
MOLECULAR CHARACTERISTICS AND GROWTH STATUS OF GLOIOSIPHONIA CAPILLARIS
-
摘要: 对分布在辽宁大连黑石礁、付家庄、石槽、金石滩、獐子岛与海洋岛沿岸的黏管藻[Gloiosiphonia capillaris (Hudson) Carmichael]的外部形态、营养与生殖结构、生物量、成熟个体比例与成熟个体生物量/总生物量(R/T指数)的变化、温度性质以及rbcL、COⅠ基因序列进行了详细研究。结果表明: (1) 黏管藻配子体为雌雄同体, 直立, 单生或丛生, 主轴明显, 固着器呈圆盘状, 质地胶质, 颜色为红色或紫红色。采集于獐子岛与海洋岛的藻体长度和宽度明显高于其他采集地点; (2) 藻体由皮层及髓部组成, 皮层由6—10层细胞组成, 髓部存在假根丝细胞。成熟囊果小, 突出于藻体表面, 呈球形或半球形, 通常2—4个囊果聚集在一起; (3) 黏管藻配子体的生物量在6月达到最大, 平均生物量为3.628 g/m2。3—6月成熟个体比例逐渐增大, 6月达到100%; (4) 黏管藻配子体生长周期为3—7月, 温度性质属于温带性; (5) rbcL基因序列分析表明, 6个采集地点的样本间无碱基差异, 与产自加拿大的黏管藻聚在一起, 形成一个独立的分支。COⅠ基因序列分析表明, 6个采集地点的样本间无碱基差异, 形成一个独立的分支, 均确定为黏管藻。Abstract: This study analyzed the external morphology, vegetative and reproductive structure, growth process, biomass variation, mature individual proportion, R/T index, as well as the large subunit of ribulose-1, 5-bisphosphate carboxylase/oxygenase (rbcL) gene and the partial cytochrome coxidase Ⅰ (COⅠ) gene sequences of Gloiosiphonia capillaris (Hudson) Carmichael populations collected in the coast of Heishijiao, Fujiazhuang, Shicao, Jinshitan, Zhangzidao and Haiyangdao in Dalian. The results indicated that the gametophytes of G. capillaris were androgynous, solitary or caespitose habit, and that the main axes were erect, disciformi affixus, gelatinous in texture, red or purplish red. The length and width of thallus collected in Zhangzidao and Haiyangdao were significantly higher than those in other sites. Thallus was composed of cortex that had 6—10 cell layers and medulla that was consisted of many rhizoidal filaments. The size of mature cystocarp was small and protruded from the thallus. Cystocarps were spherical or hemispherical, and often became a group with 2—4 members. The maximum biomass of gametophyte appeared in June with an average value of 3.628 g/m2. The proportions of mature individuals increased gradually from March to June and got 100% in June. The gametophyte growth cycle was from March to July, and the temperature property was temperate. According to the phylogenetic tree of rbcL gene sequences, there was no sequence divergence among the 6 sites and clustered in a single monophyletic subclade with the sample from Canada. There was also no sequence divergence among the 6 sites based on the phylogenetic tree of COⅠ gene sequences. The samples were clustered in a single monophyletic subclade and identified as G. capillaris.
-
黏管藻[Gloiosiphonia capillaris (Hudson) Carmichael]隶属于红藻门(Rhodophyta), 杉藻目(Gigartinales), 黏管藻科(Gloiosiphoniaceae), 黏管藻属(Gloiosiphonia Carmichael)。广泛分布于大西洋和太平洋, 国内主要分布于辽宁、山东、福建, 生长在潮间带的石沼中[1]。目前黏管藻属在全世界共报道了3个种, 模式种为黏管藻[G. capillaris (Hudson) Carmichael], 另外2个种为加利福尼亚黏管藻[G. californica (Farlow) J. Agardh]和轮生黏管藻(G. verticillaris Farlo)。对于黏管藻的研究源于18世纪70年代。Hudson[2]在英国发现了该种, 并将其命名为Fucus capillaris Hudson。Berkeley[3]发现F. capillaris Hudson的形态结构明显区别于墨角藻属(Fucus)的其他种, 依据该种建立了黏管藻属并将该种作为黏管藻属的模式种。Edelstein[4]对黏管藻的生活史进行了研究, 发现黏管藻生活史为异型世代交替, 其四分孢子体为壳状。夏邦美[1]与曾呈奎[5]对黏管藻的外部形态和内部结构进行了研究。Saunders等[6]对产自加拿大的黏管藻的核酮糖1,5-二磷酸羧化酶/氧化酶大亚基(rbcL)基因序列进行了分析, 从分子水平明确了其分类地位。
随着近几年大连海域海洋资源的过度开发以及围填海等行为, 在谋求社会发展和经济效益的同时也导致了近海岸海洋生态系统功能退化以及海水富营养化形势加剧[7]。环境的剧烈变化导致大连海域潮间带大型底栖海藻的物种组成以及群落结构发生了改变, 如大连獐子岛沿岸叉节藻(Amphiroa zonata)、柔质仙菜(Ceramium tenerrimum)等海藻分布范围越来越小甚至消失, 石莼(Ulva lactuca)、浒苔(Ulva prolifera)等海藻大量繁殖, 绿潮现象越来越严重[8]。藻类的大量繁殖会带来严重的危害, 如强壮硬毛藻(Chaetomorpha valida)的大量繁殖给大连瓦房店和复州湾的海参养殖业带来了巨大的经济损失[9], 浒苔(U. prolifera)的大规模暴发引发绿潮灾害, 给黄海近岸海域造成了严重的社会影响[10], 同时藻类的大量死亡会消耗水体中的溶解氧, 导致水体表层和底层的物理、化学性质产生不利变化[11]。笔者在标本的采集调查时发现大连海域刺松藻(Codium fragile)、海萝(Gloiopeltis furcate)、亮管藻(Hyalosiphonia caespitosa)等常见海藻的分布范围由于海洋生态环境的恶化而变得越来越小, 甚至消失。因此有必要对大连海域海藻的种类、分布及生物量等进行详细调查与评估, 达到物种保护和灾害预防的目的。黏管藻作为大型经济海藻, 在大连沿岸广泛分布, 其含有的生物活性物质具有抗炎、抗肿瘤、抗氧化和免疫抑制的作用[12]。但关于其生长状态、资源现状、生长周期与繁殖盛期、温度性质等研究未见报道。本研究将对分布在大连沿岸的黏管藻的外部形态、营养与生殖结构、生物量、成熟个体比例与成熟个体生物量/总生物量(R/T指数)的变化、温度性质等进行详细研究, 并通过rbcL和COⅠ基因序列分析结合形态结构对其进行准确的鉴定。研究结果将明确大连沿岸黏管藻的资源现状; 确定其生长周期与繁殖盛期, 为其物种的保护和繁育提供理论依据; 确定黏管藻在海洋水体污染的监测中的作用和地位, 为海洋生态环境的监测提供理论依据, 为分布于全国其他海域的黏管藻的研究提供参考。
1. 材料与方法
1.1 采集地点
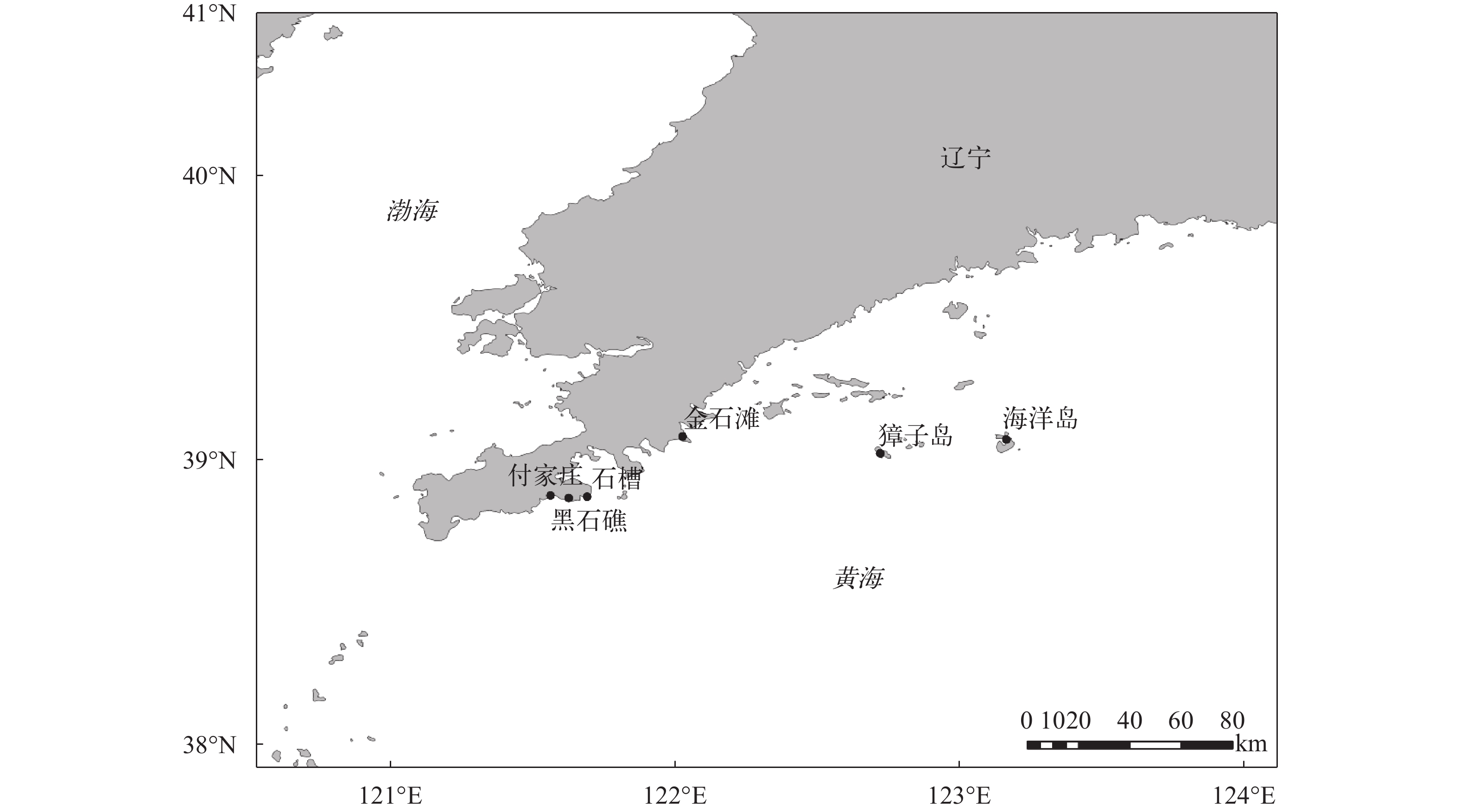
本研究样本均采自辽宁省大连海域礁石较多、生态环境保护较好的区域。采集地点的布设如图 1所示, 共布设了6个采集地点, 分别位于黑石礁(121°34′7.91″E, 38°52′40.5″N)、付家庄(121° 37′58.3″E, 38°52′8.77″N)、石槽(121°41′51.0″E, 38°52′24.7″N)、金石滩(122°1′54.8″E, 39°5′6.15″N)、獐子岛(122°43′25.9″E, 39°1′32.6″N)和海洋岛(123°9′56.4″E, 39°4′27.7″N)。
1.2 样本的采集与处理
根据海洋生物调查规范[13], 在2016年3月至2018年2月每个月大潮期间进行样本采集, 包括定性采集和定量采集。
定性采集: 黏管藻生长在潮间带的岩石上或石沼中(图 2A)。参照王永宇等[14]的方法, 选择结构完整的藻体进行采集, 将藻体在解剖镜下清理干净。部分新鲜藻体用于制作冰冻切片, 使用光学显微镜(Olympus BH2)观察藻体的内部结构, 用照相机(Nikon HFX-ⅡA)拍摄照片并记录结果, 另一部分制成硅胶干燥标本, 用于分子分析。选择结构完整的藻体制成腊叶标本并编号, 保存于辽宁师范大学生命科学学院植物标本馆(LNU)。
![]() 图 2 黏管藻的野外生境与样方设置A. 野外生境(箭头所示); B. 样方设置(25 cm×25 cm)Figure 2. The wild habitat and setting of the quadrat (25 cm×25 cm) for G. capillaris samplingA. Wild habitat (arrowheads); B. The setting of the quadrat (25 cm×25 cm)
图 2 黏管藻的野外生境与样方设置A. 野外生境(箭头所示); B. 样方设置(25 cm×25 cm)Figure 2. The wild habitat and setting of the quadrat (25 cm×25 cm) for G. capillaris samplingA. Wild habitat (arrowheads); B. The setting of the quadrat (25 cm×25 cm)定量采集: 参照王永宇等[14]的方法, 在每个采集地点黏管藻生长旺盛的区域布设2个25 cm×25 cm的样方(图 2B), 采集样方内的黏管藻, 用吸水纸吸干藻体表面水分, 放置在65℃的烘干箱中干燥24h, 称重并记录每个样方内黏管藻的生物量。统计每个样方内成熟藻体的数目, 并计算成熟个体比例与R/T指数(成熟个体的生物量/总生物量)[15]。
1.3 DNA提取与PCR扩增
使用植物基因组DNA提取试剂盒(北京天根公司)进行藻体DNA的提取, 将提取到的DNA进行PCR扩增。引物设计参照姜朋[16]的方法, 引物组合如表 1所示, 均由上海生工生物公司合成。rbcL与COⅠ基因的PCR反应程序参照王永宇等[14]的方法, 检测后的样品均由上海生工生物公司纯化和测序。
表 1 rbcL与COⅠ序列扩增所需引物Table 1. Primers used in the study基因类型
Gene type引物Primer 序列Sequences (5′—3′) rbcL F57 GTAATTCCATATGCTAAAATGGG R1296 TCAGCTGTATCTGTAGAAGTAT F1237 CCAGATGGTATTCAAGCAGGTGC RrbcS TGTGTTGCGGCCGCCCTTGTGTTAGTCTCAC COI GazF1 TCAACAAATCATAAAGATATTGG DumR1 AAAAAYCARAATAAATGTTGA 1.4 rbcL、COⅠ基因序列分析
从每个采集地点选取2个黏管藻样本, 获得12条rbcL序列与12条COⅠ序列。从GenBank中获取了5个种的rbcL序列、3个种的COⅠ序列(表 2)。获取rbcL序列的物种包括产自加拿大的黏管藻、产自美国的黏管藻属的轮生黏管藻、产自加拿大黏管藻科的Plagiospora gracilis, 而产自加拿大的亮管藻(H. caespitosa)和Schimmelmannia plumosa则作为外群种。获取COⅠ序列的物种包括产自加拿大的黏管藻属的轮生黏管藻, 产自山东青岛的亮管藻和产自加拿大的S. plumosa则作为外群种。应用软件Clustalx (1.83)进行序列比对, MEGA6.0进行碱基差异度分析及构建系统树, 使用最大似然法Maximum likelihood (ML)法、临近法Neighbor joining (NJ)法和最大简约法Maximum parsimony (MP)法构建系统发育树。计算遗传距离模型为Number of differences和Kimura 2-Parameter, Substitutions Type为d:Transitions+Transversions, Bootstrap重复1000次[13]。
表 2 rbcL、COⅠ基因序列分析所用物种、采集地点及基因库登录号列表Table 2. List of species for rbcL and COⅠ gene sequence analysis with the sample location and the accession numbers of GenBank物种Species 采集地点Sample location 登录号Accession number rbcL COⅠ Gloiosiphonia capillaris (Hudson) Carmichael 大连黑石礁(LNU2016050201) MG645982 MG645994 G. capillaris (Hudson) Carmichael 大连黑石礁(LNU2016050202) MG645983 MG645995 G. capillaris (Hudson) Carmichael 大连付家庄(LNU2016050203) MG645984 MG645996 G. capillaris (Hudson) Carmichael 大连付家庄(LNU2016050204) MG645985 MG645997 G. capillaris (Hudson) Carmichael 大连石槽(LNU2016050305) MG645986 MG645998 G. capillaris (Hudson) Carmichael 大连石槽(LNU2016050306) MG645987 MG645999 G. capillaris (Hudson) Carmichael 大连金石滩(LNU2016050307) MG645988 MG646000 G. capillaris (Hudson) Carmichael 大连金石滩(LNU2016050308) MG645989 MG646001 G. capillaris (Hudson) Carmichael 大连獐子岛(LNU2016050509) MG645990 MG646002 G. capillaris (Hudson) Carmichael 大连獐子岛(LNU2016050510) MG645991 MG646003 G. capillaris (Hudson) Carmichael 大连海洋岛(LNU2016051811) MG645992 MG646004 G. capillaris (Hudson) Carmichael 大连海洋岛(LNU2016051812) MG645993 MG646005 G. capillaris (Hudson) Carmichael 加拿大 KU382056 G. verticillaris Farlow 美国 GVU04195 G. verticillaris Farlow 加拿大 KM254896 Plagiospora gracilis Kuckuck 加拿大 KU905059 Hyalosiphonia caespitosa Okamura 加拿大 KT310684 Hyalosiphonia caespitosa Okamura 山东青岛 KC782901 Schimmelmannia plumosa (Setchell) I. A. Abbott 加拿大 KU382061 KM254413 2. 结果
2.1 黏管藻配子体个体形态的变化
六个采集地点的黏管藻配子体均直立, 单生或丛生, 质地胶质, 红色或紫红色, 固着器呈圆盘状。主轴明显, 线形圆柱状或亚圆柱状。分枝较密, 以对生、互生的方式排列在轴的所有面, 所有分枝又密被有与它们相似的较短的次分枝。但不同采集地点的配子体长度和宽度存在差异, 采自獐子岛与海洋岛的配子体在长度和宽度上明显高于其他4个采集地点(表 3)。四分孢子体壳状, 很难确定所采集的壳状海藻是否为黏管藻, 故不做研究。
表 3 不同采集点的黏管藻配子体外部形态的比较Table 3. The morphological characteristics of G. capillaris gametophyte from different localities采集地点Sample location 藻体颜色The color of thallus 长度Length (cm) 宽度Width (mm) 黑石礁Heishijiao 紫红色 8—12 1—1.5 付家庄Fujiazhuang 红色 7—10 1—1.5 石槽Shicao 紫红色 7—10 1—1.5 金石滩Jinshitan 紫红色 10—15 1—2 獐子岛Zhangzi Island 红色 9—18 1.5—2 海洋岛Haiyang Island 红色 12—20 1.5—2.5 3月初, 配子体幼苗开始出现, 藻体长度小于1 cm (图 3A)。3月中旬, 藻体出现分枝, 藻体长度达到2—2.6 cm (图 3B)。3月末, 藻体长度达到3.7—4.2 cm (图 3C)。4月下旬, 藻体长度达到3.8—6.3 cm, 部分藻体上可发现成熟的囊果(图 3D)。5月, 藻体快速生长, 藻体长度达到5.2—9.1 cm, 囊果集中成熟(图 3E)。6月, 藻体长度达到6—15 cm (图 3F), 其中最大长度可达20 cm (图3G), 囊果全部发育成熟。7月, 当黏管藻的果孢子释放完成后, 藻体颜色逐渐变浅, 腐烂消失。8月至次年2月未采集到黏管藻配子体。
![]() 图 3 黏管藻配子体个体形态的变化A. 3月上旬配子体幼苗; B—F. 3—6月藻体外部形态; G. 最大成熟配子体Figure 3. Different morphology of G. capillaris gametophyte at different stagesA. Gametophyte seedlings in early March; B—F. The external morphology of thallus from March to June; G. The largest mature gametophyte
图 3 黏管藻配子体个体形态的变化A. 3月上旬配子体幼苗; B—F. 3—6月藻体外部形态; G. 最大成熟配子体Figure 3. Different morphology of G. capillaris gametophyte at different stagesA. Gametophyte seedlings in early March; B—F. The external morphology of thallus from March to June; G. The largest mature gametophyte2.2 黏管藻配子体的营养结构与生殖结构
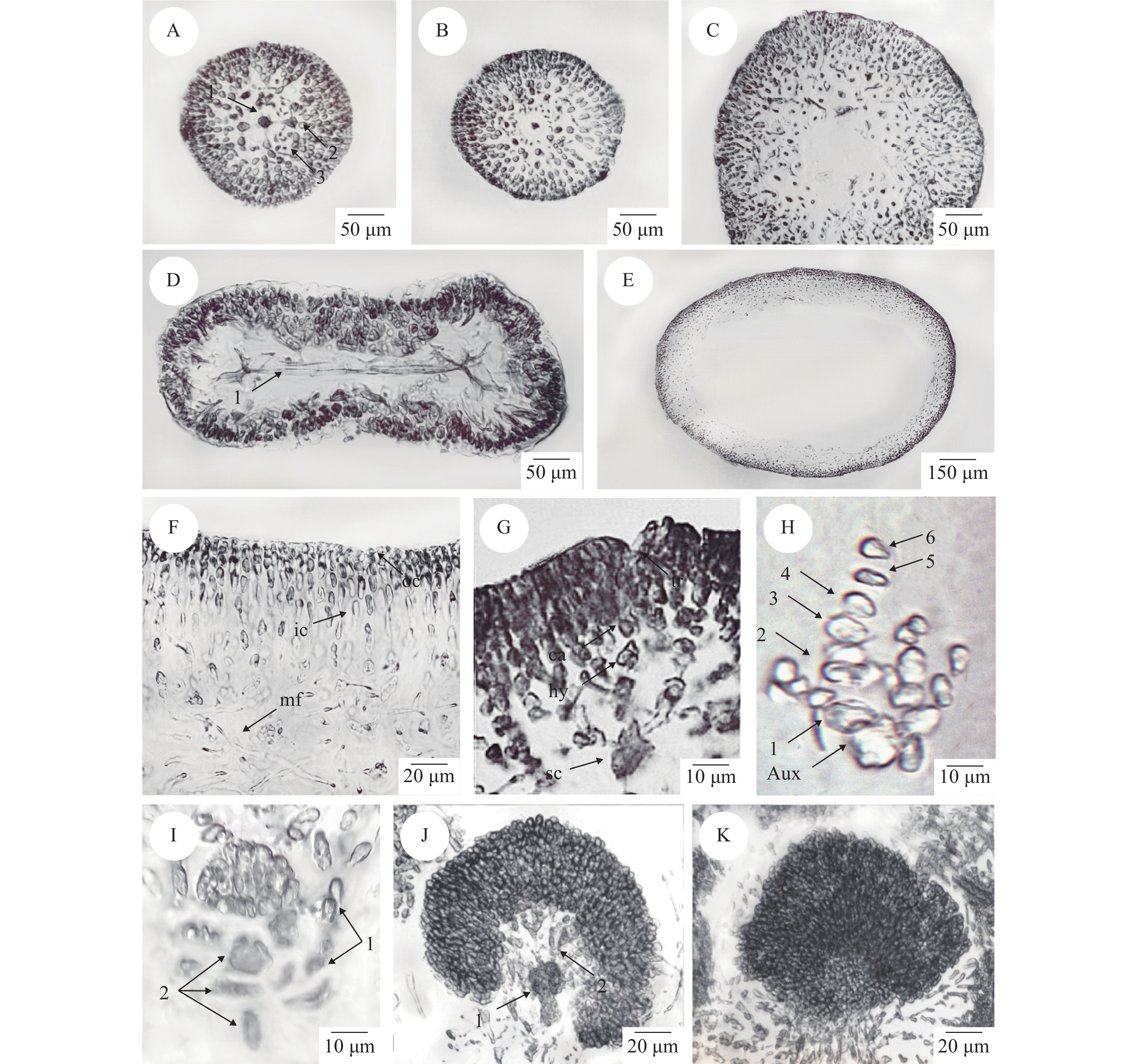
营养结构: 从次分枝顶端的横切面观中可以观察到一个近圆形的中轴细胞以及四个围轴细胞, 围轴细胞和中轴细胞呈十字形排列, 在中轴细胞和围轴细胞间可观察到假根丝细胞(图 4A)。围轴细胞逐渐衍生成排列紧密的皮层细胞, 在衍生完毕后, 围轴细胞消失。在次分枝中端只能观察到皮层细胞、中轴细胞和假根丝细胞(图 4B)。在次分枝末端中轴细胞消失, 只能观察到皮层细胞以及假根丝细胞(图 4C)。分枝与次分枝营养结构一致。从主枝上端的横切中可以观察到髓部从中间开始分裂(图4D)。主枝上端至末端内部中空呈管状, 藻体横切呈椭圆形(图 4E)。皮层由6—10层细胞组成, 厚度为45—80 μm, 外皮层由2—3层排列紧密的圆形或椭圆形细胞构成, 内皮层由4—7层不规则的近方形或卵形细胞构成。髓部充满疏松排列的髓丝, 长15—50 μm, 宽2—3 μm, 在髓部可以观察到假根丝细胞(图 4F)。
![]() 图 4 黏管藻配子体营养结构与生殖结构A. 藻体次分枝顶端的横切面观(1. 中轴细胞; 2. 围轴细胞; 3. 假根丝细胞); B. 藻体次分枝中端的横切面观; C. 藻体次分枝末端的横切面观; D. 藻体主枝上端的横切面观(1. 髓部从中间分裂); E. 藻体主枝中端的横切面观; F. 皮层横切面观(oc. 外皮层细胞; ic. 内皮层细胞; mf. 髓丝); G. 果胞枝生殖枝丛(1—3. 果胞枝生殖枝丛主枝; tr. 受精丝; cp. 果胞; hy. 下位细胞; sc. 支持细胞); H. 辅助细胞枝生殖枝丛(1—6. 辅助细胞生殖枝丛主枝; aux. 辅助细胞); I. 嚢果被包围果孢子(1. 嚢果被; 2. 融合复合体); J. 趋于成熟的囊果(1. 融合复合体; 2. 产孢丝); K. 成熟囊果横切面观Figure 4. The vegetative structure and reproductive structure of G. capillaris gametophyteA. Cross-section on the top of the secondary branch (1. centre axoblast; 2. pericentral cell; 3. rhizoidal filament); B. Cross-section in the middle of the secondary branch; C. Cross-section at the end of the secondary branch; D. Cross-section on the top of the main axes (1. medulla divided from the middle); E. Cross-section in the middle of the main axes; F. Cross-section of the cortex (oc. outer cortex; ic. inner cortex; mf. medulla filament); G. Carpogonial branch ampullae (1—3. the bough of carpogonial branch ampullae; tr. trichogyne; cp. carpogonium; hy. hypogynous cell; sc. supporting cell); H. Auxiliary cell ampullae (1—6. the bough of auxiliary branch ampullae; aux. auxiliary cell); I. Carpospores surrounded by pericarp (1. pericarp; 2. complex fusion); J. Cystocarp tend to maturity (1. complex fusion; 2. gonimoblast); K. Cross-section of mature cystocarp
图 4 黏管藻配子体营养结构与生殖结构A. 藻体次分枝顶端的横切面观(1. 中轴细胞; 2. 围轴细胞; 3. 假根丝细胞); B. 藻体次分枝中端的横切面观; C. 藻体次分枝末端的横切面观; D. 藻体主枝上端的横切面观(1. 髓部从中间分裂); E. 藻体主枝中端的横切面观; F. 皮层横切面观(oc. 外皮层细胞; ic. 内皮层细胞; mf. 髓丝); G. 果胞枝生殖枝丛(1—3. 果胞枝生殖枝丛主枝; tr. 受精丝; cp. 果胞; hy. 下位细胞; sc. 支持细胞); H. 辅助细胞枝生殖枝丛(1—6. 辅助细胞生殖枝丛主枝; aux. 辅助细胞); I. 嚢果被包围果孢子(1. 嚢果被; 2. 融合复合体); J. 趋于成熟的囊果(1. 融合复合体; 2. 产孢丝); K. 成熟囊果横切面观Figure 4. The vegetative structure and reproductive structure of G. capillaris gametophyteA. Cross-section on the top of the secondary branch (1. centre axoblast; 2. pericentral cell; 3. rhizoidal filament); B. Cross-section in the middle of the secondary branch; C. Cross-section at the end of the secondary branch; D. Cross-section on the top of the main axes (1. medulla divided from the middle); E. Cross-section in the middle of the main axes; F. Cross-section of the cortex (oc. outer cortex; ic. inner cortex; mf. medulla filament); G. Carpogonial branch ampullae (1—3. the bough of carpogonial branch ampullae; tr. trichogyne; cp. carpogonium; hy. hypogynous cell; sc. supporting cell); H. Auxiliary cell ampullae (1—6. the bough of auxiliary branch ampullae; aux. auxiliary cell); I. Carpospores surrounded by pericarp (1. pericarp; 2. complex fusion); J. Cystocarp tend to maturity (1. complex fusion; 2. gonimoblast); K. Cross-section of mature cystocarp生殖结构: 黏管藻配子体雌雄同体, 囊果突出藻体表面, 呈球形或半球形, 通常2—4个囊果聚集在一起。果胞枝生殖枝丛与辅助细胞生殖枝丛均由内皮层细胞产生。果胞枝生殖枝丛主枝较短包含3个细胞, 生长于一个较大的颜色较深的支持细胞上, 最上面是一个带有一条细长受精丝的较小的果胞, 其下有一个下位细胞, 受精丝伸直或弯曲指向藻体表面(图 4G)。辅助细胞生殖枝丛主枝较长, 包含6个细胞, 辅助细胞枝末端细胞为辅助细胞。辅助细胞带有3—4个由2—5个细胞组成的侧枝(图 4H)。在受精后, 果胞产生联络丝, 进而与辅助细胞相连, 辅助细胞与枝丛主枝上的其他细胞融合并逐渐形成一个延伸的融合复合体。该复合体末端产生产孢丝, 产孢丝末端形成果孢子, 辅助细胞生殖枝丛的侧枝与周围的髓丝相互连接和缠绕, 形成嚢果被包围果孢子(图 4I)。果孢子不断增多, 可以观察到融合完毕的融合复合体(图 5J)。成熟囊果近球形, 直径100—140 μm。果孢子卵圆形, 直径6—9 μm (图 5K)。囊果成熟后果孢子通过囊孔释放。精子囊未见。
![]() 图 5 基于rbcL序列所构建的ML系统发育树分支上的数字表示自展值(1000次重复), 分别为ML (上)、NJ (中)、MP (下), 只显示自展值大于50%Figure 5. The Maximum likelihood (ML) tree was based on rbcL sequencesNumerals at internal nodes are bootstrap values (1000 replicates) inferred from ML (upper), NJ (middle) and MP (lower). Only values above 50% bootstrap support are shown
图 5 基于rbcL序列所构建的ML系统发育树分支上的数字表示自展值(1000次重复), 分别为ML (上)、NJ (中)、MP (下), 只显示自展值大于50%Figure 5. The Maximum likelihood (ML) tree was based on rbcL sequencesNumerals at internal nodes are bootstrap values (1000 replicates) inferred from ML (upper), NJ (middle) and MP (lower). Only values above 50% bootstrap support are shown2.3 黏管藻配子体的生物量、成熟个体比例与R/T指数的变化
通过统计3—7月每个采集地点每个样方内的数据发现, 3—6月, 黏管藻的平均生物量一直增大, 6月平均生物量达到顶峰, 平均生物量为3.628 g/m2 (表 4), 其中獐子岛和海洋岛采集点的黏管藻的生物量最高, 分别为4.023和4.132 g/m2。7月随着海水温度升高, 平均生物量减小, 8月至次年2月未采集到黏管藻个体。
表 4 黏管藻配子体生物量的变化Table 4. The biomass of G. capillaris gametophyte月份Month 平均生物量Average biomass (g/m2) 生物量Biomass (g/m2) 黑石礁Heishijiao 付家庄Fujiazhuang 石槽Shicao 金石滩Jinshitan 獐子岛Zhangzi Island 海洋岛Haiyang Isalnd 3 0.511 0.478 0.463 0.480 0.493 0.569 0.584 4 1.015 0.965 0.842 0.957 1.034 1.135 1.156 5 2.738 2.478 2.366 2.747 2.895 2.964 2.977 6 3.628 3.230 3.064 3.425 3.896 4.023 4.132 7 1.659 1.442 1.346 1.785 1.559 1.957 1.864 当黏管藻配子体上出现生殖器官果胞与精子囊时, 藻体即成熟。4月下旬黏管藻配子体开始出现生殖器官, 样方内成熟个体比例为10% (表 5), R/T指数为9% (表 6)。5月囊果集中成熟, 成熟个体比例为61%, R/T指数为60%。6月份藻体全部成熟, 成熟个体比例及R/T指数均为100%。
表 5 黏管藻配子体成熟个体比例变化Table 5. The mature individual proportion of G. capillaris gametophyte月份Month 平均成熟个体比例The average of mature individual proportion (%) 成熟个体比例The mature individual proportion (%) 黑石礁Heishijiao 付家庄Fujiazhuang 石槽Shicao 金石滩Jinshitan 獐子岛Zhangzi Island 海洋岛Haiyang Isalnd 3 0 0 0 0 0 0 0 4 10 9 11 10 8 11 11 5 61 59 64 70 54 60 59 6 100 100 100 100 100 100 100 7 100 100 100 100 100 100 100 表 6 黏管藻配子体R/T指数的变化Table 6. The R/T ratio of G. capillaris gametophyte月份Month 平均R/T指数The average of R/T ratio (%) R/T指数The R/T ratio (%) 黑石礁Heishijiao 付家庄Fujiazhuang 石槽Shicao 金石滩Jinshitan 獐子岛Zhangzi Island 海洋岛Haiyang Isalnd 3 0 0 0 0 0 0 0 4 9 9 8 10 8 9 10 5 60 62 68 56 52 59 63 6 100 100 100 100 100 100 100 7 100 100 100 100 100 100 100 2.4 黏管藻配子体的温度性质分析
海藻的温度性质是海藻区系性质中最主要的一项[13]。关于海藻的区系的温度性质, 可分为寒带区系(<0—5℃)、亚寒带区系(5—10℃)、温带区系(10—20℃)、亚热带区系(20—30℃)和热带区系(25—30℃或以上)[17]。黏管藻主要分布在黄海、渤海、东海沿岸, 黏管藻配子体的生长周期为3—7月, 以大连市为例, 根据大连市3—7月海水温度变化分析黏管藻配子体的温度性质。3—6月, 海水温度从5.9℃升至15.3℃, 这段时间黏管藻配子体逐渐生长, 其中5—6月藻体生长速度最快, 平均生物量迅速增大, 海水温度范围为10.6—15.3℃。6—7月, 海水温度从15.3℃ 升至20.7℃, 黏管藻配子体逐渐减少, 8月至次年2月未采集到黏管藻配子体。综上所述, 黏管藻配子体的适宜生长温度在10.6—15.3℃, 故黏管藻配子体的温度性质属于温带性。
2.5 rbcL和COⅠ基因序列分析
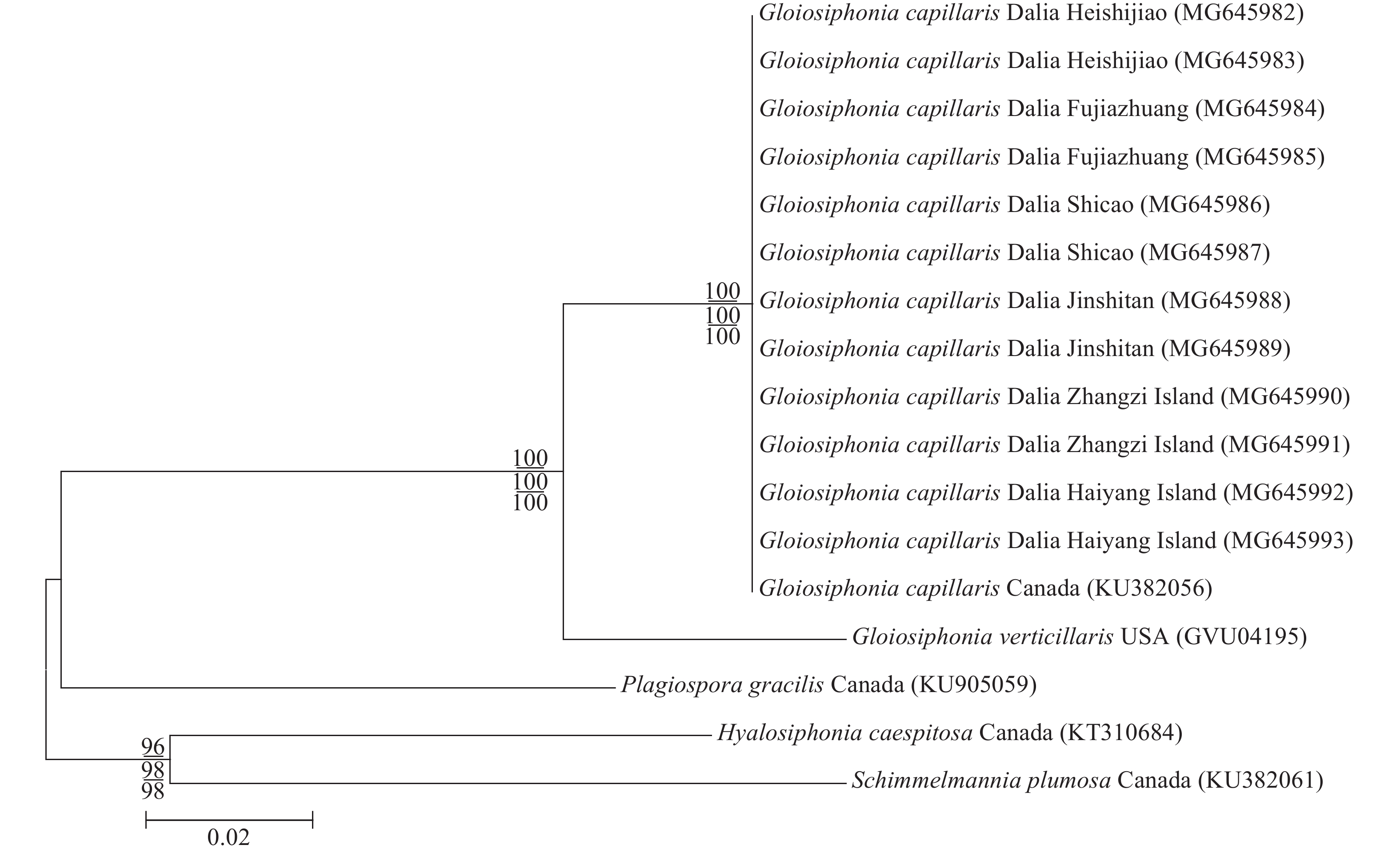
rbcL基因序列分析 本研究从6个地点各获得2条总共12条黏管藻的rbcL序列并提交到GenBank中, 所获得登录号为: MG645982、MG645983、MG645984、MG645985、MG645986、MG645987、MG645988、MG645989、MG645990、MG645991、MG645992和MG645993。共比对17条序列, 对比矫正后序列矩阵长度为1226 bp。基于ML法、NJ法和MP法构建了rbcL序列的系统发育树, 3种方法所构建的发育树具有相似的拓扑结构, 故以ML法所构建系统发育树的拓扑结构显示。结果如图 5所示, 本研究的12个样本之间无碱基差异, 与产自加拿大的黏管藻在系统树中聚在一个分支中, 无碱基差异, 确定为黏管藻。与黏管藻属的轮生黏管藻碱基差异为64 bp (7.97%)。与黏管藻科Plagiospora属的P. gracilis碱基差异为157 bp (12.56%)。与外群种亮管藻属的亮管藻碱基差异为170 bp (13.07%)以及与Schimmelmannia属的S. plumosa碱基差异为185 bp (12.83%)。
COⅠ基因序列分析 本研究从6个地点各获得2条总共12条黏管藻的COⅠ序列并提交到GenBank中, 所获得登录号为: MG645994、MG645995、MG645996、MG645997、MG645998、MG645999、MG646000、MG646001、MG646002、MG646003、MG646004和MG646005。共比对15条序列, 对比矫正后序列矩阵长度为664 bp。基于ML法、NJ法和MP法构建了COⅠ序列的系统发育树, 3种方法所构建的发育树具有相似的拓扑结构, 故以ML法所构建系统发育树的拓扑结构显示。结果如图 6所示, 本研究的12个样本之间无碱基差异。而与黏管藻属的轮生黏管藻碱基差异为44 bp (6.35%), 与外群种亮管藻属的亮管藻碱基差异为109 bp (8.83%), 与Schimmelmannia属的S. plumosa碱基差异为110 bp (8.98%)。
![]() 图 6 基于COⅠ序列所构建的ML系统发育树分支上的数字表示自展值(1000次重复), 分别为ML (上)、NJ (中)、MP (下), 只显示自展值大于50%Figure 6. The Maximum likelihood (ML) tree based on the COⅠ sequencesNumerals at internal nodes are bootstrap values (1000 replicates) inferred from ML (upper), NJ (middle) and MP (lower). Only values above 50% bootstrap support are shown
图 6 基于COⅠ序列所构建的ML系统发育树分支上的数字表示自展值(1000次重复), 分别为ML (上)、NJ (中)、MP (下), 只显示自展值大于50%Figure 6. The Maximum likelihood (ML) tree based on the COⅠ sequencesNumerals at internal nodes are bootstrap values (1000 replicates) inferred from ML (upper), NJ (middle) and MP (lower). Only values above 50% bootstrap support are shown3. 讨论
采自大连6个地点的黏管藻样本的外部形态、营养结构和生殖结构与夏邦美[1]和曾呈奎[5]的描述一致。在外部形态上, 本研究的样本具有较密的次生分枝, 而轮生黏管藻(G. verticillaris)具有3—6个渐细的轮生分枝, 无次生分枝, 二者易于区分。因此从形态学上确定本研究的样本是黏管藻。根据rbcL序列构建的ML系统发育树(图 5)显示, 本研究中的12个样本与产自加拿大的黏管藻聚在一起, 形成一个独立的分支, 自展值为100, 区别于其他种, 验证了本研究的12个样本为黏管藻。根据COⅠ序列构建的ML系统发育树(图 6)显示, 本研究的12个样本间聚在一起, 形成一个独立的分支, 自展值为99, 与同属的轮生黏管藻(G. verticillaris)碱基差异为44 bp (6.352%), 并与该种形成一个较大的黏管藻属分支, Le Gall等[18]研究了DNA条形码技术对于研究海藻种类多样性的作用, 认为海藻碱基序列差异大于2%属于种间差异, 验证了本研究的12个样本为黏管藻。
Newton[19]对分布于欧洲的黏管藻配子体的生长周期进行了调查, 发现其春天和夏天有分布, 这与我们的调查结果一致。由于黏管藻的四分孢子体为壳状, 故很难确定黏管藻四分孢子体的生长周期及温度性质。本研究发现大连獐子岛与海洋岛的黏管藻配子体在外部形态与平均生物量上明显大于其他采集地点。经过调查发现獐子岛与海洋岛远离内陆, 海洋生态环境保护较好, 海水富营养化程度较低, 所以我们推测黏管藻的生长对于环境条件要求较高, 对N、P等营养盐及重金属耐受力较弱, 可作为指示植物在海洋水体污染的监测中起到重要作用。王永宇等[14]通过对单条胶黏藻(Dumontia simplex)的生长状态进行了研究, 发现在海水富营养化严重的海域其外部形态和生物量更大, 这说明海水富营养化对不同藻类的生长状态影响不同。笔者在标本采集时发现大连海域近几年黏管藻的分布范围逐渐减小, 经研究发现这一现象与大连海域近几年5—6月份石莼(U. lactuca)的大量繁殖有关。海水富营养化为石莼的大量繁殖提供有利的条件[20], 此时正是黏管藻的繁殖盛期, 生长受到抑制的黏管藻与大量繁殖的石莼相互竞争处于弱势, 黏管藻的生活空间被侵占, 最终使其分布范围逐渐减小, 因此有必要对该种继续跟踪调查。此外, 笔者发现大连海域一些常见海藻如刺松藻(Codium fragile)、海萝(Gloiopeltis furcate)等由于海洋生态环境的恶化其分布范围也变得越来越小。据统计大连地区海洋中蕴藏的海藻有71种海藻属于药用海藻[21], 具有抗凝血、抗氧化等多种生物活性[22—24]。因此有必要对大连地区海藻物种以及海洋生态环境进行保护来实现海洋环境和海洋经济的可持续发展。
本研究明确了分布在大连沿海的黏管藻及其资源现状, 明确了其生长周期与繁殖盛期并分析了该种分布减少的原因, 为该种的保护、繁育提供详细的数据支撑并推测黏管藻可作为指示植物在海洋水体污染的监测中具有重要的作用, 为分布于全国其他海域的黏管藻的研究提供参考。
-
![]()
图 2 黏管藻的野外生境与样方设置
A. 野外生境(箭头所示); B. 样方设置(25 cm×25 cm)
Figure 2. The wild habitat and setting of the quadrat (25 cm×25 cm) for G. capillaris sampling
A. Wild habitat (arrowheads); B. The setting of the quadrat (25 cm×25 cm)
![]()
图 3 黏管藻配子体个体形态的变化
A. 3月上旬配子体幼苗; B—F. 3—6月藻体外部形态; G. 最大成熟配子体
Figure 3. Different morphology of G. capillaris gametophyte at different stages
A. Gametophyte seedlings in early March; B—F. The external morphology of thallus from March to June; G. The largest mature gametophyte
![]()
图 4 黏管藻配子体营养结构与生殖结构
A. 藻体次分枝顶端的横切面观(1. 中轴细胞; 2. 围轴细胞; 3. 假根丝细胞); B. 藻体次分枝中端的横切面观; C. 藻体次分枝末端的横切面观; D. 藻体主枝上端的横切面观(1. 髓部从中间分裂); E. 藻体主枝中端的横切面观; F. 皮层横切面观(oc. 外皮层细胞; ic. 内皮层细胞; mf. 髓丝); G. 果胞枝生殖枝丛(1—3. 果胞枝生殖枝丛主枝; tr. 受精丝; cp. 果胞; hy. 下位细胞; sc. 支持细胞); H. 辅助细胞枝生殖枝丛(1—6. 辅助细胞生殖枝丛主枝; aux. 辅助细胞); I. 嚢果被包围果孢子(1. 嚢果被; 2. 融合复合体); J. 趋于成熟的囊果(1. 融合复合体; 2. 产孢丝); K. 成熟囊果横切面观
Figure 4. The vegetative structure and reproductive structure of G. capillaris gametophyte
A. Cross-section on the top of the secondary branch (1. centre axoblast; 2. pericentral cell; 3. rhizoidal filament); B. Cross-section in the middle of the secondary branch; C. Cross-section at the end of the secondary branch; D. Cross-section on the top of the main axes (1. medulla divided from the middle); E. Cross-section in the middle of the main axes; F. Cross-section of the cortex (oc. outer cortex; ic. inner cortex; mf. medulla filament); G. Carpogonial branch ampullae (1—3. the bough of carpogonial branch ampullae; tr. trichogyne; cp. carpogonium; hy. hypogynous cell; sc. supporting cell); H. Auxiliary cell ampullae (1—6. the bough of auxiliary branch ampullae; aux. auxiliary cell); I. Carpospores surrounded by pericarp (1. pericarp; 2. complex fusion); J. Cystocarp tend to maturity (1. complex fusion; 2. gonimoblast); K. Cross-section of mature cystocarp
![]()
图 5 基于rbcL序列所构建的ML系统发育树
分支上的数字表示自展值(1000次重复), 分别为ML (上)、NJ (中)、MP (下), 只显示自展值大于50%
Figure 5. The Maximum likelihood (ML) tree was based on rbcL sequences
Numerals at internal nodes are bootstrap values (1000 replicates) inferred from ML (upper), NJ (middle) and MP (lower). Only values above 50% bootstrap support are shown
![]()
图 6 基于COⅠ序列所构建的ML系统发育树
分支上的数字表示自展值(1000次重复), 分别为ML (上)、NJ (中)、MP (下), 只显示自展值大于50%
Figure 6. The Maximum likelihood (ML) tree based on the COⅠ sequences
Numerals at internal nodes are bootstrap values (1000 replicates) inferred from ML (upper), NJ (middle) and MP (lower). Only values above 50% bootstrap support are shown
表 1 rbcL与COⅠ序列扩增所需引物
Table 1 Primers used in the study
基因类型
Gene type引物Primer 序列Sequences (5′—3′) rbcL F57 GTAATTCCATATGCTAAAATGGG R1296 TCAGCTGTATCTGTAGAAGTAT F1237 CCAGATGGTATTCAAGCAGGTGC RrbcS TGTGTTGCGGCCGCCCTTGTGTTAGTCTCAC COI GazF1 TCAACAAATCATAAAGATATTGG DumR1 AAAAAYCARAATAAATGTTGA  下载: 导出CSV
下载: 导出CSV
表 2 rbcL、COⅠ基因序列分析所用物种、采集地点及基因库登录号列表
Table 2 List of species for rbcL and COⅠ gene sequence analysis with the sample location and the accession numbers of GenBank
物种Species 采集地点Sample location 登录号Accession number rbcL COⅠ Gloiosiphonia capillaris (Hudson) Carmichael 大连黑石礁(LNU2016050201) MG645982 MG645994 G. capillaris (Hudson) Carmichael 大连黑石礁(LNU2016050202) MG645983 MG645995 G. capillaris (Hudson) Carmichael 大连付家庄(LNU2016050203) MG645984 MG645996 G. capillaris (Hudson) Carmichael 大连付家庄(LNU2016050204) MG645985 MG645997 G. capillaris (Hudson) Carmichael 大连石槽(LNU2016050305) MG645986 MG645998 G. capillaris (Hudson) Carmichael 大连石槽(LNU2016050306) MG645987 MG645999 G. capillaris (Hudson) Carmichael 大连金石滩(LNU2016050307) MG645988 MG646000 G. capillaris (Hudson) Carmichael 大连金石滩(LNU2016050308) MG645989 MG646001 G. capillaris (Hudson) Carmichael 大连獐子岛(LNU2016050509) MG645990 MG646002 G. capillaris (Hudson) Carmichael 大连獐子岛(LNU2016050510) MG645991 MG646003 G. capillaris (Hudson) Carmichael 大连海洋岛(LNU2016051811) MG645992 MG646004 G. capillaris (Hudson) Carmichael 大连海洋岛(LNU2016051812) MG645993 MG646005 G. capillaris (Hudson) Carmichael 加拿大 KU382056 G. verticillaris Farlow 美国 GVU04195 G. verticillaris Farlow 加拿大 KM254896 Plagiospora gracilis Kuckuck 加拿大 KU905059 Hyalosiphonia caespitosa Okamura 加拿大 KT310684 Hyalosiphonia caespitosa Okamura 山东青岛 KC782901 Schimmelmannia plumosa (Setchell) I. A. Abbott 加拿大 KU382061 KM254413
下载: 导出CSV
表 3 不同采集点的黏管藻配子体外部形态的比较
Table 3 The morphological characteristics of G. capillaris gametophyte from different localities
采集地点Sample location 藻体颜色The color of thallus 长度Length (cm) 宽度Width (mm) 黑石礁Heishijiao 紫红色 8—12 1—1.5 付家庄Fujiazhuang 红色 7—10 1—1.5 石槽Shicao 紫红色 7—10 1—1.5 金石滩Jinshitan 紫红色 10—15 1—2 獐子岛Zhangzi Island 红色 9—18 1.5—2 海洋岛Haiyang Island 红色 12—20 1.5—2.5
下载: 导出CSV
表 4 黏管藻配子体生物量的变化
Table 4 The biomass of G. capillaris gametophyte
月份Month 平均生物量Average biomass (g/m2) 生物量Biomass (g/m2) 黑石礁Heishijiao 付家庄Fujiazhuang 石槽Shicao 金石滩Jinshitan 獐子岛Zhangzi Island 海洋岛Haiyang Isalnd 3 0.511 0.478 0.463 0.480 0.493 0.569 0.584 4 1.015 0.965 0.842 0.957 1.034 1.135 1.156 5 2.738 2.478 2.366 2.747 2.895 2.964 2.977 6 3.628 3.230 3.064 3.425 3.896 4.023 4.132 7 1.659 1.442 1.346 1.785 1.559 1.957 1.864
下载: 导出CSV
表 5 黏管藻配子体成熟个体比例变化
Table 5 The mature individual proportion of G. capillaris gametophyte
月份Month 平均成熟个体比例The average of mature individual proportion (%) 成熟个体比例The mature individual proportion (%) 黑石礁Heishijiao 付家庄Fujiazhuang 石槽Shicao 金石滩Jinshitan 獐子岛Zhangzi Island 海洋岛Haiyang Isalnd 3 0 0 0 0 0 0 0 4 10 9 11 10 8 11 11 5 61 59 64 70 54 60 59 6 100 100 100 100 100 100 100 7 100 100 100 100 100 100 100
下载: 导出CSV
表 6 黏管藻配子体R/T指数的变化
Table 6 The R/T ratio of G. capillaris gametophyte
月份Month 平均R/T指数The average of R/T ratio (%) R/T指数The R/T ratio (%) 黑石礁Heishijiao 付家庄Fujiazhuang 石槽Shicao 金石滩Jinshitan 獐子岛Zhangzi Island 海洋岛Haiyang Isalnd 3 0 0 0 0 0 0 0 4 9 9 8 10 8 9 10 5 60 62 68 56 52 59 63 6 100 100 100 100 100 100 100 7 100 100 100 100 100 100 100
下载: 导出CSV
-
[1] 夏邦美. 中国海藻志. 见: 第二卷红藻门, 第三册隐丝藻目. 北京: 科学出版社. 2004, 57—59 Xia B M. Flora Algarum Marinarum Sinicarum [M]. In: Tomus II Rhodophyta. Part III. Cryptonemiales. Beijing: Science Press. 2004, 57—59
[2] Hudson. Flora Anglica Tomus II [M]. Strand: Londini. 1778, 335—690
[3] Berkeley. Gleanings of British Algae [M]. Whitefish: Kessinger Publishing. 2010, 13—16
[4] Edelstein T. The life history of Gloiosiphonia capillaris (Hudson) Carmichael 1 [J]. Phycologia, 1970, 9(1): 55—59 doi: 10.2216/i0031-8884-9-1-55.1
[5] 曾呈奎. 中国黄渤海海藻. 北京: 科学出版社. 2008, 156—157 Zeng S K. Seaweed from the Yello and Bohai Seas [M]. Beijing: Science Press. 2008, 156—157
[6] Saunders G W, Filloramo G, Dixon K, et al. Multigene analyses resolve early diverging lineages in the Rhodymeniophycidae (Florideophyceae, Rhodophyta) [J]. Journal of Phycology, 2016, 52(4): 505—522 doi: 10.1111/jpy.2016.52.issue-4
[7] 狄乾斌, 韩增林. 大连市围填海活动的影响及对策研究. 海洋开发与管理, 2008, 25(10): 122—126 doi: 10.3969/j.issn.1005-9857.2008.10.030 Di Q B, Han Z L. The impacts of reclamation activities in Dalian and the countermeasures [J]. Ocean Development and Management, 2008, 25(10): 122—126 doi: 10.3969/j.issn.1005-9857.2008.10.030
[8] 贾潇博. 獐子岛潮间带底栖海藻三十年变迁. 硕士学位论文, 辽宁师范大学, 大连. 2016 Jia X B. Thirty years changes of benthic algae in intertidal zone of Zhangzi Island [D]. Thesis for Master of Science. Liaoning Normal University, Dalian. 2016
[9] 迟永雪, 王丽梅, 栾日孝, 等. 中国硬毛藻属新记录种-强壮硬毛藻. 水产科学, 2009, 28(3): 162—163 doi: 10.3969/j.issn.1003-1111.2009.03.013 Chi Y X, Wang L M, Luan R X, et al. Chaetomorpha valida-A new recorded green alga species in genus Chaetomorpha Kuetzing in China [J]. Fisheries Science, 2009, 28(3): 162—163 doi: 10.3969/j.issn.1003-1111.2009.03.013
[10] 王宗灵, 傅明珠, 肖洁, 等. 黄海浒苔绿潮研究进展. 海洋学报, 2018, 40(2): 1—13 Wang Z L, Fu M Z, Xiao J, et al. Progress on the study of the Yellow Sea green tides caused by Ulva prolifera [J]. Acta Oceanologica Sinica, 2018, 40(2): 1—13
[11] 刘春兰. 青岛浒苔暴发因素的探讨及对策. 安徽农业科学, 2013, 41(4): 1695—1698 doi: 10.3969/j.issn.0517-6611.2013.04.111 Liu C L. Discussion on factors for outbreak of Enteromorpha in Qingdao and countermeasures [J]. Journal of Anhui Agricultural Sciences, 2013, 41(4): 1695—1698 doi: 10.3969/j.issn.0517-6611.2013.04.111
[12] 方玉春, 王毓, 罗艳, 等. 中国药用红藻资源调查及开发利用建议. 中国海洋药物, 2010, 29(3): 63—67 Fang Y C, Wang Y, Luo Y, et al. Investigation of medicinal red algae resources and proposals for its development and utilization [J]. Chinese Journal of Marine Drugs, 2010, 29(3): 63—67
[13] 中国国家标准化管理委员会. 海洋调查规范——第6部分: 海洋生物调查. 北京: 中国标准出版社, 2007, 48—50 China National Standardization Administration. Specifications for oceanographic survey [A]. In: Part 6: Marine biological survey [C]. Beijing: Standards Press of China. 2007, 48—50
[14] 王永宇, 王昊林, 李佳俊, 等. 单条胶黏藻Dumontia simplex Cotton的生长状态研究与分子分析. 海洋与湖沼, 2018, 49(4): 829—838 Wang Y Y, Wang H L, Li J J, et al. Growth status and molecular systematics of Dumontia simplex [J]. Oceanologia et Limnologia Sinica, 2018, 49(4): 829—838
[15] Hasegawa Y, Fukuhara E. Ecological studies on Iridophycus cornucopiae (P. et R.) Setch. et Gardn. 3. On the seasonal change of the number of female gametophytes and tetrasporophytes II [J]. Bull Hokkaido Reg Fish Research Laboratory, 1955, (12): 16—22
[16] 姜朋. 四种杉藻目红藻的形态观察、个体生态学及分子系统学研究. 硕士学位论文, 辽宁师范大学, 大连. 2016 Jiang P. The morphologic observation, autecological studies and sequences analyses for four red algae in Gigartinales [D]. Thesis for Master of Science. Liaoning Normal University, Dalian. 2016
[17] 曾呈奎, 张峻甫. 关于海藻区系性质的分析. 海洋与湖沼, 1960, 3(3): 177—187 Zeng C K, Zhang J F. An analysis of the nature of marine algal flora [J]. Oceanologia et Limnologia Sinica, 1960, 3(3): 177—187
[18] Le Gall L, Saunders G W. DNA barcoding is a powerful tool to uncover algal diversity: a case study of the Phyllophoraceae (Gigartinales, Rhodophyta) in the Canadian flora [J]. Journal of Phycology, 2010, 46(2): 374—389 doi: 10.1111/jpy.2010.46.issue-2
[19] Newton L. A Handbook of the British Seaweeds [M]. London: British Museum. 1931, 478
[20] 马元庆, 邢红艳, 宋秀凯, 等. 烟台金沙滩绿潮爆发的基本状况与应对措施. 齐鲁渔业, 2010, 27(4): 10—12 Ma Y Q, Xing H Y, Song X K, et al. The basic conditions and response measures of green tide outbreak in Yantai Golden Beach [J]. Shandong Fisheries, 2010, 27(4): 10—12
[21] 李熙宜. 药用海藻的开发利用与展望. 大连海洋大学学报, 1989, 4(1): 11—22 Li X Y. Prospect of exploitation and utilization of medicinal seaweeds [J]. Journal of Dalian Ocean University, 1989, 4(1): 11—22
[22] 易杨华, 焦炳华. 现代海洋药物学. 北京: 科学出版社. 2006 Yi Y H, Jiao B H. Modern Marine Pharmacology [M]. Beijing: Science Press. 2006
[23] 严君, 施松善, 王顺春, 等. 蜈蚣藻中一种硫酸半乳聚糖的分离纯化、结构鉴定及其抗新生血管生成活性. 高等学校化学学报, 2008, 29(9): 1755—1759 doi: 10.3321/j.issn:0251-0790.2008.09.010 Yan J, Shi S S, Wang S C, et al. Sulfated galactan from Grateloupia filicina: isolation, purification, structure characterization and its antiangiogenic activity [J]. Chemical Journal of Chinese Universities, 2008, 29(9): 1755—1759 doi: 10.3321/j.issn:0251-0790.2008.09.010
[24] Dias P F, Jr S J, Vendruscolo L F, et al. Antiangiogenic and antitumoral properties of a polysaccharide isolated from the seaweed Sargassum stenophyllum [J]. Cancer Chemotherapy Pharmacology, 2005, 56(4): 436—446 doi: 10.1007/s00280-004-0995-7



计量
- 文章访问数: 8120
- HTML全文浏览量: 5919
- PDF下载量: 69