
ROLE OF ENDOPLASMIC RETICULUM STRESS IRE1 PATHWAY IN HEPATOCYTE APOPTOSIS OF GRASS CARP CTENOPHARYNGODON IDELLA INDUCED BY SODIUM NITRITE
-
摘要: 为探究亚硝酸钠诱导草鱼肝细凋亡中内质网应激IRE1通路的作用, 草鱼(Ctenopharyngodon idellus)肝细胞L8824分别置于亚硝酸钠浓度为0、5、20和50 mg/L暴露12h和24h。同时采用三磷酸肌醇受体拮抗剂2-APB和IRE1α抑制剂STF-083010分别和20 mg/L亚硝酸钠共同孵育L8824细胞24h。检测细胞凋亡以及c-Jun氨基末端激酶(c-Jun N-terminal kinase, jnk)、B淋巴细胞瘤-2(B-cell lymphoma-2, bcl-2)、bcl-2相关X蛋白(bcl-2 associated X protein, bax)、caspase9、caspase3、肌醇酶1α (inositol requiring enzyme 1α, ire1α)、X盒结合蛋白 1s (X-box binding protein 1s, xbp1s)和葡萄糖调节蛋白78 (glucose-regulated protein78, grp78)基因的表达和细胞质内钙离子浓度。结果表明: 与对照组相比, 亚硝酸钠处理组细胞凋亡率显著上升, jnk、bax、caspase9、caspase3、ire1α、xbp1s和grp78 mRNA的表达显著升高, bcl-2 mRNA的表达量显著下降, 细胞质内钙离子浓度显著上升。与亚硝酸钠单处理组相比, STF-083010处理组细胞凋亡率显著下降, ire1α、xbp1s、grp78、jnk、bax、caspase3 mRNA表达量显著下降, bcl-2 mRNA表达量显著上升, 2-APB和STF-083010两个处理组细胞质内钙离子浓度均显著下降。结果显示, 高浓度的亚硝酸钠可诱导草鱼肝细胞凋亡和钙离子紊乱, 内质网应激IRE1通路发挥了作用。Abstract: Nitrite, a common pollutant in aquaculture, is an intermediate product of nitrogen cycle in ecosystem. To explore the mechanisms of sodium nitrite-induced cell apoptosis, grass carp liver cell (L8824) were exposed to four concentrations of sodium nitrite (0, 5 mg/L, 20 mg/L and 50 mg/L) with or without treatments of phosphoinositide receptor antagonist 2-APB and IRE1 inhibitors STF-083010. Cell apoptosis related gene expression of jnk, bcl-2, bax, caspase9, caspase3, ire1α, xbp1s and grp78 and the cytoplasmic calcium ion concentration were assessed. The results showed that nitrite significantly increased the apoptosis rate, cytoplasm calcium ion concentration and mRNA levels of jnk, bax, caspase9, caspase3, ire1α, xbp1s and grp78 and significantly decreased bcl-2 mRNA level, which were reversed by the STF-083010 treatment. Besides, both 2-APB and STF-083010 reduced the sodium nitrite-induced cytoplasmic calcium ion. These results indicate that endoplasmic reticulum stress-related IRE1 pathway plays pivotal role nitrite-mediated L8824 cell apoptosis and calcium dyshomeostasis.
-
Keywords:
- Sodium nitrite /
- IRE1 pathway /
- Apoptosis /
- Calcium ion
-
大银鱼(Protosalanx hyalocranius Abbott, 1901)和太湖新银鱼(Neosalanx taihuensis Chen, 1956)分别隶属于鲑形目(Salmoniformes)银鱼科(Salangidae)的大银鱼属和新银鱼属, 主要分布在我国黄河、淮河和长江中下游及其附属湖泊, 是我国重要的经济鱼类[1]。20世纪70年代以来, 2种银鱼被引种移植到全国众多湖泊和水库[2]。大银鱼主要被移植到长江以北[3], 太湖新银鱼则多被移植到黄河以南[4]。2种银鱼的生命周期均为一年, 个体小、生长快、世代离散, 种群波动显著 [1, 5]。
过度捕捞是影响鱼类资源可持续利用的重要因素, 分为生长型过度捕捞和补充型过度捕捞[6]。由于银鱼世代离散, 补充型过度捕捞导致剩余产卵群体数量不足, 进而使种群下一世代数量减少, 被认为是其资源量下降的重要原因[7]。单位补充量产卵群体生物量(Spawning Biomass-recruitment, SBR)模型根据种群的生长、死亡参数和种群资源动态, 估算捕捞后产卵群体的残存量, 进而评估种群的资源利用状况[8, 9]。当SBR残存量低于未开发状态的20%时, 通常视为补充型过度捕捞, 可能导致种群崩溃[10, 11]。平衡产量模型是渔业资源评估最常用的Beverton-Holt模型的简化形式[6, 12], 常用于分析开捕时间和捕捞强度对渔获产量的影响, 并据此制定合理的渔业管理方案[13, 14]。
洪泽湖(33°06—40′N, 118°10—52′E)是中国第四大淡水湖, 水域面积约2069 km2。大银鱼和太湖新银鱼是洪泽湖最主要的经济鱼类之一, 2010年以来年渔产量最高曾达106 kg[15], 但年际波动较大, 且近年来呈现下降趋势(江苏省洪泽湖渔业管理委员会办公室, 未发表数据)。洪泽湖银鱼捕捞采用银鱼拖网进行, 每年1月1日至8月8日为禁捕期, 其中5月21日左右允许捕捞1周。据渔民反映, 在每年12月下旬捕捞期的最后阶段2种银鱼均已很难捕捞、渔获量极低。本研究对洪泽湖大银鱼和太湖新银鱼的周年生长和资源量动态进行了分析, 评估种群捕捞利用状况, 进而优化渔业管理方式, 以期为洪泽湖银鱼的资源可持续利用管理提供参考。
1. 材料与方法
1.1 采样水域与采样方法
2015年7月至2016年6月每月中旬, 对洪泽湖大银鱼和太湖新银鱼进行周年采样(图 1)。采样使用渔民用的银鱼拖网进行, 拖网网口宽11.6 m, 高2.3 m; 网体长约25 m, 由4个网片组成, 侧面网片网目为1.2 cm×1.2 cm, 上下网片网目为1.5 cm×1.5 cm; 末端网袋长1.3 m, 网目为0.3 cm×0.3 cm。采样时在机动渔船左右两侧各设置一个拖网, 拖网上沿恰好没入水中, 渔船在固定起点(A, 33°17′19″N, 118°47′17″E)和终点(B, 33°19′58″N, 118°46′50″E)间拖行5 km, 拖速为4.0 km/h。采样在上午9:00—12:00间进行。样品采集后立即置于冰水保温箱中, 带回实验室计数, 进行生物学测定。每次采样2种银鱼生物学测定数量均在100尾以上, 当样品数量不足时, 增加拖行时间。
![]() 图 1 洪泽湖, 示大银鱼和太湖新银鱼采集时拖网拖行区间起点(A)和终点(B)Figure 1. Hongze Lake, showing the trawling area (from A to B) for P. hyalocranius and N. taihuensis collection
图 1 洪泽湖, 示大银鱼和太湖新银鱼采集时拖网拖行区间起点(A)和终点(B)Figure 1. Hongze Lake, showing the trawling area (from A to B) for P. hyalocranius and N. taihuensis collection1.2 数据获取与分析
体长-体重关系和生长率 在实验室对大银鱼和太湖新银鱼的渔获物进行计数和称重。测定个体体长(L, 0.1 mm), 称体重(W, 0.01 g), 解剖鉴定性别, 繁殖季节肉眼确定雌性性腺发育分期[16, 17]。体长-体重关系采用W=a×Lb进行拟合。采用单因子方差分析(ANOVA)检验各月2种银鱼体长和体重差异显著性, 并采用Bonferroni方法进行多重比较。分别求出2种银鱼各月的平均体长和体重, 计算相邻月份间体长和体重瞬时生长率(gl, gw)[18]:
${g_l}{\rm{ = ln}}{L_{n + 1}}{\rm{ - ln}}{L_n}$

${g_w}={\rm{ln}}{W_{n + 1}} - {\rm{ ln}}{W_n}$

式中, Ln 和Wn 分别为某月(n)大银鱼或太湖新银鱼的平均体长和体重; Ln+1 和Wn+1则为下一月份的平均体长和体重。
最适生长方程 洪泽湖大银鱼的繁殖季节为12至次年2月, 盛期为1月中旬; 太湖新银鱼春群的繁殖季节为3—4月, 盛期为3月(未发表数据)。据此, 我们将1月25日和3月25日分别设为大银鱼和太湖新银鱼的出生时间, 即0龄, 并推算2种银鱼各月采样时间对应的年龄。采用Logistic生长方程(Logistic growth function, Logistic GF)、Gompertz生长方程(Gompertz growth function, Gompertz GF)和von Bertalanffy生长方程(Von Bertalanffy Growth Function, VBGF)拟合2种银鱼的年龄与体长生长关系, 使用最小二乘法估算各生长方程的参数, 依据赤井信息准则(Akaike’s information criterion, AIC)选择最适生长方程[19]。3种生长方程分别为:
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
式中, t表示样本年龄; t0为理论生长起点, 即体长Lt=0时的起始年龄; L∞为渐近体长; K为生长系数。生长方程拟合采用R-3.3.1软件进行[20]。
种群死亡系数 定义单位捕捞努力量渔获量(Catch per Unit Effort, CPUE)为拖网拖行1 km2水域面积的渔获物个体数量和生物量。使用CPUE逐月数量变动数据, 结合洪泽湖当前的银鱼捕捞管理方式(8月9日—12月31日为捕捞期, 同时5月21—28日可对银鱼资源进行为期一周的捕捞), 求解以下方程组, 计算捕捞死亡系数F和自然死亡系数M [21]:
${N_{{{t_2}}}} = \left\{ {\begin{array}{*{20}{c}}\!\!\!\! {{N_{{{t_1}}}}{{\rm{e}}^{ - M({{t_2}} - {{t_1}})}}\;\;\left( \text{当不存在人为捕捞作用时} \right)}\\\!\!\!\! {{N_{{{t_1}}}}{{\rm{e}}^{ - Z({{t_2}} - {{t_1}})}}\;\;\left( \text{当存在人为捕捞作用时} \right)}\end{array}} \right.$

This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
模型构建 使用平衡产量模型(The equilibrium output model)估算种群单位补充量渔产量随开捕年龄和捕捞强度的变化[6, 13]:
$\begin{aligned}Y = & F \times {N_0} \times {{\rm{e}}^{ - M \times {{r}}}} \times {W_\infty } \times \\ & \left( {\frac{1}{Z} - \frac{{3{{\rm{e}}^{ - K \times {{r}}}}}}{{Z + K}} + \frac{{3{{\rm{e}}^{ - 2K \times {{r}}}}}}{{Z + 2K}} - \frac{{{{\rm{e}}^{ - 3K \times {{r}}}}}}{{Z + 3K}}} \right)\end{aligned}$

This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
使用SBR模型对洪泽湖当前银鱼捕捞管理方式和依据平衡产量模型制定的新的管理方式进行评估。假定2种方案下银鱼的捕捞死亡系数和自然死亡系数不变, 将模型中的时间步长设置为1周, 即0.0192 year[10], SBR模型公式为[8]:
$S \! B \! R \!\!=\!\! \sum\limits_{t = 0}^{{t_{\max }}} \!{\exp [( \!-\! (F \! \times \!\! {S_t}} \! \times \! {A_t}) \!-\! M) \!\! \times \!\! t] \!\! \times \!\! a \!\! \times \!\! {({L_t})^b} \!\! \times\! \! {G_t}$

式中, tmax表示样本的最大年龄; St和Gt分别表示刀刃式选择假设下的网具和性成熟选择性, 当t<50%性成熟体长时,Gt=0, 反之, Gt=1; 当t<初次开捕年龄时,St=0, 反之, St=1[22]; At对应于t龄时是否处于渔业开捕状态, 若是, 则At=1, 否则At=0。
对于SBR模型中使用的50%性成熟体长, 针对2种银鱼1年生且1次性成熟的特点, 以繁殖期性成熟的最小10尾雌性个体的体长均值Lm表示[23], 将其代入最适生长方程中, 计算对应的性成熟年龄Tm。
2. 结果
2.1 种群结构与生长
共测定了洪泽湖1250尾大银鱼和1818尾太湖新银鱼的体长和体重。大银鱼体长和体重分别为29.0—182.6 mm和0.10—34.79 g; 太湖新银鱼体长和体重分别为20.4—82.7 mm和0.04—3.40 g。银鱼的体长-体重关系方程分别为:
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
![]() 图 2 洪泽湖大银鱼(a)和太湖新银鱼(b)的体长频率分布逐月变化Figure 2. Monthly variations in body length frequency distribution for P. hyalocranius (a) and N. taihuensis (b) in Hongze Lake
图 2 洪泽湖大银鱼(a)和太湖新银鱼(b)的体长频率分布逐月变化Figure 2. Monthly variations in body length frequency distribution for P. hyalocranius (a) and N. taihuensis (b) in Hongze Lake大银鱼一个世代在渔获物中出现的时间从4月到翌年2月, 9月以后大银鱼个体间体长差异逐渐增大、体长范围逐渐增加, 12月后大个体在种群中所占比例下降(图 2a)。世代周期中存在4—6月和8—11月2个快速生长阶段。这2个阶段相邻月份的体长和体重均差异显著(P<0.05)。体长和体重均在11月达到最大值。体长和体重瞬时生长率总体均呈下降趋势, 12月后为负值(图 3、图 4)。
太湖新银鱼一个世代在渔获物中出现的时间从5月到翌年4月, 为春季繁殖群体(春群)的后代; 9月也采集到少量性成熟个体(秋群), 其后代于10月开始出现在渔获物中, 但数量较少(图 2b)。本研究仅考虑春群。太湖新银鱼整个世代周期中持续生长, 无明显的阶段性。5—12月间相邻月份间体长差异显著(P<0.05), 但体重差异不显著(P>0.05)。太湖新银鱼体长和体重瞬时生长率变化较大银鱼平稳, 总体呈下降趋势, 12月后近乎于零(图 3、图 4)。
![]() 图 3 洪泽湖大银鱼和太湖新银鱼(不包括秋群)的体长与其瞬时生长率的逐月变化(误差线代表标准差; *表示两个相邻月份间体长差异显著, P<0.05)Figure 3. Monthly fluctuations in body length and its instantaneous growth rate for P. hyalocranius and N. taihuensis in Hongze Lake (Error bars represent ± standard deviation; * indicates significant difference in body length between two adjacent months, P<0.05)
图 3 洪泽湖大银鱼和太湖新银鱼(不包括秋群)的体长与其瞬时生长率的逐月变化(误差线代表标准差; *表示两个相邻月份间体长差异显著, P<0.05)Figure 3. Monthly fluctuations in body length and its instantaneous growth rate for P. hyalocranius and N. taihuensis in Hongze Lake (Error bars represent ± standard deviation; * indicates significant difference in body length between two adjacent months, P<0.05)![]() 图 4 洪泽湖大银鱼和太湖新银鱼(不包括秋群)的体重与其瞬时生长率的逐月变化(误差线代表标准差; *表示两个相邻月份间体重差异显著, P<0.05)Figure 4. Monthly fluctuations in body weight and its instantaneous growth rate for P. hyalocranius and N. taihuensis in Hongze Lake (Error bars represent ± standard deviation; * indicates significant difference in body weight between two adjacent months, P<0.05)
图 4 洪泽湖大银鱼和太湖新银鱼(不包括秋群)的体重与其瞬时生长率的逐月变化(误差线代表标准差; *表示两个相邻月份间体重差异显著, P<0.05)Figure 4. Monthly fluctuations in body weight and its instantaneous growth rate for P. hyalocranius and N. taihuensis in Hongze Lake (Error bars represent ± standard deviation; * indicates significant difference in body weight between two adjacent months, P<0.05)2.2 最适生长方程与性成熟大小
在3种生长方程中, VBGF对大银鱼生长拟合的AIC值最小, Logistic GF对太湖新银鱼生长拟合的AIC值最小(表 1), 分别为2种鱼类的最适生长方程(图 5)。大银鱼和太湖新银鱼卵巢发育至Ⅳ期的10尾最小个体的体长均值Lm分别为99.9和61.1 mm, 代入对应的生长方程, 得出大银鱼和太湖新银鱼的Tm分别为0.528 year和0.564 year。
表 1 三种生长方程对大银鱼和太湖新银鱼生长拟合的方程参数Table 1. Estimated parameters of three different growth functions for P. hyalocranius and N. taihuensis in Hongze Lake参数Parameter 大银鱼P. hyalocranius 太湖新银鱼N. taihuensis Logistic GF Gompertz GF VBGF Logistic GF Gompertz GF VBGF L∞ (mm) 145.86 153.63 173.35 66.82 67.24 67.87 K 5.013 3.536 1.972 5.386 4.569 3.748 t0 (year) 0.376 0.290 0.092 0.124 0.054 –0.042 AIC 7237.47 7220.83 7202.66 12424.96 12428.04 12432.73 ![]() 图 5 洪泽湖大银鱼(a)和太湖新银鱼(b)的最适生长方程曲线Figure 5. The most fitted growth functions for P. hyalocranius (a) and N. taihuensis (b) in Hongze Lake
图 5 洪泽湖大银鱼(a)和太湖新银鱼(b)的最适生长方程曲线Figure 5. The most fitted growth functions for P. hyalocranius (a) and N. taihuensis (b) in Hongze Lake2.3 CPUE逐月变动与种群死亡系数
单位水域面积(1 km2)中大银鱼和太湖新银鱼渔获物数量均在5月最高, 分别为31809和78098 ind./km2。经历了5月中旬为期1周的捕捞后, 6月2种银鱼的资源数量均明显下降; 6月至8月间下降相对平缓; 8月捕捞开始后下降明显, 到12月时渔获物数量均降至500 ind./km2以下。单位水域面积(1 km2)大银鱼和太湖新银鱼渔获物生物量的最高值分别出现在6月和8月, 为73.07和49.33 kg/km2。大银鱼生物量在8—9月之间下降最快,太湖新银鱼生物量在9—10月间下降最快。
根据洪泽湖现行的捕捞管理方式, 5—6月间和8月以后, 2种银鱼的CPUE下降受捕捞和自然死亡的共同影响; 6—8月间, CPUE的下降只受自然死亡的影响。据此求出大银鱼F和M分别为8.583/year和3.292/year,太湖新银鱼F和M分别为7.006/year和1.146/year。
![]() 图 6 洪泽湖大银鱼和太湖新银鱼(春群)单位水域面积(1 km2)渔获物数量(a)和生物量(b)的逐月变动4月之前2种银鱼的个体小、渔获数量少, 均未计算CPUEFigure 6. Monthly variations in number of individuals (a) and biomass (b) for P. hyalocranius and N. taihuensis harvested across an area of 1 km2 by trawl in Hongze LakeIn April, size of both species was small, and numbers of catch for both species was very low. Thus, data in April was not included
图 6 洪泽湖大银鱼和太湖新银鱼(春群)单位水域面积(1 km2)渔获物数量(a)和生物量(b)的逐月变动4月之前2种银鱼的个体小、渔获数量少, 均未计算CPUEFigure 6. Monthly variations in number of individuals (a) and biomass (b) for P. hyalocranius and N. taihuensis harvested across an area of 1 km2 by trawl in Hongze LakeIn April, size of both species was small, and numbers of catch for both species was very low. Thus, data in April was not included2.4 平衡产量模型分析
使用以上获得的生长、繁殖和死亡等相关参数(表 2), 分别构建了大银鱼和太湖新银鱼单位补充量渔产量随开捕年龄(图 6a)和捕捞死亡系数(图 6b)变化的曲线。结果显示, 大银鱼和太湖新银鱼的单位补充量渔产量均随开捕年龄的增大呈先升后降的倒U型曲线关系。当大银鱼开捕年龄tc=0.525 year时, 能够取得相应最大的单位补充量渔产量Ymax=1.078 kg, 开捕年龄过早或过晚均不能获得足够的渔产量。就洪泽湖当前渔业管理模式来说, 大银鱼的开捕年龄约为0.538 year, 略大于tc, 此时的单位补充量渔产量Y=1.077 kg, 接近Ymax。当太湖新银鱼开捕年龄tc=0.525 year时, 能够取得相应最大的单位补充量渔产量Ymax=0.684 kg。就当前渔业管理模式来说, 太湖新银鱼的开捕年龄约为0.365 year, 此时的单位补充量渔产量Y=0.607 kg<Ymax。将开捕时间向后推移20d (即8月29日), 即大银鱼和太湖新银鱼的开捕年龄分别为0.593 year和0.420 year时, 能够获得最大的总单位补充量渔产量1.708 kg。
![]() 图 7 平衡产量模型估算的洪泽湖大银鱼和太湖新银鱼单位补充量渔产量随开捕年龄(a)和捕捞死亡系数(b)变化的曲线垂直虚线表示当前捕捞管理模式下2种银鱼的开捕年龄和捕捞死亡系数Figure 7. Relationships of yield per-recruitment to the age at first capture (a) and fishing mortality coefficient (b) estimated from the equilibrium output mode for P. hyalocranius and N. taihuensis in Hongze LakeThe dotted vertical lines locate the current age at first capture (a) and fishing mortality (b) for both species
图 7 平衡产量模型估算的洪泽湖大银鱼和太湖新银鱼单位补充量渔产量随开捕年龄(a)和捕捞死亡系数(b)变化的曲线垂直虚线表示当前捕捞管理模式下2种银鱼的开捕年龄和捕捞死亡系数Figure 7. Relationships of yield per-recruitment to the age at first capture (a) and fishing mortality coefficient (b) estimated from the equilibrium output mode for P. hyalocranius and N. taihuensis in Hongze LakeThe dotted vertical lines locate the current age at first capture (a) and fishing mortality (b) for both species大银鱼和太湖新银鱼的单位补充量渔产量均先随捕捞死亡系数的增加而快速增高, 当捕捞死亡系数增大到一定程度后, 渔产量上升速度减慢, 之后不再升高甚至出现下降趋势。大银鱼捕捞死亡系数F=24.38/year时, 获得最大单位补充量渔产量Ymax=1.114 kg; 当前洪泽湖大银鱼捕捞死亡系数为8.583/year, 相应渔产量Y=1.077 kg, 接近Ymax且尚未开发过度。太湖新银鱼捕捞死亡系数F=4.72/year时, 获得最大的单位补充量渔产量Ymax=0.616 kg; 而当前洪泽湖太湖新银鱼捕捞死亡系数为7.006/year, 已大于最适捕捞强度, 且相应的渔产量Y=0.607 kg, 小于Ymax。
2.5 基于SBR模型分析的资源状况评价
使用以上获得的生长、繁殖和死亡等相关参数(表 2), 分别构建了2种不同渔业管理方案下大银鱼和太湖新银鱼种群的SBR随捕捞死亡系数变动的曲线:
方案1: 洪泽湖现行的渔业管理方式。8月9日—12月31日为捕捞期, 同时5月可对银鱼资源进行为期1周的捕捞。
方案2: 假定对两种银鱼的捕捞强度不发生变化(即F恒定), 将开捕时间向后推迟20d, 8月29日—12月31日为捕捞期, 同时取消5月为期1周的捕捞。
大银鱼在当前渔业管理方案1下, 相对于未开发状态(100%), SBR残存量为20.234%, 即3.326 g, 接近于补充型过度捕捞评价阈值20%, 种群有可能因产卵亲体不足而导致资源衰退; 在方案2下, SBR残存量相当于未开发状态的36.721%, 即6.036 g, 远大于补充型过度捕捞评价阈值20%, 大银鱼渔业资源和产卵群体得到了较好的保护(图 7)。在2种不同管理方案下, 太湖新银鱼SBR残存量分别相当于未开发状态的7.503%和12.862%, 即0.972 g和1.666 g (图 8)。在优化的管理方案2下, 太湖新银鱼的产卵亲体资源虽然得到了一定程度的保护, 但其渔业资源仍处于补充型过度捕捞水平。
表 2 洪泽湖大银鱼和太湖新银鱼平衡产量模型和单位补充量产卵群体生物量模型参数Table 2. Growth and mortality parameters fitted into the equilib-rium output model and SBR model for P. hyalocranius and N. taihuensis in Hongze Lake参数
Parameter大银鱼
P. hyalocranius太湖新银鱼
N. taihuensisa 1.082×10–6 2.113×10–6 b 3.279 3.215 K 1.972 5.386 L∞ (mm) 173.35 66.82 W∞ (g) 23.75 1.56 t0 (year) 0.092 0.124 Z (/year) 11.875 8.152 M (/year) 3.292 1.146 F (/year) 8.583 7.006 tmax (year) 1 1 Lm (mm) 99.9 61.1 Tm (year) 0.528 0.564 tc (year) 0.538 0.365 ![]() 图 8 两种不同捕捞管理方案下洪泽湖大银鱼(a)和太湖新银鱼(b)单位补充量产卵群体生物量随捕捞死亡系数F变化的曲线当F=0时, 大银鱼和太湖新银鱼的单位补充量产卵群体生物量分别为16.44 g和12.95 g。捕捞管理方案1: 洪泽湖现行的渔业管理方式, 即8月9日—12月31日为捕捞期, 同时5月可对银鱼资源进行为期1周的捕捞; 捕捞管理方案2: 假定对两种银鱼的捕捞强度不变, 将开捕时间向后推迟20d, 即8月29日—12月31日为捕捞期, 同时取消5月为期1周的捕捞。虚线示两种管理方案下对应的捕捞死亡系数Figure 8. Relationships of spawner biomass per-recruitment to fishing mortality coefficients for P. hyalocranius (a) and N. taihuensis (b) under two different fishery management schemes in Hongze LakeThe spawner biomass per-recruitment at F=0 is 16.44 g for P. hyalocranius and 12.95 g for N. taihuensis. Fishery management scheme 1 is the current status, with fishing season of one week in Mid-May and between August 9 and December 31; the scheme 2 is an adjusted status, with fishing season between August 29 and December 31. The dotted vertical lines represent the corresponding fishing mortality coefficients under these two fishery management schemes
图 8 两种不同捕捞管理方案下洪泽湖大银鱼(a)和太湖新银鱼(b)单位补充量产卵群体生物量随捕捞死亡系数F变化的曲线当F=0时, 大银鱼和太湖新银鱼的单位补充量产卵群体生物量分别为16.44 g和12.95 g。捕捞管理方案1: 洪泽湖现行的渔业管理方式, 即8月9日—12月31日为捕捞期, 同时5月可对银鱼资源进行为期1周的捕捞; 捕捞管理方案2: 假定对两种银鱼的捕捞强度不变, 将开捕时间向后推迟20d, 即8月29日—12月31日为捕捞期, 同时取消5月为期1周的捕捞。虚线示两种管理方案下对应的捕捞死亡系数Figure 8. Relationships of spawner biomass per-recruitment to fishing mortality coefficients for P. hyalocranius (a) and N. taihuensis (b) under two different fishery management schemes in Hongze LakeThe spawner biomass per-recruitment at F=0 is 16.44 g for P. hyalocranius and 12.95 g for N. taihuensis. Fishery management scheme 1 is the current status, with fishing season of one week in Mid-May and between August 9 and December 31; the scheme 2 is an adjusted status, with fishing season between August 29 and December 31. The dotted vertical lines represent the corresponding fishing mortality coefficients under these two fishery management schemes3. 讨论
3.1 大银鱼和太湖新银鱼的阶段性生长差异
鱼类的生长具有阶段性, 生活史的不同阶段, 个体的生理特征、食性以及环境因素等差异, 会引起生长速度的改变[24, 25]。本研究发现, 洪泽湖大银鱼存在2个快速生长阶段, 分别在4—6月和8—11月间。鱼类一般在早期生活史阶段和性成熟前体长生长较快[26], 这一阶段的快速生长可能是鱼类维持种群数量的一种生态适应方式, 有利于降低其被捕食的压力、快速达到性成熟体长[27, 28]。洪泽湖大银鱼繁殖期在12—2月(作者未发表数据), 4—6月间的第一个快速生长阶段, 在个体性成熟之前; 同时, 该时期水体温度逐步上升, 初级生产力升高, 水体中浮游动物等大银鱼的主要饵料生物丰富, 是这一阶段大银鱼快速生长的营养保障[29]。8—11月间第二阶段快速生长可能主要源于大银鱼的食性转化。唐富江发现兴凯湖大银鱼的体长大于60 mm以后, 逐渐由摄食浮游动物转为摄食鱼虾类[30]; 本文作者也发现, 自8月起虾类和鱼类逐渐取代枝角类和桡足类成为洪泽湖大银鱼的主要食物(作者未发表数据)。鱼虾类相对于浮游动物而言具有体积大、能量高、动物蛋白丰富等特点, 更易于被鱼类吸收和利用, 作为生长和繁殖的能量供应[31]。因此, 大银鱼在8—11月的快速生长很可能是因为有更多的能量来源投入到生长中。12月后, 大银鱼种群体长和体重均表现为负增长, 这可能与其进入繁殖期后大个体较早的繁殖和死亡有关[32], 需进一步研究确证。
太湖新银鱼个体相对较小, 其体长在5—10月呈快速持续生长状态, 无明显阶段性。太湖新银鱼终生摄食浮游动物, 与大银鱼不同, 没有明显的食性转化[33]。太湖新银鱼这一食性特点可能是其生长过程没有明显阶段性的主要原因。11月以后, 太湖新银鱼个体体长无明显的增长趋势。洪泽湖太湖新银鱼繁殖季节主要在3—4月, 11月之后, 繁殖季节前, 是性腺发育能量积累的主要阶段[34]。因此, 能量投入到性腺发育中, 可能是太湖新银鱼11月后体长停止增长的重要原因。
3.2 洪泽湖银鱼资源利用评估与管理
鱼类种群可提供渔获量的多少取决于种群数量、生长、存活、开捕年龄和捕捞强度等, 其中仅有捕捞强度(捕捞死亡系数)和初次捕捞年龄(初次开捕体长)能够被人为控制, 因此渔业管理中常通过调整这2个因素以获取最大可持续渔产量[6]。依据平衡产量模型分析, 洪泽湖大银鱼和太湖新银鱼在当前捕捞强度下的渔产量均已接近其资源群体所能提供的最大渔产量, 调整捕捞强度对进一步提高渔产量的影响不大; 而调整2种银鱼的初次开捕年龄, 更有利于获得较高的总渔产量。
在洪泽湖当前的捕捞管理方式下, 大银鱼和太湖新银鱼的SBR残存量分别为未开发状态的20.23%和7.50%, 表明对2种银鱼的捕捞均处于补充型过度捕捞水平, 且太湖新银鱼捕捞后的SBR残余量已远低于种群崩溃的警戒线。由此看来, 较大的捕捞强度和补充型过度捕捞, 可能是导致近年来洪泽湖大银鱼和太湖新银鱼资源波动明显的重要原因。因此, 很有必要对洪泽湖当前银鱼资源捕捞管理方式进行优化, 避免过度捕捞的发生。
本研究构建平衡产量模型时发现, 将洪泽湖银鱼开捕时间推迟20d可获得大银鱼和太湖新银鱼的最大总渔产量。在渔业管理中, 这种开捕时间的推迟也将影响最终捕捞后的产卵亲体生物量[8, 9]。对洪泽湖的银鱼捕捞管理方式进行优化后, 大银鱼的SBR残存量提高至未开发状态的36.72%, 既满足了获得最高渔产量的需求, 又不会带来太大的渔业风险; 优化管理后太湖新银鱼的SBR残存量亦提高至未开发状态的12.86%。可见, 优化的渔业管理方式实现了获取最高渔产量和保护产卵亲体资源上的“双赢”。同时也应注意到, 优化的渔业管理方案对大银鱼产卵群体的保护作用更为显著; 在未改变当前捕捞强度的情况下, 尽管太湖新银鱼在捕捞后的产卵亲体资源量也得到了一定程度的提升, 但依然低于补充型过度捕捞评价阈值20%, 故还需减小对太湖新银鱼的捕捞强度。因此, 针对洪泽湖太湖新银鱼的渔业资源管理, 在延迟开捕时间的基础上, 建议还可以通过限制渔获物的起捕体长、增大捕捞网具的网目规格等方式控制捕捞强度, 同时可采取人工放流等措施提高其下一代的资源补充能力, 以避免种群资源的进一步衰退。
-
![]()
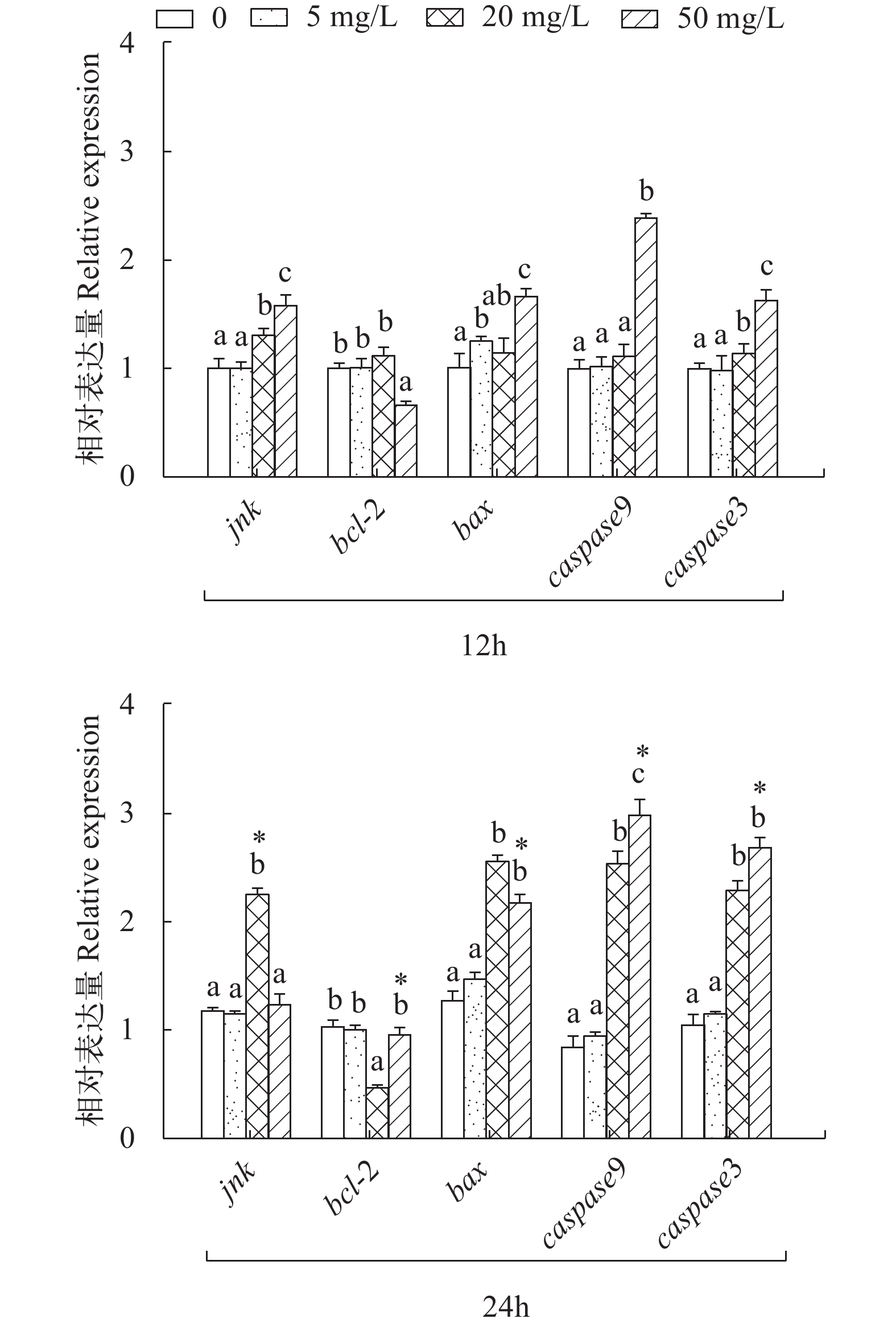
图 1 亚硝酸钠对L8824细胞jnk、bcl-2、bax、caspase9和caspase3表达量的影响
不同字母表示同一时间点组内具有显著性差异(P<0.05), *表示不同时间点组间具有显著性差异(P<0.05), 下同
Figure 1. Effects of sodium nitrite on the expression of jnk, bcl-2, bax, caspase9 and caspase3 in L8824 cells
Different letters indicates significant differences between groups at the same time point (P<0.05), * indicates significant differences between groups at different time points (P<0.05), the same applies below
![]()
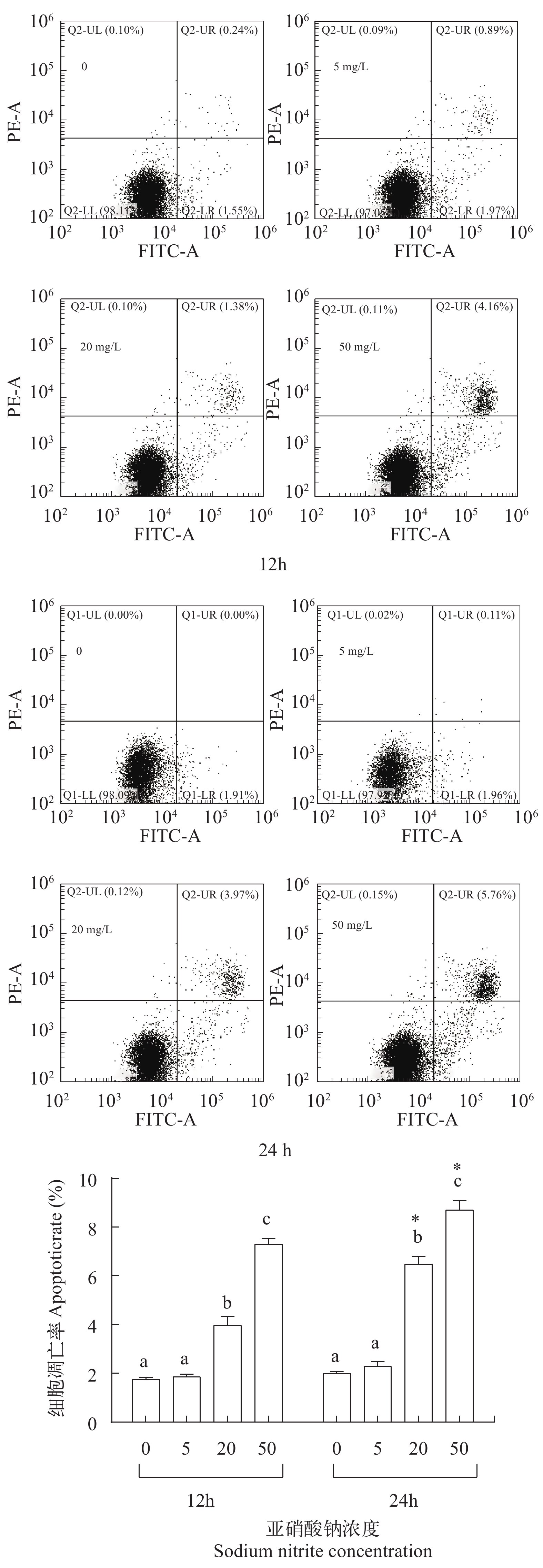
图 2 亚硝酸钠对L8824细胞凋亡的影响
Figure 2. The effect of sodium nitrite on apoptosis of L8824 cells
![]()
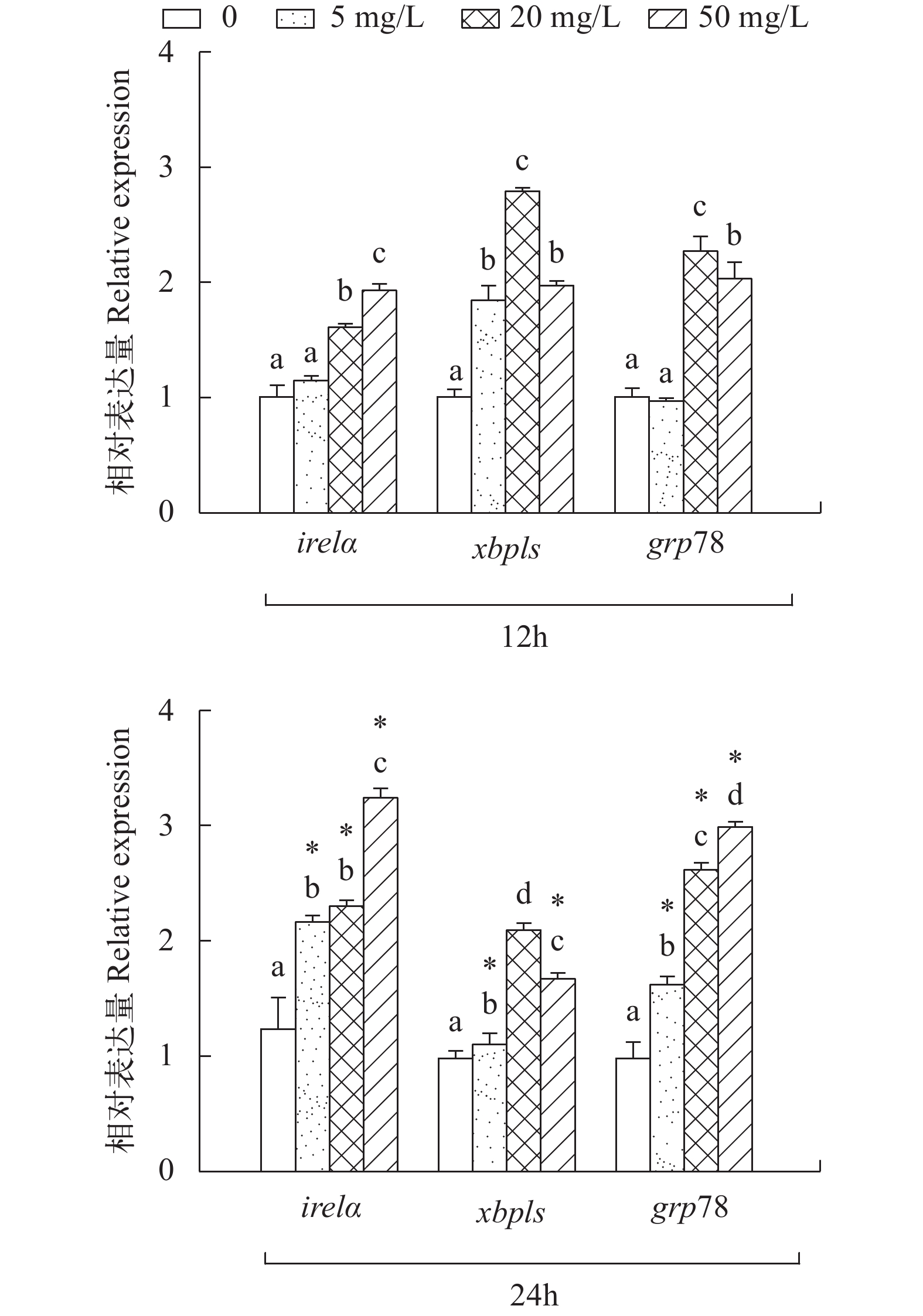
图 3 亚硝酸钠对L8824细胞ire1α, xbp1s和grp78表达量的影响
Figure 3. Effects of sodium nitrite on the expression of ire1α, xbp1s and grp78 in L8824 cells
![]()
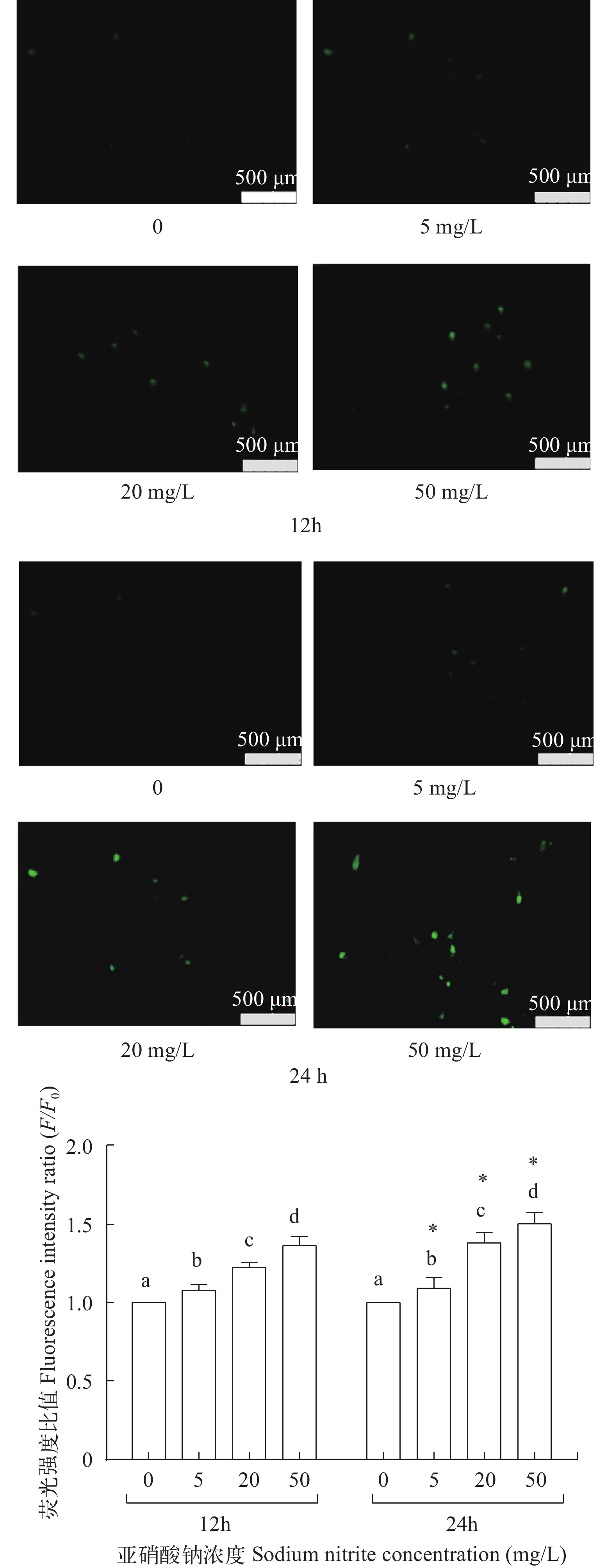
图 4 亚硝酸钠对L8824细胞内钙离子浓度的影响
Figure 4. The effect of sodium nitrite on the intracellular calcium content in L8824 cells
![]()
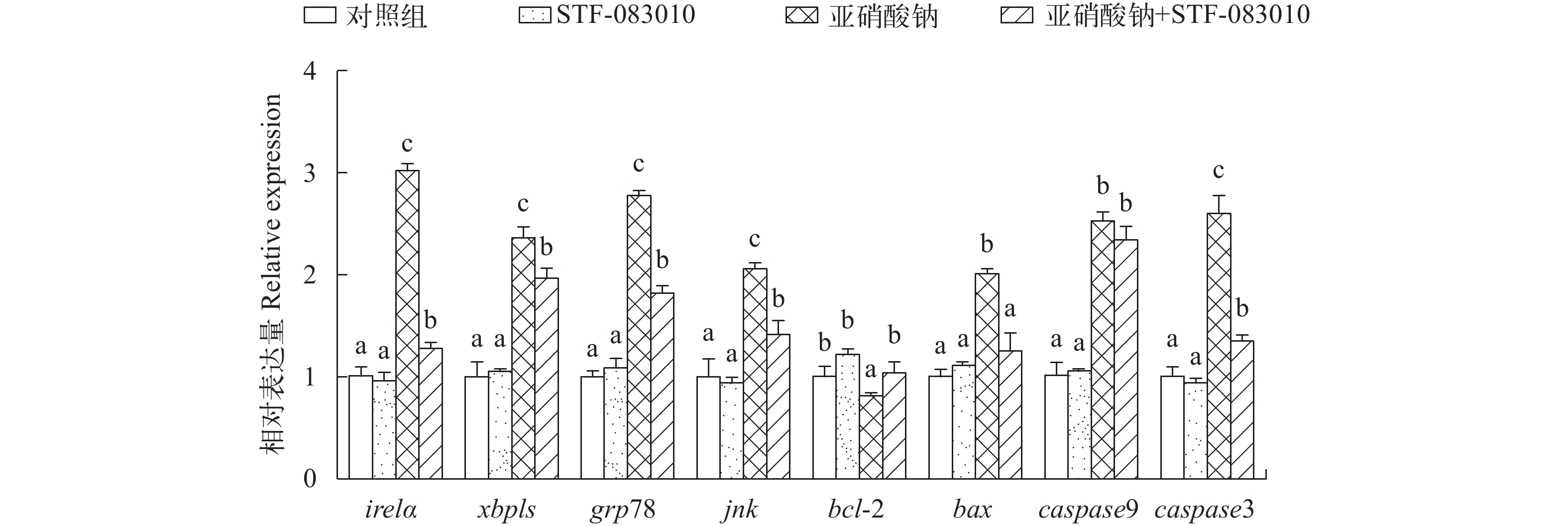
图 5 亚硝酸钠和STF-083010及其配伍对L8824细胞ire1α、xbp1s、grp78、jnk、bcl-2、bax、caspase9和caspase3表达量的影响
Figure 5. Effects of sodium nitrite, STF-083010 and their combination on the expression of ire1α, xbp1s, grp78, jnk, bcl-2, bax, caspase9 and caspase3 in L8824 cells
![]()
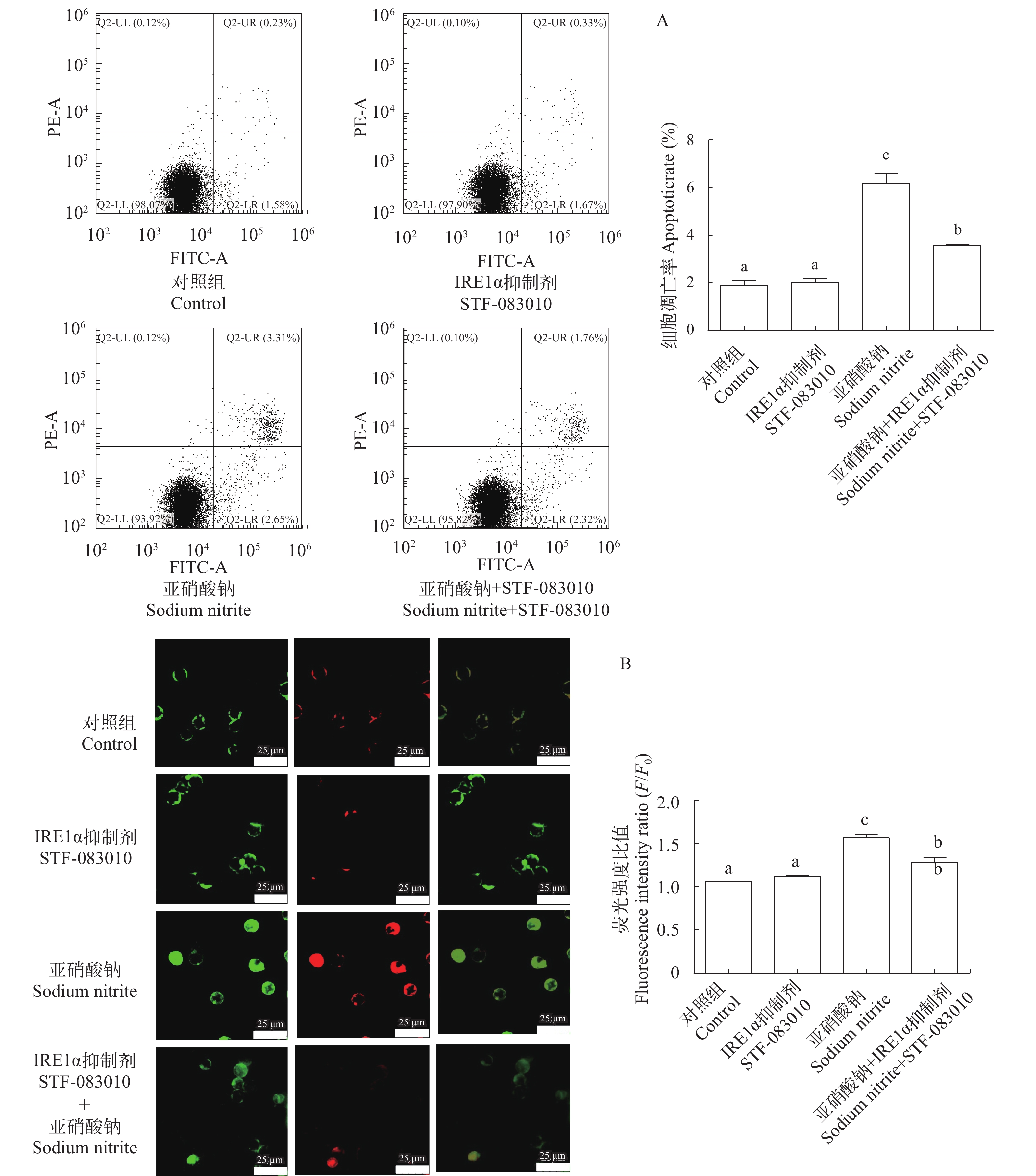
图 6 亚硝酸钠和STF-083010及其配伍对L8824细胞凋亡的影响
A. 流式Annexin-FITC/PI 双染法检测L8824细胞凋亡; B. 流式Annexin-FITC/PI 双染法在荧光显微镜下检测L8824细胞凋亡, Annexin-FITC染色为绿色荧光, PI染色为红色荧光
Figure 6. Effects of sodium nitrite, STF-083010 and their combination on the cell apoptosis of L8824 cells
A. Detection of apoptosis in L8824 cells by flow Annexin-FITC/PI double staining; B. Annexin-FITC/PI double staining was used to detect apoptosis of L8824 cells under fluorescence microscope, Annexin-FITC staining was green fluorescence and PI staining was red fluorescence
![]()
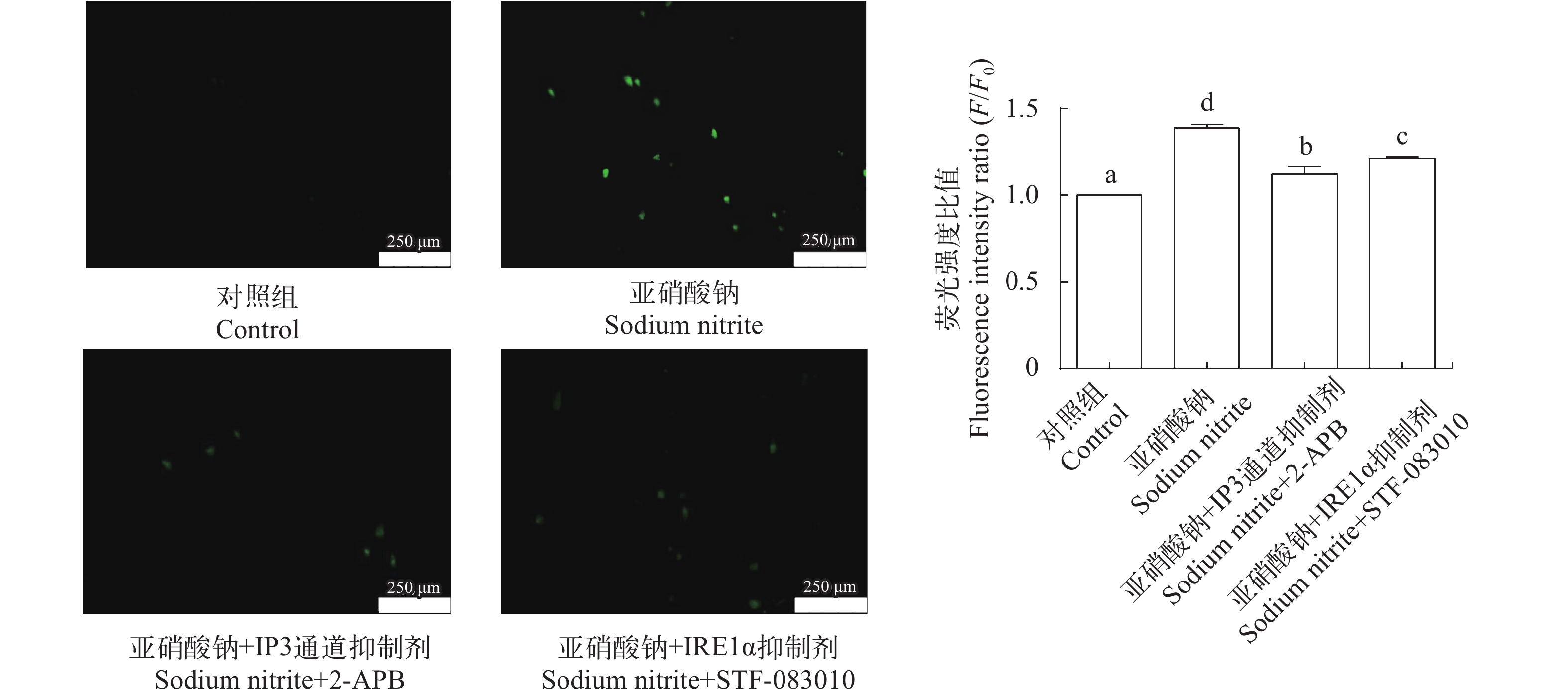
图 7 2-APB和STF-083010 对亚硝酸钠暴露L8824细胞内钙离子浓度的影响
Figure 7. Effects of 2-APB and STF-083010 on the intracellular calcium content under sodium nitrite exposure in L8824 cells
表 1 引物序列
Table 1 Primers used in the study
目的基因Target gene 引物序列Primer sequences (5′-3′) 作物长度Product length (bp) jnk F: ACAGCGTAGATGTGGGTGATT
R: GCTCAAGGTTGTGGTCATACG108 bcl-2 F: CCACCCATTGTCATACCTAACG
R: ACCCACA GGGCTGTCACTTCT98 bax F: GGATCAGTTGGGTGGCGTTGA
R: TGAGTCGGTTGAAGAGCAGAGT104 caspase9 F: GGGATAGATGACCAGATGGA
R: GGGATAGATGACCAGATGGA164 caspase3 F: GGCAGATGGTGACAGTGATGGA
R: TGGCAGATTGGAGGAGGATTCG154 ire1α F: GAACGCCACATACTCTGA
R: TGTCCACTGTCACCACTA146 xbp1s F: AGGGGAAGTGAATACGGGGAAT
R: GACCTCACAGAAGGCAACGGAATC102 grp78 F: GGTGCTTAATTCTGCGTTAT
R: CCTTCTTCTCGTCGTCTTC92 β-actin F: CACTGTGCCCATCTACGA
R: CCATCTCCTGCTCGAAGTC80  下载: 导出CSV
下载: 导出CSV
表 2 不同浓度亚硝酸钠对L8824 细胞活力的影响
Table 2 The effect of sodium nitrite on the viability of L8824 cells
亚硝酸钠浓度Sodium nitrite concentration (mg/L) 细胞活力Cell viability(%) 12h 24h 0 100.00±0.00c 100.00±0.00c 5 98.44±1.46bc 97.65±1.94c 20 97.68±0.47b 93.67±0.83b 50 90.05±1.74a 87.44±0.76a 注: 同列上标小写字母不同表示二者之间差异显著(P<0.05); 下同
Note: Values in the same row with different superscripts are significantly different (P<0.05); the same applies below
下载: 导出CSV
表 3 亚硝酸钠和STF-083010共同作用对L8824细胞活力的影响
Table 3 Effects of sodium nitrite and STF-083010 on the viability of L8824 cells
组别 Group 细胞活力 Cell viability (%) 对照组 Control 100.00±0.00c IRE1α抑制剂 STF-083010 99.18±0.52c 亚硝酸钠 Sodium nitrite 92.64±1.01a IRE1α抑制剂+亚硝酸钠 STF-083010+Sodium nitrite 96.31±1.38b
下载: 导出CSV
-
[1] Jensen F B. Nitrite disrupts multiple physiological functions in aquatic animals [J]. Comparative Biochemistry and Physiology Part A Molecular and Integrative Physiology, 2003, 135(1): 9-24. doi: 10.1016/S1095-6433(02)00323-9
[2] 余瑞兰, 聂湘平, 魏泰莉, 等. 分子氨和亚硝酸盐对鱼类的危害及其对策 [J]. 中国水产科学, 1999, 6(3): 73-77. doi: 10.3321/j.issn:1005-8737.1999.03.017 Yu R L, Nie X P, Wei T L, et al. Toxicity of molecular ammonia and nitrite to fishes and the control measures [J]. Journal of Fishery Sciences of China, 1999, 6(3): 73-77. doi: 10.3321/j.issn:1005-8737.1999.03.017
[3] 王鸿泰, 胡德高. 池塘中亚硝酸盐对草鱼种的毒害及防治 [J]. 水产学报, 1989, 13(3): 207-214. Wang H T, Hu D G. Toxicity of nitrite to Ctenopharyngodon idella in ponds and its way of prevention [J]. Journal of Fisheries of China, 1989, 13(3): 207-214.
[4] Sun S, Ge X, Xuan F, et al. Nitrite-induced hepatotoxicity in Bluntsnout bream (Megalobrama amblycephala): The mechanistic insight from transcriptome to physiology analysis [J]. Environmental Toxicology and Pharmacology, 2014, 37(1): 55-65. doi: 10.1016/j.etap.2013.11.010
[5] Marcio Jose dos Santos Silva, Leme F P, Takata R, et al. Biological responses of Neotropical freshwater fish Lophiosilurus alexandri exposed to ammonia and nitrite [J]. Science of the Total Environment, 2018(616-617): 1566-1575.
[6] Svobodova Z, Machova J, Poleszczuk G, et al. Nitrite poisoning of fish in aquaculture facilities with water-recirculating systems [J]. Acta Veterinaria Brno, 2005, 74(1): 9584-9585.
[7] Wang W N, Wang A L, Zhang Y J, et al. Effects of nitrite on lethal and immune response of Macrobrachium nipponense [J]. Aquaculture, 2004, 232(1-4): 679-686. doi: 10.1016/j.aquaculture.2003.08.018
[8] Fu L Z, Yong Q L, Qi S, et al. Effects of sodium nitrite on proliferation and apoptosis on human hepatocarcinoma cells [J]. Chinese Pharmacological Bulletin, 2011, 27(2): 191-195.
[9] 屈三甫, 张小榕, 郑从义, 等. 鱼呼肠孤病毒诱导草鱼肾细胞凋亡 [J]. 水生生物学报, 2000, 24(6): 616-620. Qu S P, Zhang X R, Zheng C Y, et al. Apoptosis of the cell line from grass carp (CIK) induced by fish reovirus [J]. Acta Hydrobiologica Sinica, 2000, 24(6): 616-620.
[10] Walter P, Ron D. The unfolded protein response: from stress pathway to homeostatic regulation [J]. Science, 2011, 334(6059): 1081-1086. doi: 10.1126/science.1209038
[11] Xiang C, Wang Y, Zhang H, et al. The role of endoplasmic reticulum stress in neurodegenerative disease [J]. Apoptosis, 2017, 22(1): 1-26. doi: 10.1007/s10495-016-1296-4
[12] Xu Z, Chikka M R, Xia H, et al. Ire1 supports normal ER differentiation in developing Drosophila photore ceptors [J]. Journal of Cell Science, 2016, 129(5): 921-929. doi: 10.1242/jcs.180406
[13] Cao S S, Kaufman R J. Endoplasmic reticulum stress and oxidative stress in cell fate decision and human disease [J]. Antioxidants and Redox Signaling, 2014, 21(3): 396-413. doi: 10.1089/ars.2014.5851
[14] Schroder M. Endoplasmic reticulum stress responses [J]. Cellular and Molecular Life Sciences, 2008, 65(6): 862-894. doi: 10.1007/s00018-007-7383-5
[15] Liu X H, Zhang Z Y, Sun S, et al. Ischemic postconditioning protects myocardium from ische-mia/reperfusion injury through attenuating endoplasmic reticulum stress [J]. Shock, 2008, 30(4): 422-427. doi: 10.1097/SHK.0b013e318164ca29
[16] Long M, Lin W, Hou J, et al. Dietary supplementation with selenium yeast and tea polyphenols improve growth performance and nitrite tolerance of Wuchang bream (Megalobrama amblycephala) [J]. Fish and Shellfish Immunology, 2017(68): 74-83. doi: 10.1016/j.fsi.2017.07.017
[17] 李雪贤, 孙健, 吉红, 等. 脂肪酸影响草鱼肝细胞脂质蓄积状态及诱导其凋亡的离体研究 [J]. 水生生物学报, 2017, 41(1): 56-64. doi: 10.7541/2017.8 Li X X, Su J, Ji H, et al. Influence of fatty acids on lipid accumulation and apoptosis status of grass carp Ctenopharyngodon idellus hepatocyte in vitro [J]. Acta Hydrobiologica Sinica, 2017, 41(1): 56-64. doi: 10.7541/2017.8
[18] 萧培珍, 吉红, 张宝彤, 等. 水飞蓟素抑制草鱼肝细胞脂质蓄积的作用及其机制研究 [J]. 水生生物学报, 2017, 41(6): 137-146. Xiao P Z, Ji H, Zhang B T, et al. Inhibitory effect of silymarin on oleic acid-induced lipid accumulation in grass carp (Ctenopharyngodon idellus) hepatocyte in vitro [J]. Acta Hydrobiologica Sinica, 2017, 41(6): 137-146.
[19] Papandreou I, Denko N C, Olson M, et al. Identification of an Ire1alpha endonuclease specific inhibitor with cytotoxic activity against human multiple myeloma [J]. Blood, 2011, 117(4): 1311-1314. doi: 10.1182/blood-2010-08-303099
[20] Rodrigues A D S L, Gnocchi K G, Medeiros L C C, et al. Acute toxicity of nitrite in fat snook (Centropomus parallelus) [J]. Marine and Freshwater Behaviour and Physiology, 2018, 51(2): 1-13.
[21] Le T H G, Jensen F B, Damsgaard C, et al. Extreme nitrite tolerance in the clown knifefish Chitala ornata is linked to up-regulation of methaemoglobin reductase activity [J]. Aquatic Toxicology, 2017, (187): 9-17.
[22] Evans D H, Piermarini P M, Potts WTW. Ionic transport in the fish gill epithelium [J]. Journal of Experimental Zoology, 1999, 283(7): 641-652. doi: 10.1002/(SICI)1097-010X(19990601)283:7<641::AID-JEZ3>3.0.CO;2-W
[23] Onken H, Putzenlechner M. A V-ATPase drives active, electrogenic and Na+-independent Cl- absorption across the gills of Eriocheir sinensis [J]. Journal of Experimental Biology, 1995, 198(3): 767-774.
[24] Bath R N, Eddy F B. Transport of nitrite across fish gills [J]. Journal of Experimental Zoology, 1980, 214(1): 119-121. doi: 10.1002/jez.1402140115
[25] Krous S R, Blazer V S, Meade T L. Effect of acclimation time on nitrite movement across the gill epithelia of rainbow trout: the role of “chloride cells” [J]. Progressive Fish Culturist, 1982, 44(3): 126-130. doi: 10.1577/1548-8659(1982)44[126:EOATON]2.0.CO;2
[26] Campos M L, Barbas Luis André Luz, Fiori N L, et al. Oxidative stress and antioxidant responses in juvenile Brazilian flounder Paralichthys orbignyanus exposed to sublethal levels of nitrite [J]. Fish Physiology and Biochemistry, 2018(44): 1349-1362.
[27] 强俊, 徐跑, 何杰, 等. 氨氮与拥挤胁迫对吉富品系尼罗罗非鱼幼鱼生长和肝脏抗氧化指标的联合影响 [J]. 水产学报, 2011, 35(12): 1837-1848. Qiang J, Xu P, He J, et al. The combined effects of ammonia nitrogen and crowding stress on the growth and liver antioxidant index of the Nile tilapia juveniles [J]. Journal of Fisheries, 2011, 35(12): 1837-1848.
[28] Doblander C, Lackner R. Metabolism and detoxification of nitrite by trout hepatocytes [J]. Biochimica et Biophysica Acta, 1996, 1289(2): 270-274. doi: 10.1016/0304-4165(95)00166-2
[29] Michael M I, Hilmy A M, El-Domiaty N A, et al. Serum transminases activity and histopathological changes in Clarias lazera chronically exposed to nitrite [J]. Comparative Biochemistry and Physiology Part C Comparative Pharmacology, 1987, 86(2): 255-262.
[30] Xian J A, Wang A L, Hao X M, et al. In vitro toxicity of nitrite on haemocytes of the tiger shrimp, Penaeus monodon, using flow cytometric analysis [J]. Comparative Biochemistry and Physiology Part C: Toxicology and Pharmacology, 2012, 156(2): 75-79. doi: 10.1016/j.cbpc.2012.04.001
[31] Al-Gayyar M M, Hassan H M, Alyoussef A, et al. Nigella sativa oil attenuates chronic nephrotoxicity in-duced by oral sodium nitrite: Effects on tissue fibrosis and apoptosis [J]. Redox Report, 2015, 21(2): 50-60.
[32] Al-Rasheed N M, Fadda L M, Attia H A, et al. Quercetin inhibits sodium nitrite-induced inflammation and apoptosis in different rats organs by suppressing Bax, HIF1-α, TGF-β, Smad-2, and AKT pathways [J]. Journal of Biochemical and Molecular Toxicology, 2016, 31(5): 1-7.
[33] Liu C, Huang Y, Zhang Y, et al. Intracellular methylglyoxal induces oxidative damage to pancreatic beta cell line INS-1 cell through Ire1α-JNK and mitochondrial apoptotic pathway [J]. Free Radic Research, 2017, 51(4): 1-14.
[34] 等. 过氧化亚硝酸盐在人血管内皮引起内质网应激和凋亡预示动脉粥样硬化形成 [J]. 中国动脉硬化杂志, 2006, 25(12): 623-629. Dickhout, Hossain, Pozza, et al. Endoplasmic reticulum stress and apoptosis induced by nitrite peroxide in human vascular endothelium predict atherosclerosis [J]. Chinese Journal of Arteriosclerosis, 2006, 25(12): 623-629. [Dickhout, Hossain, Pozza,
[35] Dudek J, Benedix J, Cappel S, et al. Functions and pathologies of BiP and its interaction partners [J]. Cellular and Molecular Life Sciences (CMLS)
, 2009, 66(9): 1556-1569. doi: 10.1007/s00018-009-8745-y [36] Argon Y, Simen B B. GRP94, an ER chaperone with protein and peptide binding properties [J]. Seminars in Cell and Developmental Biology, 1999, 10(5): 495-505. doi: 10.1006/scdb.1999.0320
[37] Lievremont J P, Rizzuto R, Hendershot L, et al. BiP, a major chaperone protein of the endoplasmic reticulum lumen, plays a direct and important role in the storage of the rapidly exchanging pool of Ca2+ [J]. Journal of Biological Chemistry, 1997, 272(49): 30873-30879. doi: 10.1074/jbc.272.49.30873
[38] Ron D, Walter P. Signal integration in the endoplasmic reticulum unfolded protein response [J]. Nature Reviews Molecular Cell Biology, 2007, 8(7): 519-529. doi: 10.1038/nrm2199
[39] Pan Y, Xu J, Wang X, et al. The role of cell apoptosis mediated by endoplasmic reticulum stress (ERS) of deep tissue injury of pressure ulcer of rats [J]. Chinese Journal of Applied Physiology, 2015, 31(5): 396-400.
[40] Kato H, Nakajima S, Saito Y, et al. mTORC1 serves ER stress-triggered apoptosis via selective activation of the IRE1-JNK pathway [J]. Cell Death and Differentiation, 2012, 19(2): 310-320. doi: 10.1038/cdd.2011.98
[41] Huang M, Xu A, Wu X, et al. Japanese encephalitis virus induces apoptosis by the IRE1/JNK pathway of ER stress response in BHK-21 cells [J]. Archives of Virology, 2016, 161(3): 699-703. doi: 10.1007/s00705-015-2715-5
-
期刊类型引用(8)
1. 公莉,过龙根,尹成杰,杨姣姣,杨雅兰. 洱海西太公鱼和太湖新银鱼生长特性及种群调控效果研究. 水生态学杂志. 2022(01): 117-123 .  百度学术
百度学术
2. 李大命,唐晟凯,刘燕山,谷先坤,刘小维,殷稼雯,张彤晴,潘建林. 基于Cytb基因的江苏省大银鱼种群遗传多样性和遗传结构分析. 上海海洋大学学报. 2021(03): 416-425 . 百度学术
3. 赵旭昊,徐东坡,任泷,徐跑. 基于Ecopath模型的太湖鲢鳙生态容量评估. 中国水产科学. 2021(06): 785-795 . 百度学术
4. 王煜恒,陈军,李华. 大银鱼仔鱼的人工培育技术探究. 南方农业. 2021(29): 178-179 . 百度学术
5. 李大命,唐晟凯,刘燕山,谷先坤,刘小维,李强,高军,张伟,穆新武,张彤晴. 骆马湖大银鱼、太湖新银鱼线粒体Cytb和COⅠ基因序列多态性分析. 江苏农业科学. 2020(11): 47-52 . 百度学术
6. 李大命,唐晟凯,刘燕山,谷先坤,刘小维,殷稼雯,张彤晴,潘建林. 江苏省4个太湖新银鱼种群遗传多样性和遗传结构分析. 渔业科学进展. 2020(05): 52-60 . 百度学术
7. 唐富江,高文燕,李慧琴,刘伟. 大银鱼生物学与渔业生态学研究进展. 水产学报. 2020(12): 2100-2111 . 百度学术
8. 毛志刚,谷孝鸿,龚志军,曾庆飞,陈辉辉,李红敏,张胜宇,穆欢. 洪泽湖鱼类群落结构及其资源变化. 湖泊科学. 2019(04): 1109-1119 . 百度学术
其他类型引用(7)








计量
- 文章访问数: 3753
- HTML全文浏览量: 975
- PDF下载量: 84
- 被引次数: 15