. 2024(12): 1993-2005 . 本站查看
5. 胡俊茹,蓝青,张春燕,宋鹏,陈晓瑛,黄玲玲. 发酵豆粕替代鱼粉对大口黑鲈生长、生化指标及肠道健康的影响. 饲料工业. 2023(02): 66-76 . 百度学术
6. 胡雪晴,郎宇曦,吕福军,黄紫薇,潘雨佳,李应东. 酶解豆粕饲料对草鱼肠道微生物菌群结构的影响. 饲料工业. 2023(02): 89-94 . 百度学术
7. 胡骏鹏,黄鑫,陈敬帮,周迪,徐杰,廖灿青,董毅. 不同蛋白原料对仔猪生长性能及血清生化指标的影响. 中国饲料. 2023(09): 169-172 . 百度学术
8. 唐兴刚,郭伟干,袁明贵,孙凯辉,王晓珊,向蓉. 发酵豆粕的抗氧化活性及其对尖吻鲈生长性能的研究. 广东畜牧兽医科技. 2023(04): 20-24 . 百度学术
9. 向倩,张蕉南,罗悦,王帅杰,陶敏,徐丽,吴建军,蒋思婧. 酶解蛋白原料替代鱼粉在水产中的应用. 饲料研究. 2023(24): 148-153 . 百度学术
10. Xuanyi Yang,Xinyan Zhi,Ziling Song,Guanghui Wang,Xumin Zhao,Shuyan Chi,Beiping Tan. Flesh quality of hybrid grouper(Epinephelus fuscoguttatus ♀ ×Epinephelus lanceolatus ♂) fed with hydrolyzed porcine mucosasupplemented low fishmeal diet. Animal Nutrition. 2022(01): 114-124 . 必应学术
11. 时于惠,祝书杰,谭北平,张文兵,周小秋,郜卫华,迟淑艳. 大口黑鲈对六种非粮蛋白源的表观消化率. 水生生物学报. 2022(08): 1187-1196 . 本站查看
12. 钟立强,王海骁,王明华,张世勇,姜虎成,陈校辉. 不同饵料对大口黑鲈生长性能和肠道微生物的影响. 水生生物学报. 2022(11): 1609-1617 . 本站查看
13. 朱昱铮,申亮,郭冉,荆冰妍,梁倍宁,高佳朋. 鱼肽和蛋白酶替代部分鱼粉对凡纳滨对虾生长、消化酶及肠道菌群的影响. 大连海洋大学学报. 2022(05): 756-763 . 百度学术
14. 潘雨佳,黄紫薇,吕福军,郎宇曦,赵莹莹,李应东. 酶解豆粕对克氏原螯虾生长及肠道微生物菌群的影响. 饲料研究. 2022(20): 44-48 . 百度学术
15. 钟立强,王海骁,王明华,张世勇,姜虎成,陈校辉. 鲈-蟹混养模式下不同饵料对大口黑鲈生长和肠道菌群的影响. 基因组学与应用生物学. 2022(08): 1653-1664 . 百度学术
16. 陈晓瑛,陈绍坚,黄文,王国霞,黄敏,吴浩敏,曹俊明,孙育平. 菌酶协同发酵豆粕替代鱼粉对大口黑鲈生长性能、血清生化、免疫和抗氧化指标及肝脏组织形态的影响. 动物营养学报. 2021(05): 2848-2863 . 百度学术
17. 敬庭森,周明瑞,李哲,李玉林,孙文波,梁钧畑,王新靓,叶华,成廷水,罗辉. 大豆小肽蛋白替代鱼粉对黄颡鱼幼鱼生长性能、消化酶活性和抗氧化功能的影响. 渔业科学进展. 2021(05): 149-157 . 百度学术
18. 吴远彩,李日美,申光荣,黄放,杨奇慧,谭北平,迟淑艳. 小肽对凡纳滨对虾生长、抗氧化能力、非特异性免疫及肠道菌群结构的影响. 广东海洋大学学报. 2021(05): 1-9 . 百度学术
19. 许合金,曾楠. 饲粮添加小肽优蛋白对南美白对虾生长性能和免疫性能的影响. 广东畜牧兽医科技. 2021(06): 64-70 . 百度学术
其他类型引用(18)
计量
- 文章访问数: 3094
- HTML全文浏览量: 1649
- PDF下载量: 71
- 被引次数: 37