
ROLE OF ENDOPLASMIC RETICULUM STRESS IRE1 PATHWAY IN HEPATOCYTE APOPTOSIS OF GRASS CARP CTENOPHARYNGODON IDELLA INDUCED BY SODIUM NITRITE
-
摘要: 为探究亚硝酸钠诱导草鱼肝细凋亡中内质网应激IRE1通路的作用, 草鱼(Ctenopharyngodon idellus)肝细胞L8824分别置于亚硝酸钠浓度为0、5、20和50 mg/L暴露12h和24h。同时采用三磷酸肌醇受体拮抗剂2-APB和IRE1α抑制剂STF-083010分别和20 mg/L亚硝酸钠共同孵育L8824细胞24h。检测细胞凋亡以及c-Jun氨基末端激酶(c-Jun N-terminal kinase, jnk)、B淋巴细胞瘤-2(B-cell lymphoma-2, bcl-2)、bcl-2相关X蛋白(bcl-2 associated X protein, bax)、caspase9、caspase3、肌醇酶1α (inositol requiring enzyme 1α, ire1α)、X盒结合蛋白 1s (X-box binding protein 1s, xbp1s)和葡萄糖调节蛋白78 (glucose-regulated protein78, grp78)基因的表达和细胞质内钙离子浓度。结果表明: 与对照组相比, 亚硝酸钠处理组细胞凋亡率显著上升, jnk、bax、caspase9、caspase3、ire1α、xbp1s和grp78 mRNA的表达显著升高, bcl-2 mRNA的表达量显著下降, 细胞质内钙离子浓度显著上升。与亚硝酸钠单处理组相比, STF-083010处理组细胞凋亡率显著下降, ire1α、xbp1s、grp78、jnk、bax、caspase3 mRNA表达量显著下降, bcl-2 mRNA表达量显著上升, 2-APB和STF-083010两个处理组细胞质内钙离子浓度均显著下降。结果显示, 高浓度的亚硝酸钠可诱导草鱼肝细胞凋亡和钙离子紊乱, 内质网应激IRE1通路发挥了作用。Abstract: Nitrite, a common pollutant in aquaculture, is an intermediate product of nitrogen cycle in ecosystem. To explore the mechanisms of sodium nitrite-induced cell apoptosis, grass carp liver cell (L8824) were exposed to four concentrations of sodium nitrite (0, 5 mg/L, 20 mg/L and 50 mg/L) with or without treatments of phosphoinositide receptor antagonist 2-APB and IRE1 inhibitors STF-083010. Cell apoptosis related gene expression of jnk, bcl-2, bax, caspase9, caspase3, ire1α, xbp1s and grp78 and the cytoplasmic calcium ion concentration were assessed. The results showed that nitrite significantly increased the apoptosis rate, cytoplasm calcium ion concentration and mRNA levels of jnk, bax, caspase9, caspase3, ire1α, xbp1s and grp78 and significantly decreased bcl-2 mRNA level, which were reversed by the STF-083010 treatment. Besides, both 2-APB and STF-083010 reduced the sodium nitrite-induced cytoplasmic calcium ion. These results indicate that endoplasmic reticulum stress-related IRE1 pathway plays pivotal role nitrite-mediated L8824 cell apoptosis and calcium dyshomeostasis.
-
Keywords:
- Sodium nitrite /
- IRE1 pathway /
- Apoptosis /
- Calcium ion
-
年龄鉴定是研究鱼类生物学和生态学特性的重要组成部分, 也是分析和评价鱼类种群数量变动的基本依据之一[1]。鱼类年龄鉴定最普遍的方法是钙化组织分析法[2]。对于裂腹鱼类而言, 受特定生长环境的影响, 不同种类、不同阶段个体的鉴定材料年轮特征各不相同, 呈现出多样化的特点[3]。由于遗传差异和生存环境的差异, 鱼类的生长呈现出不同特点, 不同钙化组织上的年轮特征也表现各异。因此需要对不同的年龄材料进行对比, 最终选出最佳的年龄材料[4]。
双须叶须鱼(Ptychobarbus dipogon)隶属于裂腹鱼亚科(Schizothoracinae), 叶须鱼属(Ptychobarbus), 主要分布在西藏雅鲁藏布江中游干支流砂石底质的缓流处[5]。双须叶须鱼独特的种群特征、生活地理环境决定了其对环境变化和人为干扰极为敏感和脆弱[6]。近年来, 随着水利交通工程建设、渔业捕捞及工程材料索取加剧, 双须叶须鱼的种群数量和种群资源下降, 对双须叶须鱼种群资源构成了威胁。双须叶须鱼作为一个极为脆弱的种群资源, 为保护该物种的可持续性和稳定的种群而做出相应的管理是至关重要的[7]。杨鑫[6]主要研究雅鲁藏布江中游双须叶须鱼的耳石和脊椎骨两种年龄材料, Li等[7]主要研究雅鲁藏布江及其支流流域双须叶须鱼三对耳石年龄材料和微耳石的微结构特征及日增量。而本文研究主要是研究拉萨河流域双须叶须鱼8种年龄材料的年轮特征和年龄鉴定。
年龄鉴定结果可以阐明鱼类生长、性成熟年龄, 直接关系到鱼类种群分析与资源评估现状[8]。因此, 评估和比较每个物种所使用的年龄材料的研究是最基础和必要的内容[9]。本文对双须叶须鱼的微耳石、星耳石、脊椎骨、鳃盖骨、臀鳞、胸鳞、侧线鳞和背鳞等8种年龄材料的年轮特征进行了描述, 并对8种材料鉴定年龄的准确性和精确性进行了比较研究, 旨在为其生长特征、种群动态和资源变动提供可靠的年龄依据。
1. 材料与方法
1.1 样本采集
样本于2014年2月至12月在拉萨河上游采集双须叶须鱼193尾。全部测量全长、体长和体重, 长度精确到1 mm, 重量精确到0.1 g。取出左右微耳石、左右星耳石、鳃盖骨和6—8枚脊椎骨以及4种鳞片。样品用清水清洗, 晾干后放入封口袋冷冻保存。
1.2 年龄材料处理
微耳石 将微耳石远极面朝上, 用指甲油包埋, 固定在载玻片上, 静置, 让其凝固, 然后先用#1500的砂纸打磨, 再用#2000#的砂纸抛光, 打磨期间时刻加水, 并随时在显微镜下观察。直至微耳石核区年轮清晰为止, 最后放在显微镜下拍照保存。
星耳石 将星耳石内侧面朝上, 用指甲油包埋, 固定在载玻片上, 静置, 让其凝固。首先用#1500的砂纸打磨。再用#2000的砂纸抛光, 打磨期间时刻加水, 并在显微镜下拍照保存。
脊椎骨 将脊椎骨放入开水中浸泡1—2min, 用牙刷轻轻刷去附着的肌肉和结缔组织, 剪去多余的骨棘, 从中间将脊椎骨剪断, 静置让其干燥。调整好角度在显微镜下拍照保存。
鳃盖骨 将鳃盖骨放入开水中1min, 用牙刷剔除附着的肌肉的结缔组织, 让其自然干燥, 然后放在体视显微镜下用透射光观察, 并拍照保存。
鳞片 将鳞片放入清水中, 用牙刷轻轻刷去附着的黏膜, 取6片放在载玻片上, 然后盖上载玻片, 用胶带固定并用记号笔编号。最后放在显微镜下拍照保存。
1.3 年龄鉴定方法及年轮特征描述
根据殷名称[10]《鱼类生态学》的年轮鉴别特点及鉴定和分析鱼类年龄的方法, 在不清楚样本大小、性别的情况下对八种年龄材料进行年龄鉴定。每个年龄材料的年龄由同一观察者进行2次独立鉴定, 时间间隔不少于1周。
以体长大小为依据, 划分低龄、中龄和高龄3个年龄段[11], 并根据每一年龄段的同一样本, 描述8种年龄材料的年轮特征。
1.4 八种年龄材料的差异性
平均年龄的显著性分析 对8种年龄材料的平均年龄读数采用单因素方差分析(ANOVA)和多重比较进行统计检验和分析, 来解释8种年龄材料鉴定结果的差异性。
平均百分比误差(IAPE) 采用平均百分比误差(IAPE)来计算不同观察者年龄鉴定结果的精确性, 公式如下:
$ IAP{E_j} = \frac{1}{N}\sum\limits_{j = 1}^N {\left\{ {\frac{1}{R}\sum\limits_{{{i}} = 1}^R {\frac{{\left| {X_{ij} - X_j} \right|}}{X}} } \right\}} \times 100{{\%}} $

式中, N为进行年龄鉴定的鱼尾数; R为每尾鱼进行年龄鉴定的次数; Xij为第j尾鱼进行的第i次年龄鉴定结果; Xj为第j尾鱼的平均年龄。
为了更好地对不同年龄材料进行比较, 前后鉴定结果最终必须一致。若结果不一致, 则重新对该年龄材料进行年龄鉴定, 直到最终结果统一为止。若再次鉴定后, 结果差距较大, 则将其除去。在西藏其他裂腹鱼类[11, 12]的年龄材料鉴定中微耳石的清晰度是最好的, 并且双须叶须鱼微耳石的年轮比较清晰, 故以微耳石鉴定的年龄为准, 将星耳石、脊椎骨、鳃盖骨、臀鳞、胸鳞、侧线鳞和背鳞7种年龄材料分别和微耳石计算所得IAPE来进行比较。
清晰度评分 对8种年龄材料年轮的清晰度按照5个等级进行评分[11]: 1是非常好, 2是好, 3是一般, 4是很差, 5是难以辨认。
1.5 数据分析
统计分析采用SPSS 21和Excel 2003, 数据采用平均数±标准差表示, 当P<0.05时, 存在显著性差异。
2. 结果
总共采集到193尾双须叶须鱼, 体长为155—550 mm, 体重为46.5—1704.5 g。
2.1 年轮特征
微耳石 微耳石为不规则的椭圆形, 近极面前端膨大, 后端似铲型; 远极面微微隆起, 前端似由许多晶体组成, 从前端到后端逐渐变薄(图 1d1—d4)。中心核靠近前端, 在入射光下, 从微耳石核心向外边缘颜色逐渐变淡。在年轮排列区域, 年轮间距呈现有规律的缩短, 靠近中心核的年轮间距较大, 到10龄年轮间距明显变窄, 在20龄后年轮间距显著变窄(图 2f、图 3f、图 4g)。
![]() 图 1 双须叶须鱼背鳞(a)、侧线鳞(b)、胸鳞(c)、微耳石(d1、d2、d3、d4)、臀鳞(e)、鳃盖骨(f)、脊椎骨(g)、星耳石(h1、h2)的形态特征PZ为前区; UZ为上侧区; DZ为下侧区; BZ为后区; d1为远极面; d2为背面; d3为腹面; d4为近极面; h1为内侧面; h2为外侧面; 1为脊突; 2为翼叶; 3为主间沟; 4为基叶; 5为中央听沟; 6为叶突Figure 1. Morphological characteristics of back scale (a), lateral-line scale (b), chest scale (c), lapillus (d1, d2, d3, d4), hip scale (e), opercular bone (f), vertebrae (g), asteriscus (h1, h2) of Ptychobarbus dipogonPZ is the front zone; UZ is the upper zone; DZ is the lower zone; BZ is the rear zone; d1 is the far pole face; d2 is the back face; d3 is the ventral face; d4 is the near pole face; h1 is the inside face; h2 is the lateral side; 1 is the spinal protrusion; 2 is the wing leaf; 3 is the main groove; 4 is the basal leaf; 5 is the central auditory ditch; 6 is a leaf protrusion
图 1 双须叶须鱼背鳞(a)、侧线鳞(b)、胸鳞(c)、微耳石(d1、d2、d3、d4)、臀鳞(e)、鳃盖骨(f)、脊椎骨(g)、星耳石(h1、h2)的形态特征PZ为前区; UZ为上侧区; DZ为下侧区; BZ为后区; d1为远极面; d2为背面; d3为腹面; d4为近极面; h1为内侧面; h2为外侧面; 1为脊突; 2为翼叶; 3为主间沟; 4为基叶; 5为中央听沟; 6为叶突Figure 1. Morphological characteristics of back scale (a), lateral-line scale (b), chest scale (c), lapillus (d1, d2, d3, d4), hip scale (e), opercular bone (f), vertebrae (g), asteriscus (h1, h2) of Ptychobarbus dipogonPZ is the front zone; UZ is the upper zone; DZ is the lower zone; BZ is the rear zone; d1 is the far pole face; d2 is the back face; d3 is the ventral face; d4 is the near pole face; h1 is the inside face; h2 is the lateral side; 1 is the spinal protrusion; 2 is the wing leaf; 3 is the main groove; 4 is the basal leaf; 5 is the central auditory ditch; 6 is a leaf protrusion![]() 图 2 低龄双须叶须鱼(体长215 mm)背鳞(a)、侧线鳞(b)、胸鳞(c)、臀鳞(d)、星耳石(e)、微耳石(f)、鳃盖骨(g)、脊椎骨(h)的年轮特征(圆点示年轮)背鳞5龄(a); 侧线鳞4龄(b); 胸鳞5龄(c); 臀鳞5龄(d); 星耳石7龄(e); 微耳石7龄(f); 鳃盖骨7龄(g); 脊椎骨7龄(h)Figure 2. Annuli characteristics of back scale (a), lateral-line scale (b), chest scale (c), hip scale (d), asteriscus (e), lapillus (f), opercular bone (g), vertebrae (h) from the same Ptychobarbus dipogon with 215 mm SL(dots show annuli)back scale 5 years (a); lateral-line scale 4 years (b); chest scale 5 years (c); hip scale 5 years (d); asteriscus 7 years (e); lapillus 7 years (f); opercular bone 7 years (g); vertebrae 7 years (h)
图 2 低龄双须叶须鱼(体长215 mm)背鳞(a)、侧线鳞(b)、胸鳞(c)、臀鳞(d)、星耳石(e)、微耳石(f)、鳃盖骨(g)、脊椎骨(h)的年轮特征(圆点示年轮)背鳞5龄(a); 侧线鳞4龄(b); 胸鳞5龄(c); 臀鳞5龄(d); 星耳石7龄(e); 微耳石7龄(f); 鳃盖骨7龄(g); 脊椎骨7龄(h)Figure 2. Annuli characteristics of back scale (a), lateral-line scale (b), chest scale (c), hip scale (d), asteriscus (e), lapillus (f), opercular bone (g), vertebrae (h) from the same Ptychobarbus dipogon with 215 mm SL(dots show annuli)back scale 5 years (a); lateral-line scale 4 years (b); chest scale 5 years (c); hip scale 5 years (d); asteriscus 7 years (e); lapillus 7 years (f); opercular bone 7 years (g); vertebrae 7 years (h)![]() 图 3 中龄双须叶须鱼(体长400 mm)背鳞(a)、侧线鳞(b)、胸鳞(c)、臀鳞(d)、星耳石(e)、微耳石(f)、脊椎骨(g)、鳃盖骨(h)的年轮特征(圆点示年轮)背鳞9龄(a); 侧线鳞9龄(b); 胸鳞8龄(c); 臀鳞10龄(d); 星耳石17龄(e); 微耳石17龄(f); 脊椎骨15龄(g); 鳃盖骨15龄(h)Figure 3. Annuli characteristics of back scale (a), lateral-line scale (b), chest scale (c), hip scale (d), asteriscus (e), lapillus (f), vertebrae (g), opercular bone (h) from the same Ptychobarbus dipogon with 400 mm SL (dots show annuli)back scale 9 years (a); lateral-line scale 9 years (b); chest scale 8 years (c); hip scale 10 years (d); asteriscus 17 years (e); lapillus 17 years (f); vertebrae 15 years (g); opercular bone15 years (h)
图 3 中龄双须叶须鱼(体长400 mm)背鳞(a)、侧线鳞(b)、胸鳞(c)、臀鳞(d)、星耳石(e)、微耳石(f)、脊椎骨(g)、鳃盖骨(h)的年轮特征(圆点示年轮)背鳞9龄(a); 侧线鳞9龄(b); 胸鳞8龄(c); 臀鳞10龄(d); 星耳石17龄(e); 微耳石17龄(f); 脊椎骨15龄(g); 鳃盖骨15龄(h)Figure 3. Annuli characteristics of back scale (a), lateral-line scale (b), chest scale (c), hip scale (d), asteriscus (e), lapillus (f), vertebrae (g), opercular bone (h) from the same Ptychobarbus dipogon with 400 mm SL (dots show annuli)back scale 9 years (a); lateral-line scale 9 years (b); chest scale 8 years (c); hip scale 10 years (d); asteriscus 17 years (e); lapillus 17 years (f); vertebrae 15 years (g); opercular bone15 years (h)![]() 图 4 高龄双须叶须鱼(体长488 mm)背鳞(a)、侧线鳞(b)、胸鳞(c)、臀鳞(d)、鳃盖骨(e)、星耳石(f)、微耳石(g)、脊椎骨(h)的年轮特征(圆点示年轮)背鳞11龄(a); 侧线鳞12龄(b); 胸鳞12龄(c); 臀鳞16龄(d); 鳃盖骨23龄(e); 星耳石26龄(f); 微耳石34龄(g); 脊椎骨21龄(h)Figure 4. Annuli characteristics of back scale (a), lateral-line scale (b), chest scale (c), hip scale (d), opercular bone(e), asteriscus (f), lapillus (g), vertebrae (h) from the same Ptychobarbus dipogon with 488 mm SL (dots show annuli)back scale 11 years (a); lateral-line scale 12 years (b); chest scale 12 years (c); hip scale 16 years (d); opercular bone 23 years (e); asteriscus 26 years (f); lapillus 34 years (g); vertebrae 21 years (h)
图 4 高龄双须叶须鱼(体长488 mm)背鳞(a)、侧线鳞(b)、胸鳞(c)、臀鳞(d)、鳃盖骨(e)、星耳石(f)、微耳石(g)、脊椎骨(h)的年轮特征(圆点示年轮)背鳞11龄(a); 侧线鳞12龄(b); 胸鳞12龄(c); 臀鳞16龄(d); 鳃盖骨23龄(e); 星耳石26龄(f); 微耳石34龄(g); 脊椎骨21龄(h)Figure 4. Annuli characteristics of back scale (a), lateral-line scale (b), chest scale (c), hip scale (d), opercular bone(e), asteriscus (f), lapillus (g), vertebrae (h) from the same Ptychobarbus dipogon with 488 mm SL (dots show annuli)back scale 11 years (a); lateral-line scale 12 years (b); chest scale 12 years (c); hip scale 16 years (d); opercular bone 23 years (e); asteriscus 26 years (f); lapillus 34 years (g); vertebrae 21 years (h)星耳石 星耳石为星状, 轮纹不明显, 边缘为锯齿状, 内侧面中间长轴处为凹槽, 外侧面微微隆起, 具有辐射状脊(图 1h1、图 1h2)。到12龄间距明显变窄, 而到20龄后年轮间距显著变窄(图 2e、图 3e、图 4f)。
脊椎骨 脊椎骨为双凹型, 中心有一小孔, 前后凹面呈现出宽窄交替的同心圆轮纹, 在入射光下, 呈明暗交替分布。小孔的周围较为透明, 轮纹很少很细, 很难确认起始轮的位置。脊椎骨的边缘有较厚的结缔组织, 难以辨认末轮(图 1g)。脊椎骨的年轮宽度不会随着年龄的增加而显著变窄(图 2h、图 3g、图 4h)。
鳃盖骨 鳃盖骨为不规则的四边形, 基部内侧有许多小孔, 表面轮纹平行排列, 边缘为锯齿状。基部较厚, 且呈黄色, 难以辨认首轮位置(图 1f)。低龄鱼的鳃盖骨轮纹间隔大且稀疏, 不易分辨年轮; 高龄鱼轮纹排列紧密, 容易辨认。年轮间距呈现有规律的缩短, 到12龄年轮间距明显变窄(图 2g、图 3h、图 4e)。
臀鳞 臀鳞取自肛门至臀鳍的两侧, 大多数臀鳞形态特征特化, 下侧区向内弯曲, 呈“L”形。前区轮纹密集, 很难有效辨识年轮; 后区各年轮间隙较明显, 但夹杂副轮。有少部分鳞片上后区内的副轮与年轮十分相似, 较难区别, 需依靠侧区辅助观察(图 1e、图 2d、图 3d、图 4d)。
胸鳞、侧线鳞和背鳞 胸鳞取自侧线下方, 胸鳍基部(图 1c)。侧线鳞取自鱼体两侧的侧线部位, 中央有一条透明的管道(图 1b)。背鳞取自侧线上方, 背鳍附近(图 1a)。这3种环片疏密不明显, 但在入射光下可见到明亮的脊高出相邻环片, 脊的外缘即为年轮。前区、上侧区和下侧区均可作为年龄读取区。
2.2 八种年龄材料的比较
共采集193尾样本, 体长为155—550 mm, 体重为46.5—1704.5 g, 其中微耳石鉴定年龄在4—49龄, 星耳石鉴定年龄在4—35龄, 脊椎骨鉴定年龄范围在4—34龄, 鳃盖骨鉴定年龄范围在4—34龄, 臀鳞鉴定年龄范围在4—22龄, 胸鳞鉴定年龄范围在4—19龄, 侧线龄鉴定年龄范围在4—16龄, 背鳞鉴定年龄范围在4—17龄。不同年龄组的样本数和体长信息见表 1。
表 1 双须叶须鱼八种年龄材料所鉴定的各个年龄组的样本数和体长Table 1. Number of specimens and body length at different ages of Ptychobarbus dipogon年龄
Age (y)微耳石
Lapillus星耳石
Asteriscus脊椎骨
Vertebrae鳃盖骨
Opercular bones臀鳞
Hip scale胸鳞
Chest scale侧线鳞
Lateral-line scale背鳞
Back scaleN Mean±SD (mm) N Mean±SD (mm) N Mean±SD (mm) N Mean±SD (mm) N Mean±SD (mm) N Mean±SD (mm) N Mean±SD (mm) N Mean±SD (mm) 4 1 155.0 1 155.0 1 155.0 2 182.5±38.9 4 201.3±11.1 1 210 2 197.5±17.7 5 6 211.7±25.4 6 211.7±25.4 4 198.8±8.5 6 198.3±10.3 3 203.3±47.5 5 207.0±41.0 8 199.4±29.3 7 211.4±35.1 6 12 222.1±23.9 11 224.6±23.4 11 225.0±23.4 11 232.7±22.2 8 211.3±25.3 11 245.9±41.9 9 261.4±51.7 8 240.5±52.7 7 4 248.8±25.9 4 231.3±33.8 6 238.3±33.0 6 250.8±28.5 11 251.4±87.1 11 296.9±95.4 11 260.2±48.6 17 295.3±77.3 8 3 268.3±7.6 3 268.3±7.6 4 258.8±20.2 1 270.0 7 299.1±49.0 13 331.9±34.9 20 341.6±57.6 18 349.1±49.7 9 2 287.0±9.9 2 272.5±10.6 2 295.5±21.9 1 280.0 8 314.1±46.3 14 354.4±59.0 22 369.2±47.7 23 363.9±48.1 10 4 294.3±20.7 6 299.5±20.7 3 282.7±12.1 6 326.7±42.8 19 343.9±31.6 35 389.6±58.5 28 403.4±61.7 31 400.4±57.1 11 1 342.0 8 342.6±23.4 10 335.0±11.0 9 341.2±35.0 16 364.0±43.8 22 405.2±51.4 27 402.7±56.5 34 417.2±57.4 12 4 335.3±9.6 5 352.8±36.6 6 360.3±39.3 8 329.6±21.9 29 399.8±65.4 28 424.5±55.4 24 438.2±62.7 22 442.1±60.3 13 5 338.2±19.5 8 358.6±38.1 7 339.3±22.0 11 355.2±50.0 22 424.7±53.2 21 436.9±57.4 24 451.0±53.0 16 440.2±56.9 14 11 342.3±33.0 11 350.7±36.4 13 364.1±46.5 16 364.9±31.4 21 424.2±51.5 18 438.0±64.2 9 431.9±54.5 10 474.7±53.2 15 12 344.9±31.2 12 363.4±49.6 13 380.0±53.0 12 361.3±35.7 14 415.2±59.8 5 482.6±24.9 6 445.2±75.2 2 480.5±12.0 16 10 365.0±43.5 23 388.1±55.7 17 388.0±52.7 7 367.6±35.1 14 448.6±43.0 4 481.0±17.2 4 469.0±54.7 2 431.0±97.6 17 8 376.3±34.5 14 402.5±39.2 13 406.2±36.8 12 405.9±48.5 7 487.3±39.8 1 510.0 1 510.0 18 11 386.5±49.6 5 411.6±31.3 17 403.8±55.4 13 437.5±45.3 9 450.8±49.3 19 7 402.9±36.0 6 420.0±26.4 5 416.4±44.2 6 410.3±55.5 1 490.0 1 490.0 20 9 414.6±60.7 8 415.6±58.5 7 423.0±53.0 7 379.3±17.0 2 476.0±33.9 21 7 406.7±44.9 10 457.8±49.3 9 446.3±66.9 9 453.1±36.9 1 490.0 22 5 380.6±61.8 10 443.1±50.0 10 452.4±43.1 9 481.1±21.8 1 550.0 23 2 432.5±29.0 7 451.7±26.9 12 470.3±27.2 10 460.8±33.4 24 7 433.6±16.9 11 469.5±43.6 5 448.8±36.6 7 443.4±27.3 25 7 445.0±49.4 6 488.7±29.6 6 486.8±51.4 5 470.8±56.3 26 4 459.8±49.1 5 452.0±64.5 1 452.0 5 480.8±11.7 27 9 447.0±41.4 2 453.5±50.2 4 497.3±16.8 2 485.0±0.0 28 9 450.2±33.1 2 451.0±1.4 1 450.0 3 451.7±42.5 29 5 472.0±35.0 2 525.0±35.4 2 467.0±7.1 3 475.3±49.8 30 3 459.3±44.5 1 550.0 2 520.0 31 2 451.0±1.4 2 492.5±10.6 2 510.0±14.1 1 510.0 32 3 482.0±55.8 1 500.0 2 511.0±15.6 33 3 439.7±54.0 1 520.0 34 2 490.5±0.7 1 500.0 1 550.0 35 4 489.5±14.7 1 500.0 36 2 471.0±26.9 37 2 515.0±7.1 39 1 462.0 40 1 550.0 41 1 508.0 42 1 522.0 43 1 500.0 44 1 500.0 49 1 500.0 总计
Total193 193 193 193 193 193 193 193 2.3 八种年龄材料的平均年龄比较
双须叶须鱼不同年龄材料所鉴定的平均年龄见表 2。8种年龄材料所鉴定的平均年龄存在显著性差异(P<0.05)。用微耳石作为年龄材料所鉴定的平均年龄最高(20.05龄), 显著高于星耳石(16.84龄)、脊椎骨(16.56)、鳃盖骨(16.70龄)、臀鳞(11.34龄)、胸鳞(9.58龄)、侧线鳞(9.33龄)和背鳞(9.33龄)所鉴定的平均年龄(P<0.05)。23龄以下和23龄以上样本分开统计时, 4—23龄样本中, 微耳石鉴定的平均年龄与星耳石和脊椎骨鉴定的平均年龄较为接近, 分别为14.39龄、13.13龄、13.20龄, 显著高于鳃盖骨(12.96龄)、臀鳞(9.93龄)、胸鳞(8.49龄)、侧线鳞(8.30龄)和背鳞(8.13龄)所鉴定的平均年龄(P<0.05)。大于23龄时, 星耳石(23.52龄)、脊椎骨(22.61龄)和鳃盖骨(23.42龄)所鉴定的平均年龄较为接近, 星耳石、脊椎骨、鳃盖骨、臀鳞、胸鳞、侧线鳞和背鳞7种年龄鉴定材料都显著低于微耳石所鉴定的平均年龄30.23龄(P<0.05)。
表 2 不同年龄材料所鉴定的平均年龄的比较Table 2. The average age of different age materials年龄材料
Ages
structure平均年龄
Mean values of
age estimate年龄Total Age 4—14 Age 4—23 Age>23 微耳石
Lapillus20.05±9.32a 9.32±3.51a 14.39±5.26a 30.23±5.59a 星耳石
Asteriscus16.84±6.52b 9.11±3.25ab 13.13±4.44ab 23.52±3.71b 脊椎骨
Vertebrae16.56±6.18b 9.06±3.05ab 13.20±4.39ab 22.61±3.92b 鳃盖骨
Opercular
bones16.70±6.70b 8.98±3.28ab 12.96±4.62b 23.42±4.10b 臀鳞
Hip scale11.34±3.47c 7.74±2.49bc 9.93±3.04c 13.87±2.68c 胸鳞
Chest scale9.58±2.85d 6.62±2.26c 8.49±2.65d 11.54±2.03d 侧线鳞
Lateral-line
scale9.33±2.67d 6.49±1.85c 8.30±2.51d 11.17±1.81d 背鳞
Back scale9.33±2.68d 6.47±1.90c 8.13±2.41d 10.65±1.82d N 193 53 124 69 注: 表中同列数字上标不同字母表示有显著差异性(P<0.05)Note: Values with different superscripts in each column are significantly different from each other (P<0.05) 2.4 八种年龄材料的平均百分比误差比较
用微耳石鉴定年龄时, 多次读数的平均百分比误差(IAPE)最低(3.31%), 从小到大依次为: 星耳石(4.72%)、脊椎骨(4.79%)、臀鳞(5.11%)、鳃盖骨(5.17%)、胸鳞(5.19%)、侧线鳞(5.88%)、背鳞(5.86%)。其他7种年龄材料和微耳石比较的IAPE从小到大依次为星耳石(12.28%)、脊椎骨(15.67%)、鳃盖骨(17.81%)、臀鳞(41.63%)、侧线鳞(50.50%)、胸鳞(51.26%)、背鳞(51.74%)。从星耳石、脊椎骨、鳃盖骨、臀鳞、胸鳞、侧线鳞和背鳞分别与耳石作比较的IAPE值(图 5)可以看出, 星耳石、脊椎骨、鳃盖骨鉴定的年龄结果比四种鳞片准确。但仅从IAPE值来看, 并不能确定星耳石、脊椎骨和鳃盖骨做鉴定的年龄结果哪个更精确。但是统计分析表明(表 2), 星耳石和脊椎骨的年龄读数与微耳石的年龄读数在低于23龄时没有显著性差异(P>0.05), 而鳃盖骨所鉴定的年龄无论是在低龄还是高龄都显著低于耳石所鉴定的年龄(P<0.05)。随着年龄增加, IAPE值基本上呈现上升趋势, 差异也逐渐增大, 对于鉴定高龄鱼时所产生的差异越来越大。
![]() 图 5 星耳石、脊椎骨、鳃盖骨、臀鳞、胸鳞、侧线鳞和背鳞分别与微耳石比较的平均百分比误差(IAPE)分布图Figure 5. The average percentage error (IAPE) between asteriscus, vertebrae, opercular bone, hip scale, chest scale, lateral-line scale, back scale and lapillus
图 5 星耳石、脊椎骨、鳃盖骨、臀鳞、胸鳞、侧线鳞和背鳞分别与微耳石比较的平均百分比误差(IAPE)分布图Figure 5. The average percentage error (IAPE) between asteriscus, vertebrae, opercular bone, hip scale, chest scale, lateral-line scale, back scale and lapillus2.5 八种年龄材料的清晰度评分比较
由于4种鳞片年龄读数在高于14龄组内都与微耳石有显著差异, 因此在清晰度评分中分为低于14龄组和高于14龄组。微耳石上的年龄较清晰, 在低于14龄和高于14龄组内被评为“非常好”和“好”的都较其他材料多, 故微耳石在进行年龄鉴定时准确性最好(表 3、表 4)。
表 3 双须叶须鱼不同年龄材料的清晰度评分(4—14龄)Table 3. Distribution of readability scores for different age materials of Ptychobarbus dipogon (4—14 years)年龄材料
Ages structure清晰度评分比例
Proportion of readability score1 2 3 4 5 微耳石
Lapillus9.43 66.04 18.87 5.66 星耳石
Asteriscus1.89 43.40 35.85 16.98 1.89 脊椎骨
Vertebrae3.77 43.40 45.28 3.77 3.77 鳃盖骨
Opercular bones1.89 64.15 30.19 1.89 1.89 臀鳞
Hip scale16.98 56.60 24.53 1.89 胸鳞
Chest scale9.43 50.94 30.19 9.43 侧线鳞
Lateral-line scale11.32 56.60 28.30 1.89 1.89 背鳞
Back scale11.32 60.38 24.53 3.77 表 4 双须叶须鱼不同年龄材料的清晰度评分(14龄以上)Table 4. Distribution of readability scores for different calcified struc-tures of Ptychobarbus dipogon (more than 14 years)年龄材料
Ages structure清晰度评分比例
Proportion of readability score1 2 3 4 5 微耳石
Lapillus4.29 57.86 29.29 6.43 2.14 星耳石
Asteriscus2.14 49.29 35.00 11.43 2.14 脊椎骨
Vertebrae4.29 50.00 34.29 8.57 2.86 鳃盖骨
Opercular bone2.86 51.43 36.43 6.43 2.86 2.6 年龄与体长关系
微耳石年龄鉴定的年龄读数(A)与相对应的体长(L)呈极显著的对数函数关系(P<0.01), 其相关关系式为: L=146.76 ln (A)–38.139, R2=0.8312。随着年龄的增加, 双须叶须鱼体长随之增长, 但其增长速率随之减缓(图 6)。
3. 讨论
3.1 年龄鉴定的准确性比较
鱼类年轮特征的研究是进行鱼类年龄鉴定的前提[1]。而年龄鉴定的准确性直接关系到鱼类生长参数估算的可靠程度[13], 这对于渔业的管理和资源的合理开发尤为重要; 低估年龄的结果将导致对鱼类生长估计过快和自然死亡率估计过高, 因而对产量做出过于乐观的估计, 往往会造成资源的过度开发[14]。
就同一种鱼而言, 不同的年龄材料轮纹的清晰度和数目不一定相同, 所以可能导致不同年龄段的最适材料也有所不同[15]。本文微耳石鉴定所得的最大年龄为49龄, 星耳石鉴定所得的最大年龄为35龄, 脊椎骨鉴定所得最大年龄为34龄, 鳃盖骨为34龄, 臀鳞为22龄, 胸鳞为19龄, 侧线鳞为16龄, 背鳞为17龄。在低于23龄时, 微耳石与星耳石和脊椎骨鉴定的平均年龄之间无显著性差异(P>0.05), 相差年龄在1龄以内, 显著高于鳃盖骨和四种鳞片(P<0.05)。反映出在鉴定低龄个体时, 微耳石、星耳石和脊椎骨作为年龄材料是可行的。分析原因为, 鳃盖骨仅在边缘可观察到明暗相间排列的环纹, 鳃盖骨基部变厚[16], 中心轮纹判别能力差, 不易确定年龄[13]; 鳞片随年龄增长易出现磨损或者停滞现象[17], 通常会低估高龄和生长速率慢的个体年龄[13], 只适用于对低龄、生长较快的鱼类进行年龄鉴定[18], 特别是裂腹鱼特有的臀鳞与其繁殖行为密切相关, 致使鳞片的磨损很难避免[19], 裂腹鱼类的鳞片在长期进化过程中为适应高原寒冷的水域环境而存在着不同程度的退化[12]。
然而高于23龄时, 7种年龄鉴定材料均显著低于微耳石鉴定的平均年龄(P<0.05)。星耳石高龄个体年轮特征不明显, 杂纹较多[20]; 脊椎骨首轮较难辨认[21], 轮纹较为密集, 增加高龄鱼年龄鉴定的误差[11], 生活的地理环境特殊、环境条件恶劣, 脊椎骨上年轮的形成和排列可能受到环境条件的影响[11, 12]。而耳石年轮标志明显, 可判读力高[22]。耳石生长是以与机体生长相对独立, 在慢生长和高龄个体中比其他骨质材料生长更快, 从而能更真实地记录周期性季节生长和年龄[23], 所以采用耳石估计生长缓慢和相对长寿命群体或种类的年龄更为准确[24]。此外, 与微耳石比较的平均百分比误差相比, 星耳石、脊椎骨、鳃盖骨与之相差不大, 4种鳞片与之差别较大。并且IAPE值随着年龄增加基本上呈现上升趋势反映出在高龄个体年龄鉴定水平上所产生的差异越来越大。
在国内许多学者对裂腹鱼不同年龄鉴定材料的比较研究中, 得出耳石读数较其他年龄材料准确。马宝珊[11]和周贤君[12]分别通过研究异齿裂腹鱼和拉萨裂腹鱼的3种年龄材料(耳石、脊椎骨和鳃盖骨)发现耳石是年龄鉴定的最合适材料, 霍斌[25]研究尖裸鲤年龄材料发现釆用耳石作为年龄鉴定材料其准确性和精确性要优于脊椎骨, 并且耳石还是伊犁裂腹鱼[26]、色林错裸鲤[14]、软刺裸裂尻鱼[27]、青海湖裸鲤[3]的最佳年龄材料。
总而言之, 双须叶须鱼8种年龄鉴定材料均为每年形成一个年轮, 本研究通过不同年龄鉴定材料的比较发现, 在进行年龄鉴定时微耳石所鉴定的最大年龄较其他年龄材料大, 而其他的几种年龄材料均有不同程度的年龄阶段性差异。在鉴定低龄个体时, 微耳石是双须叶须鱼年龄鉴定的最佳材料, 星耳石和脊椎骨次之, 鳃盖骨较差, 鳞片不宜作为年龄鉴定材料; 在鉴定高龄个体时, 微耳石是双须叶须鱼年龄鉴定的最佳材料。
3.2 双须叶须鱼年龄结构与年龄鉴定探讨
杨鑫[6]研究发现雅鲁藏布江中游的双须叶须鱼群体的年龄结构为3—24 龄, 反映出雅鲁藏布江双须叶须鱼群体年龄结构趋于简单。Li等[7]研究了雅鲁藏布江及其支流的双须叶须鱼, 其种群的年龄结构分别为2—44 龄, 与本文研究结果类似, 反映出双须叶须鱼在雅鲁藏布江中游流域年龄趋向低龄化, 而双须叶须鱼在拉萨河流域的年龄结构群体尚未受到严重干扰。究其原因, 采样点环境和人为干扰强度等差异是导致种群结构产生差异的主要因素[6]。而且在3—6月, 双须叶须鱼正处于繁殖期, 加之浮游生物在此季节繁殖生长, 易捕捞较大个体。
在不同水域中双须叶须鱼[6]2种年龄材料的比较显示, 脊椎骨与耳石鉴定的结果无显著性差异, 耳石鉴定10 龄以上的年龄大于脊椎骨鉴定的年龄。而本文研究仅是在鉴定低龄个体(小于23龄)时, 微耳石、星耳石和脊椎骨作为年龄材料是可行的。分析其原因, 可能是由于不同水域资源现状的差异和人为干扰的程度不同所致。交叉水域中双须叶须鱼3种耳石年龄材料的比较显示, 核心模棱两可的星耳石导致难以辨别年龄, 矢耳石易脆并经常破裂[7], 所以微耳石是双须叶须鱼的最佳年龄鉴定材料。
总而言之, 无论是不同水域或者交叉水域, 微耳石一直是双须叶须鱼最好的年龄鉴定材料, 这与杨鑫[6]、Li等[7]的研究结果一致。
-
![]()
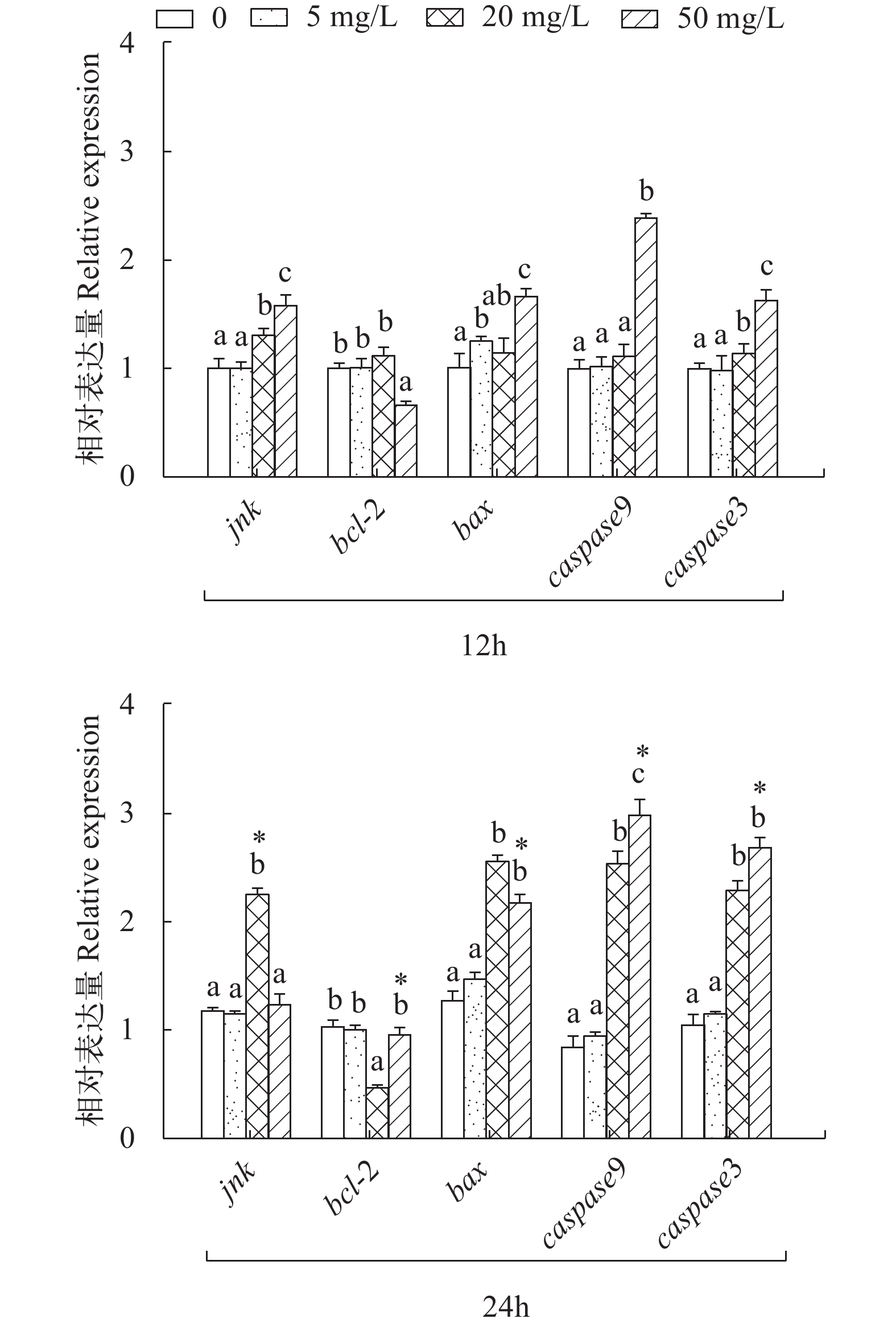
图 1 亚硝酸钠对L8824细胞jnk、bcl-2、bax、caspase9和caspase3表达量的影响
不同字母表示同一时间点组内具有显著性差异(P<0.05), *表示不同时间点组间具有显著性差异(P<0.05), 下同
Figure 1. Effects of sodium nitrite on the expression of jnk, bcl-2, bax, caspase9 and caspase3 in L8824 cells
Different letters indicates significant differences between groups at the same time point (P<0.05), * indicates significant differences between groups at different time points (P<0.05), the same applies below
![]()
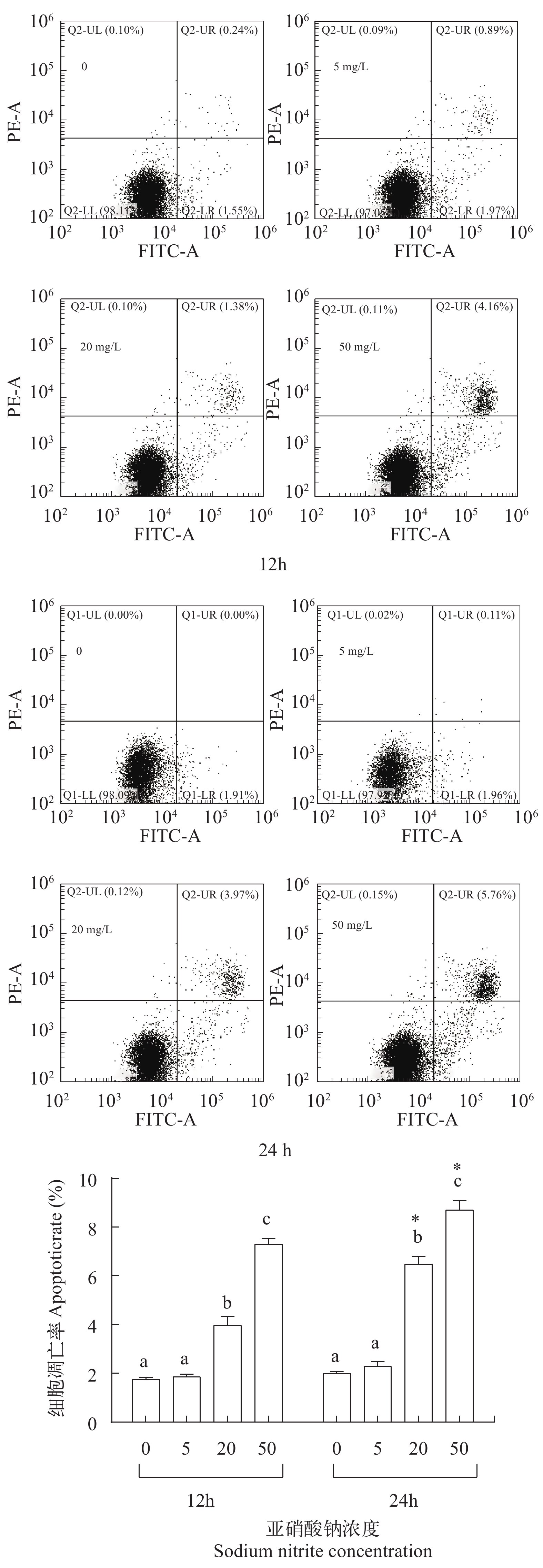
图 2 亚硝酸钠对L8824细胞凋亡的影响
Figure 2. The effect of sodium nitrite on apoptosis of L8824 cells
![]()
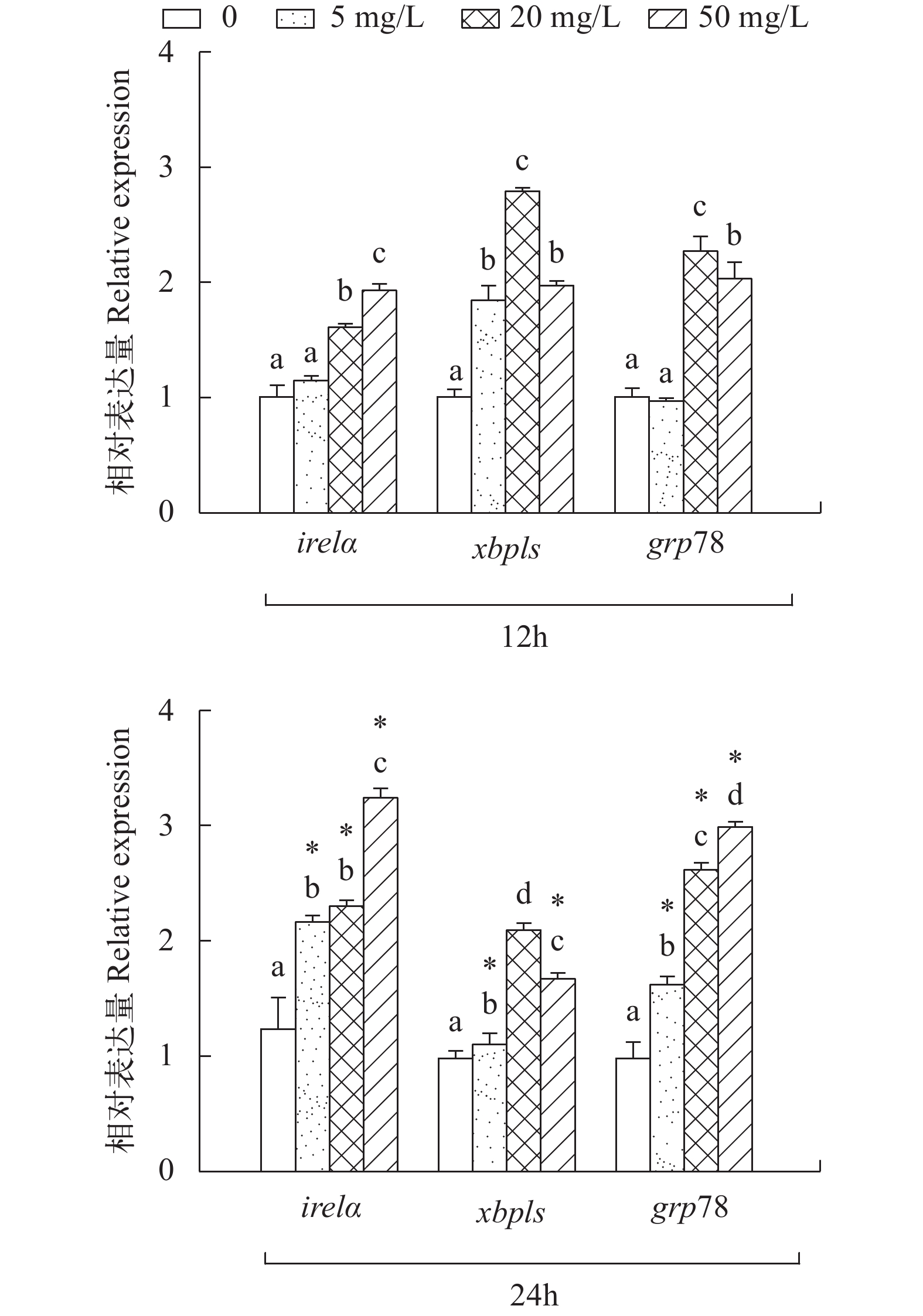
图 3 亚硝酸钠对L8824细胞ire1α, xbp1s和grp78表达量的影响
Figure 3. Effects of sodium nitrite on the expression of ire1α, xbp1s and grp78 in L8824 cells
![]()
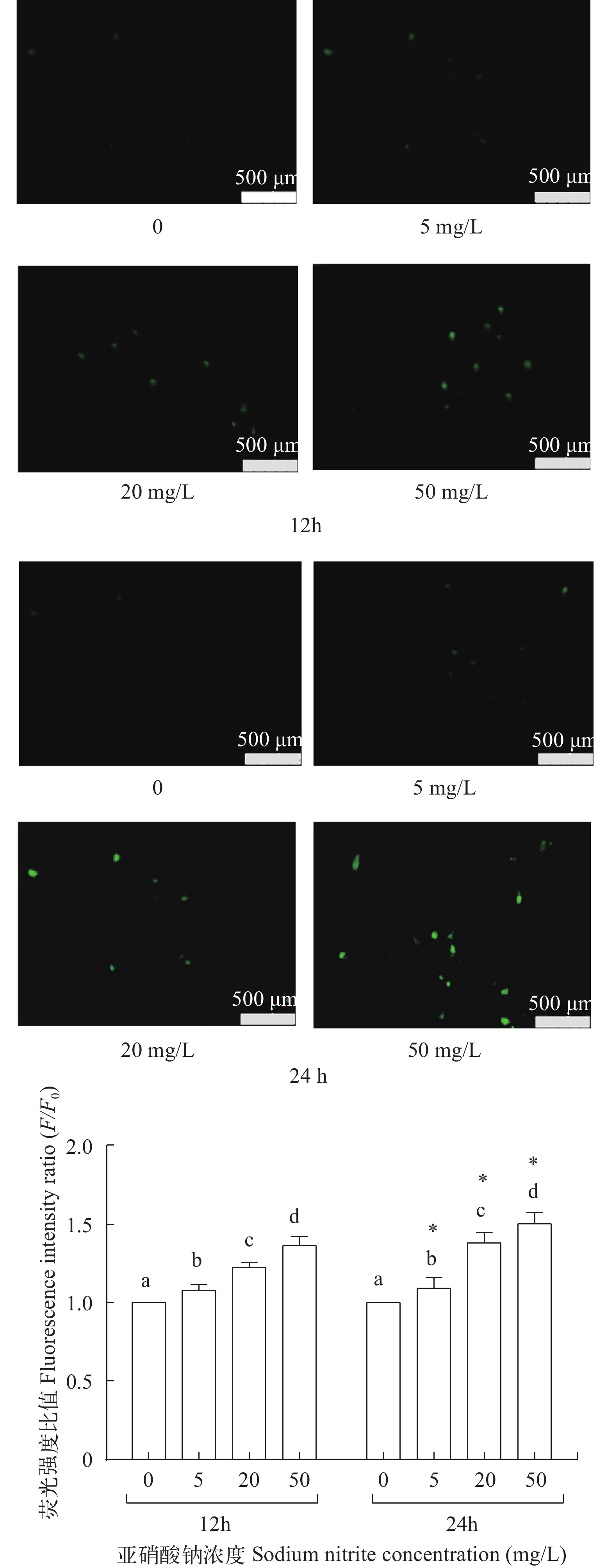
图 4 亚硝酸钠对L8824细胞内钙离子浓度的影响
Figure 4. The effect of sodium nitrite on the intracellular calcium content in L8824 cells
![]()
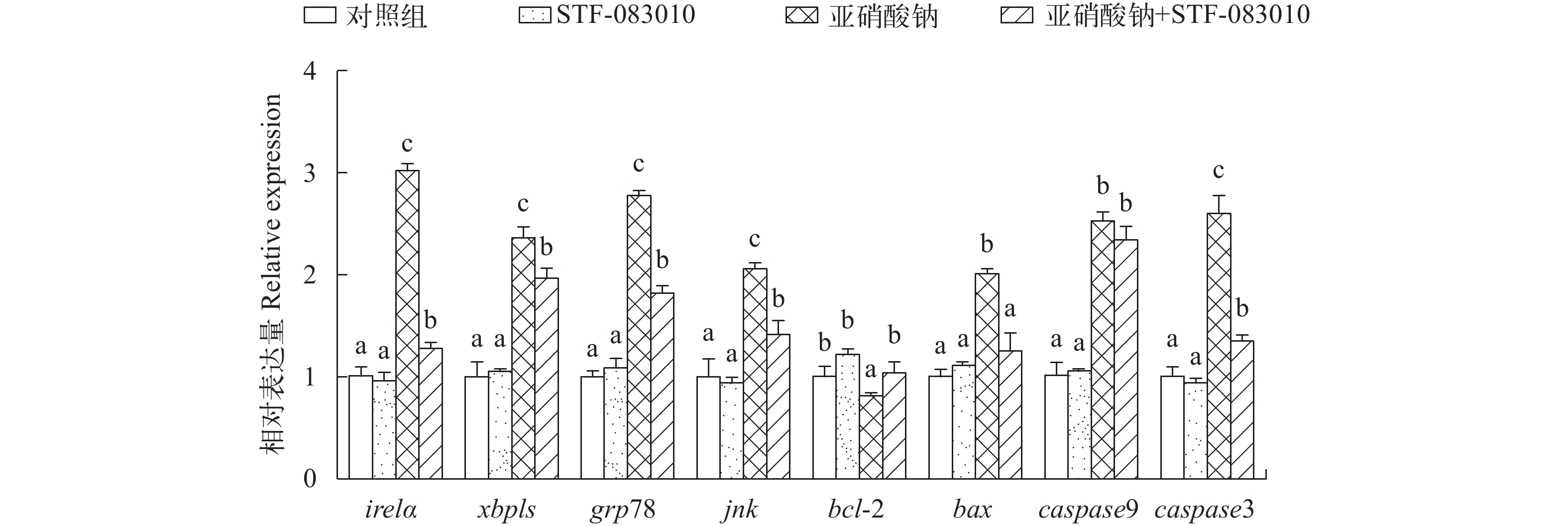
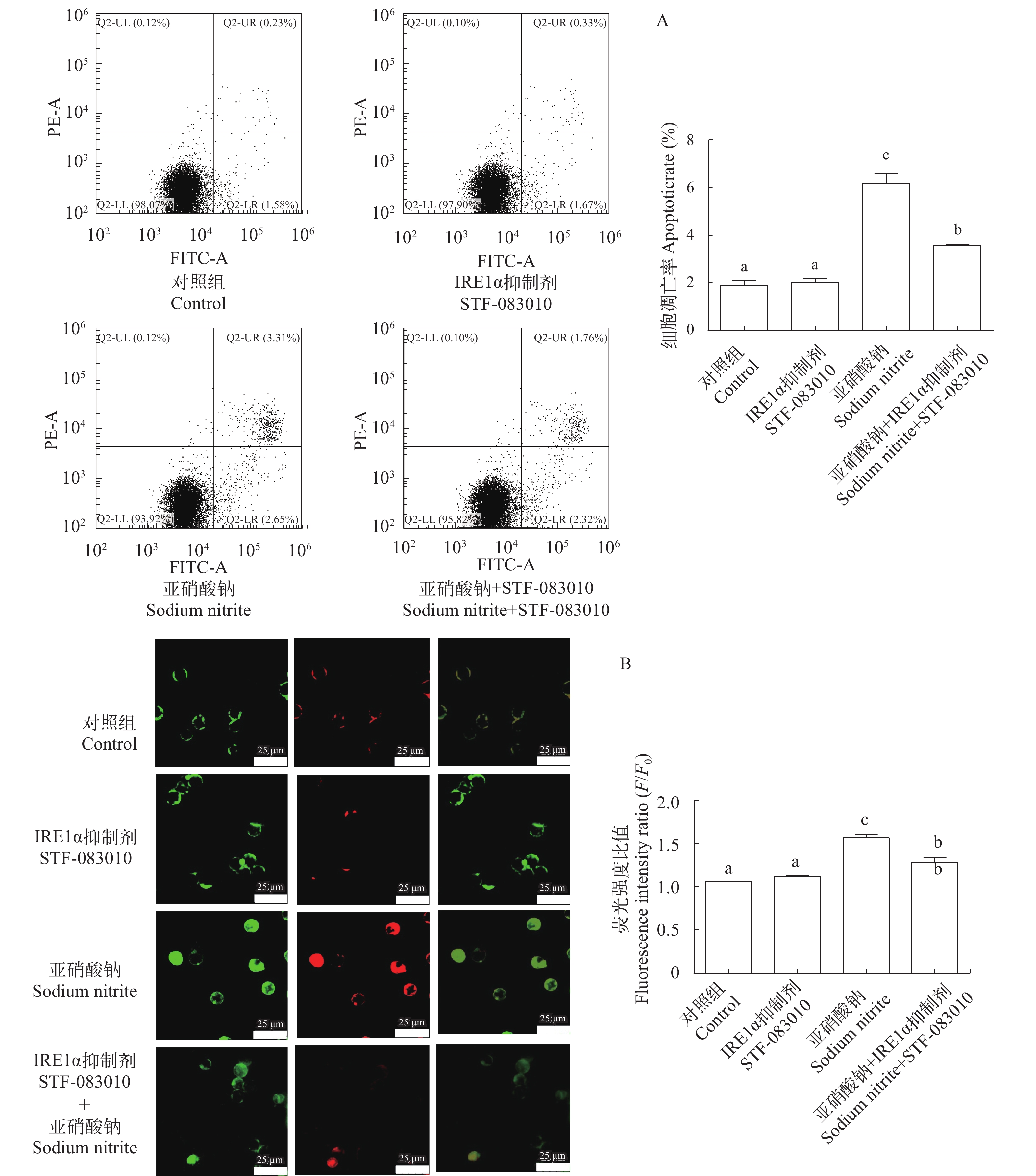
图 5 亚硝酸钠和STF-083010及其配伍对L8824细胞ire1α、xbp1s、grp78、jnk、bcl-2、bax、caspase9和caspase3表达量的影响
Figure 5. Effects of sodium nitrite, STF-083010 and their combination on the expression of ire1α, xbp1s, grp78, jnk, bcl-2, bax, caspase9 and caspase3 in L8824 cells
![]()
图 6 亚硝酸钠和STF-083010及其配伍对L8824细胞凋亡的影响
A. 流式Annexin-FITC/PI 双染法检测L8824细胞凋亡; B. 流式Annexin-FITC/PI 双染法在荧光显微镜下检测L8824细胞凋亡, Annexin-FITC染色为绿色荧光, PI染色为红色荧光
Figure 6. Effects of sodium nitrite, STF-083010 and their combination on the cell apoptosis of L8824 cells
A. Detection of apoptosis in L8824 cells by flow Annexin-FITC/PI double staining; B. Annexin-FITC/PI double staining was used to detect apoptosis of L8824 cells under fluorescence microscope, Annexin-FITC staining was green fluorescence and PI staining was red fluorescence
![]()
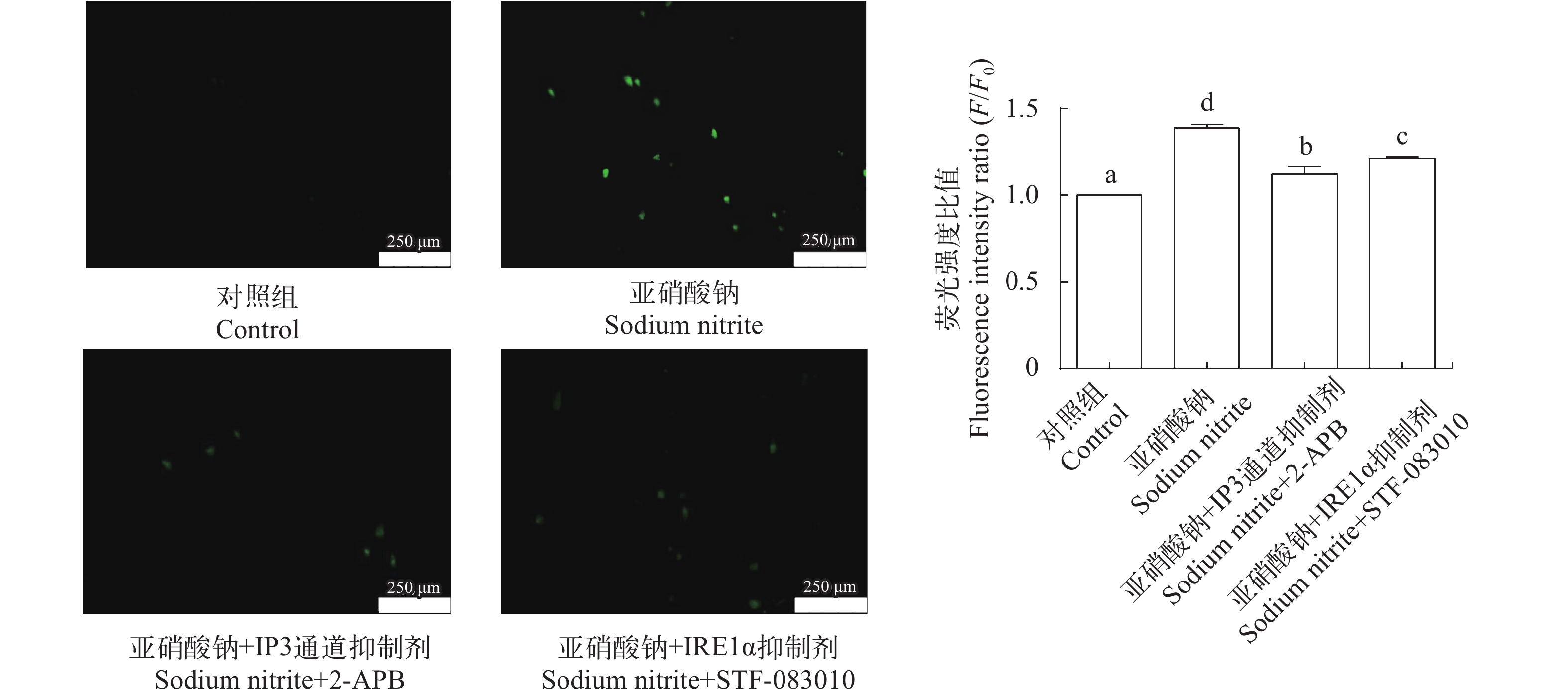
图 7 2-APB和STF-083010 对亚硝酸钠暴露L8824细胞内钙离子浓度的影响
Figure 7. Effects of 2-APB and STF-083010 on the intracellular calcium content under sodium nitrite exposure in L8824 cells
表 1 引物序列
Table 1 Primers used in the study
目的基因Target gene 引物序列Primer sequences (5′-3′) 作物长度Product length (bp) jnk F: ACAGCGTAGATGTGGGTGATT
R: GCTCAAGGTTGTGGTCATACG108 bcl-2 F: CCACCCATTGTCATACCTAACG
R: ACCCACA GGGCTGTCACTTCT98 bax F: GGATCAGTTGGGTGGCGTTGA
R: TGAGTCGGTTGAAGAGCAGAGT104 caspase9 F: GGGATAGATGACCAGATGGA
R: GGGATAGATGACCAGATGGA164 caspase3 F: GGCAGATGGTGACAGTGATGGA
R: TGGCAGATTGGAGGAGGATTCG154 ire1α F: GAACGCCACATACTCTGA
R: TGTCCACTGTCACCACTA146 xbp1s F: AGGGGAAGTGAATACGGGGAAT
R: GACCTCACAGAAGGCAACGGAATC102 grp78 F: GGTGCTTAATTCTGCGTTAT
R: CCTTCTTCTCGTCGTCTTC92 β-actin F: CACTGTGCCCATCTACGA
R: CCATCTCCTGCTCGAAGTC80  下载: 导出CSV
下载: 导出CSV
表 2 不同浓度亚硝酸钠对L8824 细胞活力的影响
Table 2 The effect of sodium nitrite on the viability of L8824 cells
亚硝酸钠浓度Sodium nitrite concentration (mg/L) 细胞活力Cell viability(%) 12h 24h 0 100.00±0.00c 100.00±0.00c 5 98.44±1.46bc 97.65±1.94c 20 97.68±0.47b 93.67±0.83b 50 90.05±1.74a 87.44±0.76a 注: 同列上标小写字母不同表示二者之间差异显著(P<0.05); 下同
Note: Values in the same row with different superscripts are significantly different (P<0.05); the same applies below
下载: 导出CSV
表 3 亚硝酸钠和STF-083010共同作用对L8824细胞活力的影响
Table 3 Effects of sodium nitrite and STF-083010 on the viability of L8824 cells
组别 Group 细胞活力 Cell viability (%) 对照组 Control 100.00±0.00c IRE1α抑制剂 STF-083010 99.18±0.52c 亚硝酸钠 Sodium nitrite 92.64±1.01a IRE1α抑制剂+亚硝酸钠 STF-083010+Sodium nitrite 96.31±1.38b
下载: 导出CSV
-
[1] Jensen F B. Nitrite disrupts multiple physiological functions in aquatic animals [J]. Comparative Biochemistry and Physiology Part A Molecular and Integrative Physiology, 2003, 135(1): 9-24. doi: 10.1016/S1095-6433(02)00323-9
[2] 余瑞兰, 聂湘平, 魏泰莉, 等. 分子氨和亚硝酸盐对鱼类的危害及其对策 [J]. 中国水产科学, 1999, 6(3): 73-77. doi: 10.3321/j.issn:1005-8737.1999.03.017 Yu R L, Nie X P, Wei T L, et al. Toxicity of molecular ammonia and nitrite to fishes and the control measures [J]. Journal of Fishery Sciences of China, 1999, 6(3): 73-77. doi: 10.3321/j.issn:1005-8737.1999.03.017
[3] 王鸿泰, 胡德高. 池塘中亚硝酸盐对草鱼种的毒害及防治 [J]. 水产学报, 1989, 13(3): 207-214. Wang H T, Hu D G. Toxicity of nitrite to Ctenopharyngodon idella in ponds and its way of prevention [J]. Journal of Fisheries of China, 1989, 13(3): 207-214.
[4] Sun S, Ge X, Xuan F, et al. Nitrite-induced hepatotoxicity in Bluntsnout bream (Megalobrama amblycephala): The mechanistic insight from transcriptome to physiology analysis [J]. Environmental Toxicology and Pharmacology, 2014, 37(1): 55-65. doi: 10.1016/j.etap.2013.11.010
[5] Marcio Jose dos Santos Silva, Leme F P, Takata R, et al. Biological responses of Neotropical freshwater fish Lophiosilurus alexandri exposed to ammonia and nitrite [J]. Science of the Total Environment, 2018(616-617): 1566-1575.
[6] Svobodova Z, Machova J, Poleszczuk G, et al. Nitrite poisoning of fish in aquaculture facilities with water-recirculating systems [J]. Acta Veterinaria Brno, 2005, 74(1): 9584-9585.
[7] Wang W N, Wang A L, Zhang Y J, et al. Effects of nitrite on lethal and immune response of Macrobrachium nipponense [J]. Aquaculture, 2004, 232(1-4): 679-686. doi: 10.1016/j.aquaculture.2003.08.018
[8] Fu L Z, Yong Q L, Qi S, et al. Effects of sodium nitrite on proliferation and apoptosis on human hepatocarcinoma cells [J]. Chinese Pharmacological Bulletin, 2011, 27(2): 191-195.
[9] 屈三甫, 张小榕, 郑从义, 等. 鱼呼肠孤病毒诱导草鱼肾细胞凋亡 [J]. 水生生物学报, 2000, 24(6): 616-620. Qu S P, Zhang X R, Zheng C Y, et al. Apoptosis of the cell line from grass carp (CIK) induced by fish reovirus [J]. Acta Hydrobiologica Sinica, 2000, 24(6): 616-620.
[10] Walter P, Ron D. The unfolded protein response: from stress pathway to homeostatic regulation [J]. Science, 2011, 334(6059): 1081-1086. doi: 10.1126/science.1209038
[11] Xiang C, Wang Y, Zhang H, et al. The role of endoplasmic reticulum stress in neurodegenerative disease [J]. Apoptosis, 2017, 22(1): 1-26. doi: 10.1007/s10495-016-1296-4
[12] Xu Z, Chikka M R, Xia H, et al. Ire1 supports normal ER differentiation in developing Drosophila photore ceptors [J]. Journal of Cell Science, 2016, 129(5): 921-929. doi: 10.1242/jcs.180406
[13] Cao S S, Kaufman R J. Endoplasmic reticulum stress and oxidative stress in cell fate decision and human disease [J]. Antioxidants and Redox Signaling, 2014, 21(3): 396-413. doi: 10.1089/ars.2014.5851
[14] Schroder M. Endoplasmic reticulum stress responses [J]. Cellular and Molecular Life Sciences, 2008, 65(6): 862-894. doi: 10.1007/s00018-007-7383-5
[15] Liu X H, Zhang Z Y, Sun S, et al. Ischemic postconditioning protects myocardium from ische-mia/reperfusion injury through attenuating endoplasmic reticulum stress [J]. Shock, 2008, 30(4): 422-427. doi: 10.1097/SHK.0b013e318164ca29
[16] Long M, Lin W, Hou J, et al. Dietary supplementation with selenium yeast and tea polyphenols improve growth performance and nitrite tolerance of Wuchang bream (Megalobrama amblycephala) [J]. Fish and Shellfish Immunology, 2017(68): 74-83. doi: 10.1016/j.fsi.2017.07.017
[17] 李雪贤, 孙健, 吉红, 等. 脂肪酸影响草鱼肝细胞脂质蓄积状态及诱导其凋亡的离体研究 [J]. 水生生物学报, 2017, 41(1): 56-64. doi: 10.7541/2017.8 Li X X, Su J, Ji H, et al. Influence of fatty acids on lipid accumulation and apoptosis status of grass carp Ctenopharyngodon idellus hepatocyte in vitro [J]. Acta Hydrobiologica Sinica, 2017, 41(1): 56-64. doi: 10.7541/2017.8
[18] 萧培珍, 吉红, 张宝彤, 等. 水飞蓟素抑制草鱼肝细胞脂质蓄积的作用及其机制研究 [J]. 水生生物学报, 2017, 41(6): 137-146. Xiao P Z, Ji H, Zhang B T, et al. Inhibitory effect of silymarin on oleic acid-induced lipid accumulation in grass carp (Ctenopharyngodon idellus) hepatocyte in vitro [J]. Acta Hydrobiologica Sinica, 2017, 41(6): 137-146.
[19] Papandreou I, Denko N C, Olson M, et al. Identification of an Ire1alpha endonuclease specific inhibitor with cytotoxic activity against human multiple myeloma [J]. Blood, 2011, 117(4): 1311-1314. doi: 10.1182/blood-2010-08-303099
[20] Rodrigues A D S L, Gnocchi K G, Medeiros L C C, et al. Acute toxicity of nitrite in fat snook (Centropomus parallelus) [J]. Marine and Freshwater Behaviour and Physiology, 2018, 51(2): 1-13.
[21] Le T H G, Jensen F B, Damsgaard C, et al. Extreme nitrite tolerance in the clown knifefish Chitala ornata is linked to up-regulation of methaemoglobin reductase activity [J]. Aquatic Toxicology, 2017, (187): 9-17.
[22] Evans D H, Piermarini P M, Potts WTW. Ionic transport in the fish gill epithelium [J]. Journal of Experimental Zoology, 1999, 283(7): 641-652. doi: 10.1002/(SICI)1097-010X(19990601)283:7<641::AID-JEZ3>3.0.CO;2-W
[23] Onken H, Putzenlechner M. A V-ATPase drives active, electrogenic and Na+-independent Cl- absorption across the gills of Eriocheir sinensis [J]. Journal of Experimental Biology, 1995, 198(3): 767-774.
[24] Bath R N, Eddy F B. Transport of nitrite across fish gills [J]. Journal of Experimental Zoology, 1980, 214(1): 119-121. doi: 10.1002/jez.1402140115
[25] Krous S R, Blazer V S, Meade T L. Effect of acclimation time on nitrite movement across the gill epithelia of rainbow trout: the role of “chloride cells” [J]. Progressive Fish Culturist, 1982, 44(3): 126-130. doi: 10.1577/1548-8659(1982)44[126:EOATON]2.0.CO;2
[26] Campos M L, Barbas Luis André Luz, Fiori N L, et al. Oxidative stress and antioxidant responses in juvenile Brazilian flounder Paralichthys orbignyanus exposed to sublethal levels of nitrite [J]. Fish Physiology and Biochemistry, 2018(44): 1349-1362.
[27] 强俊, 徐跑, 何杰, 等. 氨氮与拥挤胁迫对吉富品系尼罗罗非鱼幼鱼生长和肝脏抗氧化指标的联合影响 [J]. 水产学报, 2011, 35(12): 1837-1848. Qiang J, Xu P, He J, et al. The combined effects of ammonia nitrogen and crowding stress on the growth and liver antioxidant index of the Nile tilapia juveniles [J]. Journal of Fisheries, 2011, 35(12): 1837-1848.
[28] Doblander C, Lackner R. Metabolism and detoxification of nitrite by trout hepatocytes [J]. Biochimica et Biophysica Acta, 1996, 1289(2): 270-274. doi: 10.1016/0304-4165(95)00166-2
[29] Michael M I, Hilmy A M, El-Domiaty N A, et al. Serum transminases activity and histopathological changes in Clarias lazera chronically exposed to nitrite [J]. Comparative Biochemistry and Physiology Part C Comparative Pharmacology, 1987, 86(2): 255-262.
[30] Xian J A, Wang A L, Hao X M, et al. In vitro toxicity of nitrite on haemocytes of the tiger shrimp, Penaeus monodon, using flow cytometric analysis [J]. Comparative Biochemistry and Physiology Part C: Toxicology and Pharmacology, 2012, 156(2): 75-79. doi: 10.1016/j.cbpc.2012.04.001
[31] Al-Gayyar M M, Hassan H M, Alyoussef A, et al. Nigella sativa oil attenuates chronic nephrotoxicity in-duced by oral sodium nitrite: Effects on tissue fibrosis and apoptosis [J]. Redox Report, 2015, 21(2): 50-60.
[32] Al-Rasheed N M, Fadda L M, Attia H A, et al. Quercetin inhibits sodium nitrite-induced inflammation and apoptosis in different rats organs by suppressing Bax, HIF1-α, TGF-β, Smad-2, and AKT pathways [J]. Journal of Biochemical and Molecular Toxicology, 2016, 31(5): 1-7.
[33] Liu C, Huang Y, Zhang Y, et al. Intracellular methylglyoxal induces oxidative damage to pancreatic beta cell line INS-1 cell through Ire1α-JNK and mitochondrial apoptotic pathway [J]. Free Radic Research, 2017, 51(4): 1-14.
[34] 等. 过氧化亚硝酸盐在人血管内皮引起内质网应激和凋亡预示动脉粥样硬化形成 [J]. 中国动脉硬化杂志, 2006, 25(12): 623-629. Dickhout, Hossain, Pozza, et al. Endoplasmic reticulum stress and apoptosis induced by nitrite peroxide in human vascular endothelium predict atherosclerosis [J]. Chinese Journal of Arteriosclerosis, 2006, 25(12): 623-629. [Dickhout, Hossain, Pozza,
[35] Dudek J, Benedix J, Cappel S, et al. Functions and pathologies of BiP and its interaction partners [J]. Cellular and Molecular Life Sciences (CMLS)
, 2009, 66(9): 1556-1569. doi: 10.1007/s00018-009-8745-y [36] Argon Y, Simen B B. GRP94, an ER chaperone with protein and peptide binding properties [J]. Seminars in Cell and Developmental Biology, 1999, 10(5): 495-505. doi: 10.1006/scdb.1999.0320
[37] Lievremont J P, Rizzuto R, Hendershot L, et al. BiP, a major chaperone protein of the endoplasmic reticulum lumen, plays a direct and important role in the storage of the rapidly exchanging pool of Ca2+ [J]. Journal of Biological Chemistry, 1997, 272(49): 30873-30879. doi: 10.1074/jbc.272.49.30873
[38] Ron D, Walter P. Signal integration in the endoplasmic reticulum unfolded protein response [J]. Nature Reviews Molecular Cell Biology, 2007, 8(7): 519-529. doi: 10.1038/nrm2199
[39] Pan Y, Xu J, Wang X, et al. The role of cell apoptosis mediated by endoplasmic reticulum stress (ERS) of deep tissue injury of pressure ulcer of rats [J]. Chinese Journal of Applied Physiology, 2015, 31(5): 396-400.
[40] Kato H, Nakajima S, Saito Y, et al. mTORC1 serves ER stress-triggered apoptosis via selective activation of the IRE1-JNK pathway [J]. Cell Death and Differentiation, 2012, 19(2): 310-320. doi: 10.1038/cdd.2011.98
[41] Huang M, Xu A, Wu X, et al. Japanese encephalitis virus induces apoptosis by the IRE1/JNK pathway of ER stress response in BHK-21 cells [J]. Archives of Virology, 2016, 161(3): 699-703. doi: 10.1007/s00705-015-2715-5
-
期刊类型引用(2)
1. 陈家琪,董丽,麻晓梅,田凯,白洁,赵彦伟. 基于eDNA技术的白洋淀微型生物群落监测. 农业环境科学学报. 2021(08): 1773-1786+1839 .  百度学术
百度学术
2. 姚新民,陈云,沈燕,李禹昕,龚美萍,金凤,邵未艾,刁永卿,师乐,胡洁兰. 2013—2019年上海市青浦区出厂水水质状况及趋势分析. 职业与健康. 2021(21): 2976-2978+2982 . 百度学术
其他类型引用(3)






计量
- 文章访问数: 3753
- HTML全文浏览量: 975
- PDF下载量: 84
- 被引次数: 5