
EFFECTS OF DIETARY VITAMIN D3 ON THE GROWTH AND EXPRESSION OF TLR18, TLR19 AND TLR21 IN THE JUVENILE YELLOW CATFISH PELTEOBAGRUS FULVIDRACO
-
摘要: 为了探讨饲料中维生素D3添加水平对黄颡鱼(Pelteobagrus fulvidraco)生长和Toll样受体的影响, 研究设计了5个不同浓度梯度的维生素D3饲料(1120、2260、3950、8030和16600 IU/kg), 对体重为(5.0±0.2) g的黄颡鱼进行了为期12周的生长实验, 并在生长实验结束后进行鮰爱德华氏菌(Edwardsiella ictaluri)攻毒72h。于攻毒前(0)和攻毒后(72h)采样, 每个饲料组分别取6尾鱼的脾脏、头肾、肝脏和前肠四个组织, 检测不同浓度维生素D3处理对攻毒前和攻毒后TLR18、TLR19和TLR21基因表达量的影响。同时另取6条新鲜黄颡鱼的肌肉、头肾、肾脏、皮肤、脑、鳃、脾脏、胃上皮、小肠和肝脏, 检测TLR18、TLR19和TLR21基因在黄颡鱼中的组织分布。结果表明: 不同的维生素D3添加水平会显著影响黄颡鱼幼鱼的生长性能; TLR18、TLR19和TLR21基因在所检测的组织中均有表达, 但在脾脏中表达量最高; 饲料中不同维生素D3含量在攻毒前后均会显著影响TLR18、TLR19和TLR21在头肾、脾脏、肝脏和前肠中的表达, 攻毒后基因的表达显著高于攻毒前; TLR18、TLR19和TLR21在不同组织中的表达和饲料中维生素D3的浓度相关, 研究结果表明饲料中添加合适剂量的维生素D3, 可以促进相关免疫基因的表达, 从而增强黄颡鱼对病原微生物的抵抗力。Abstract: This experiment was conducted to study the effects of dietary vitamin D3 levels on the growth performance and the expression of TLR18, TLR19 and TLR21 in Pelteobagrus fulvidraco. A total of 800 healthy Pelteobagrus fulvidraco with an average body weight of (5.0±0.2) g were randomly assigned to 5 groups with 4 replicates per group and 40 fishes per replicate. Juvenile yellow catfish were fed with the basal diet supplemented with 1120, 2260, 3950, 8030 and 16600 IU/kg vitamin D3. After a 12-week growth trial, 50 fish from each treatment were injected with Edwardsiella ictaluri, and the fish were challenged for 72h. The spleen, head kidney, liver and anterior intestine from fish in each group were collected at 0 and 72h after the Edwardsiella ictaluri challenge and were used to detect the expression of three toll-like receptors, i.e., TLR18, TLR19 and TLR21. Meanwhile, the muscles, head kidney, kidney, skin, brain, gill, spleen, stomach, anterior intestine and liver were collected from fresh nonexperimental yellow catfish to detect the tissue distribution of TLR18, TLR19 and TLR21. The results showed that different levels of dietary vitamin D3 can significantly affect growth performance. The TLR18, TLR19 and TLR21 genes were expressed in all tested tissues, but the highest expression was found in the spleen. The Edwardsiella ictaluri challenge significantly increased the expression of TLR18, TLR19 and TLR21. Different levels of dietary vitamin D3 had a significant influence on TLR18, TLR19 and TLR21 gene mRNA expression in the head kidney, spleen, liver and anterior intestine both before and after the pathogen challenge. These results showed that appropriate supplementation with dietary vitamin D3 can stimulate the growth and pathogen resistance of juvenile yellow catfish.
-
Keywords:
- Pelteobagrus fulvidraco /
- Vitamin D3 /
- TLR18 /
- TLR19 /
- TLR21
-
浮游动物作为次级生产者, 是水域食物网中的重要一环, 其种类和数量的变动会直接或间接的影响着水体中其他水生生物的分布[1]。由于浮游动物对水环境变化的敏感性和适应性差异, 利用浮游动物群落结构的变化来检测和评价水环境具有重要的应用价值, 弥补水体理化指标在水质评价上的不足[2]。近年来相关研究显示, 利用浮游动物多样性指数可以反映水体的富营养化状况和水质受污染程度[3—6]。同时, 由于我国水体污染状况严峻, 各大湖泊、流域、水库等也均积极开展相关的生态学调查, 利用浮游动物群落结构变化及多样性指数对水质进行评价[7, 8]。
沙颍河是淮河流域最大的一条支流, 西起河南省伏牛山, 跨豫、皖两省, 流经郑州、平顶山、漯河、周口、阜阳等市, 东至安徽省颍上县沫河口入淮。全长约620 km, 流域面积39880 km2, 接近淮河流域总面积的1/7, 其中周口以上为沙颍河的上游部分, 长324 km, 流域面积为25880 km2; 周口到阜阳市是沙颍河的中游部分, 河长172 km; 阜阳市以下为下游部分, 河长124 km[9]。自1989年以来, 淮河干流共发生5次较大污染, 都与沙颍河有关[10]。尤其, 沙颍河流域中的贾鲁河中牟段主要承担着郑州市工业废水和城市生活污水排泄, 排放量逐年增长, 河道及地下水均受到不同程度的污染, 河流断面水质超过Ⅴ类水标准, 由于水体严重污染、生态环境水量不足, 造成生态环境恶化, 水生植物、鱼类、底栖生物、微生物、湿地等大多消亡, 既丧失了水体的自净能力, 也破坏了河道滨水景观环境[11]。
目前, 关于沙颍河流域的调查研究包括沙颍河河南段水质理化指标的时空变化[12, 13]、浮游甲壳动物时空变化[14]、沙颍河流域着生藻和大型底栖无脊椎动物[15]等的研究, 缺乏对整个沙颍河流域浮游动物的系统调查研究。本次研究以沙颍河流域主要支流贾鲁河和其干流沙颍河为研究主体, 调查沙颍河流域浮游动物群落结构的空间变化, 并利用浮游动物评价水质污染状况。本研究可为沙颍河流域水环境治理、水体生态环境持续健康发展和淮河流域水环境治理等提供理论依据。
1. 材料与方法
1.1 采样时间与采样点设置
2016年秋季对沙颍河流域浮游动物进行调查研究, 共设置20个采样点。其中, 9月中旬采样点为14个, 10月初又补充采样点6个。在每个采样点均有重复样。具体采样位置如图 1, 详细信息见表 1。所有采样点均经GPS定位。
1.2 浮游动物样品的采集与鉴定
浮游动物定性样品: 以25号筛绢制成的浮游生物网(孔径64 μm)在表层水面以下呈“∞”字形捞取3—5min, 并将滤取的样本放入标本瓶中, 加4%的福尔马林溶液固定。
浮游动物定量样品: 用2.5 L的有机玻璃采水器(水生所制)取表层和底层的水样混合, 再从混合样中取10 L和1 L分别用于大型浮游甲壳类和轮虫的定量。10 L样先用孔径64 μm的浮游生物网过滤, 收集于50 mL的塑料瓶中并用4%的福尔马林溶液保存; 1 L样加Lugol氏液固定后带回实验室沉降, 自然沉降48h后, 用虹吸管除去上清液收集沉积物再浓缩成50 mL并保存。
参照分类资料[16—22]的描述, 在OLYMPUS BX53显微镜和解剖镜下进行浮游动物鉴定和计数。
表 1 沙颍河流域各采样位点详细情况Table 1. Distributions of sampling sites in the Shaying River Basin样品编号
Sample number采样地点
Sampling site位置(经度, 纬度)
Location (longitude and latitude)样点类型
Sites type河流名称
River name1 郑州市惠济区祥云寺 E113°43′45.71″, N34°52′6.74″ 河道 贾鲁河 2 郑州市中牟县白沙镇 E113°52′′37.28″, N34°46′31.49″ 河道 贾鲁河 3 郑州市中牟县明山庙村 E114°0′12.92″, N34°45′41.91″ 河道 贾鲁河 4 郑州市中牟县陈桥村 E114°06′32.26″, N34°40′54.28″ 河道 贾鲁河 5 扶沟县摆渡口村 E114°20′11.85″, N34°9′21.44″ 河道 贾鲁河 6 新密市大槐镇铁匠沟村 E113°29′23.62″, N34°26′16.06″ 河道 双洎河 7 新密市大槐镇桃园村 E113°31′29.48″, N34°26′8.57″ 河道 双洎河 8 新密市新寨遗址 E113°32′54.25″, N34°26′12.45″ 河道 双洎河 9 扶沟县双洎河桥头 E114°18′49.24″, N34°9′39.26″ 河道 双洎河 10 周口市商水县邓城镇 E114°36′28.49″, N33°37′52.36″ 河道 沙河 11 西华县徐桥村徐桥 E114°29′51.65″, N33°42′12.21″ 河道 颍河 12 周口市西沙河大桥 E114°18′48.81″, N34°9′40.18″ 河道 沙颍河汇合 13 周口市李埠乡村牛滩村 E114°43′45.88″, N33°35′12.93″ 河道 沙颍河 14 周口项城市郑埠航运枢纽 E114°52′12.72″, N33°30′35.32″ 河道 沙颍河 15 周口市沈丘县纸店镇沙河大桥 E114°52′0.91″, N33°30′37.00″ 河道 沙颍河 16 阜阳市沙颍河国家湿地公园 E115°16′35.19″, N33°18′9.00″ 河道 沙颍河 17 阜阳市袁集镇沙颍河特大桥 E115°54′52.60″, N33°50′25.57″ 河道 沙颍河 18 阜阳市颍上县老沫口子 E115°54′53.14″, N33°50′27.10″ 河道 沙颍河 19 阜阳市颍上县老沫口子 E115°54′54.10″, N33°50′27.10″ 河道 淮河(干流) 20 阜阳市颍上县老沫口子 E115°54′54.42″, N33°50′27.64″ 河道 淮河(干流) 1.3 数据分析
运用优势度Y计数出优势物种, 计算公式为: Y=(Ni/N) fi, 式中Ni为第i种的个体数, N为样品中所有种类的个体数, fi为第i种的出现频率, Y>0.02为优势种。
$H' = - \sum {\left( {\frac{{{n_i}}}{N}} \right)} \left( {{{\log }_2}\frac{{{n_i}}}{N}} \right)\;\;J' = \frac{{H'}}{{{{\log }_2}S}}$

运用Shannon-Wiener指数(H′)[23]来表示浮游动物群落的多样性, 式中, N为浮游动物总个体数, ni为第i种的个体数。同时, 运用Pielou指数(J′)[24]表示物种均匀度, 式中, S为物种总数。
2. 结果
2.1 浮游动物的种类组成
通过对20个采样点的采样及检测分析, 发现共有浮游动物36属78种, 其中, 轮虫20属60种, 占浮游动物总物种数的76.92%; 枝角类10属12种, 占浮游动物总物种数的15.38%; 桡足类6属6种, 占浮游动物总物种数的7.69%。总体上而言, 沙颍河流域浮游动物物种类型以轮虫为主。各采样点具体物种数详见图 2, 可以明显看出贾鲁河浮游动物种类数较多, 双洎河浮游动物种类数相对较低, 且沿着沙颍河支流汇入淮河干流的过程中, 浮游动物的种类数呈现先增加后逐渐降低的趋势。
2.2 浮游动物的优势种
如表 2所示, 由于桡足类各采样点优势种主要为桡足类无节幼体和桡足类幼体, 因此文中未计算其优势度, 仅计算轮虫、枝角类的优势度, 结果表明: 沙颍河流域轮虫主要优势种为9种, 枝角类主要优势种为4种。各个采样位点浮游动物的优势种为1至7种。其中, 沙颍河流域上游贾鲁河浮游动物的优势种为长肢多肢轮虫、裂痕龟纹轮虫、角突臂尾轮虫、萼花臂尾轮虫、微型裸腹溞、象鼻溞、圆形盘肠溞; 双洎河优势种为长肢多肢轮虫、罗氏异尾轮虫、角突臂尾轮虫、萼花臂尾轮虫、狭甲轮虫、微型裸腹溞、象鼻溞、圆形盘长溞, 上游浮游动物优势种多为喜富营养化条件下的浮游动物。沙颍河流域中下游浮游动物优势种为长肢多肢轮虫、裂痕龟纹轮虫、角突臂尾轮虫、曲腿龟甲轮虫、无柄轮虫、奇异六腕轮虫、微型裸腹溞、象鼻溞。
![]() 图 2 沙颍河流域各采样点浮游动物各类群物种数Figure 2. The abundance of each species of zooplankton in different sampling sites in the Shaying River Basin
图 2 沙颍河流域各采样点浮游动物各类群物种数Figure 2. The abundance of each species of zooplankton in different sampling sites in the Shaying River Basin2.3 浮游动物的密度与生物量
各采样点浮游动物密度为125—13701 ind./L(图 3) 。轮虫在4号采样点的密度最高为13650 ind./L, 以长肢多肢轮虫、角突臂尾轮虫、萼花臂尾轮虫、三肢轮虫等为主要组成部分, 其他采样点轮虫的密度为125—7889 ind./L, 以长肢多肢轮虫和裂痕龟纹轮虫为主要组成; 枝角类在12号采样点的密度最高为380 ind./L, 以微型裸腹溞和短尾秀体溞为主要组成部分; 桡足类在12号采样点的密度最高为817 ind./L, 主要以桡足类幼体为主要组成部分; 无节幼体在9号采样点密度最高, 为1550 ind./L。
从生物量(图 4)上而言, 浮游动物的总生物量范围为0.15—9.32 mg/L。轮虫在4号采样点的生物量最高为9.16 mg/L; 枝角类和桡足类在12号采样点的生物量最高, 分别为3.68和2.64 mg/L; 无节幼体在9号采样点的生物量最高, 为4.65 mg/L。
2.4 浮游动物多样性指数和均匀度指数
检测位点中的Shannon-Wiener多样性指数H′为1.03—3.51, Pielou指数J′的变化范围为0.26— 0.70(图 5)。当H′大于3.0时表示水体轻污染或无污染, 1.0<H′<3.0时表示水体中污染,H′<1.0表示水体重污染。同时, 当0.5<J′<0.8表示水质轻污染或无污染, 0.3<J′<0.5表示水质中污染,J′<0.3表示水质重污染。参照评价标准[25]对检测位点的水质进行评价: 沙颍河流域各采样位点的水质, 除了4号、5号、15号、17号表现出轻污染或无污染, 其他采样位点水质均为中污染或重污染。
表 2 沙颍河流域浮游动物优势种空间变化及其优势度Table 2. Spatial variations of dominant species and dominance of zooplankton in the Shaying River Basin种名Species 拉丁名Latin name 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 长肢多肢轮虫 Polyarthra
dolichoptera0.35 0.54 0.22 0.20 0.21 0.22 0.66 0.12 0.43 0.43 0.51 0.21 0.12 0.39 0.37 0.43 裂痕龟纹轮虫 Anuraeopsis fissa 0.05 0.03 0.11 0.05 0.04 0.17 0.14 0.26 0.04 0.03 0.09 罗氏异尾轮虫 Trichocerca rousseleti 0.07 0.13 0.11 角突臂尾轮虫 Brachionus angularis 0.05 0.04 0.05 0.03 0.04 0.13 萼花臂尾轮虫 B. calyciflorus 0.02 0.03 0.06 0.03 0.11 曲腿龟甲轮虫 KeratelIa valga 0.09 0.03 0.02 0.20 0.03 0.21 0.18 0.16 一种狭甲轮虫 Colurella sp. 0.25 0.25 一种无柄轮虫 Ascomopha sp. 0.14 奇异六腕轮虫 Hexarthra mira 0.12 0.20 0.02 微型裸腹溞 Moina micrura 0.69 0.27 0.03 0.19 0.23 象鼻溞 Bosmina sp. 0.03 0.10 0.62 0.50 0.69 0.92 0.02 0.90 0.92 0.51 0.92 0.67 0.89 0.62 0.68 短尾秀体溞 Diaphanosoma
brachyurum0.18 圆形盘肠溞 Chydorus sphaericus 0.56 0.52 0.67 0.17 ![]() 图 5 沙颍河流域各采样点浮游动物的生物多样性指数和物种均匀度指数Figure 5. Shannon-Wiener index and Pielou evenness index of zooplankton in different sites in the Shaying River表 3 淮河流域浮游动物群落结构的时间变化Table 3. Temporal variations of zooplankton community structure in the Huaihe River Basin
图 5 沙颍河流域各采样点浮游动物的生物多样性指数和物种均匀度指数Figure 5. Shannon-Wiener index and Pielou evenness index of zooplankton in different sites in the Shaying River表 3 淮河流域浮游动物群落结构的时间变化Table 3. Temporal variations of zooplankton community structure in the Huaihe River Basin时间
Date地点
Station原生动物
Protozoa轮虫
Rotifera枝角类
Cladocera桡足类
Copepoda备注
Remarks1982年5月 淮河干流
蚌埠闸[31]765 ind./L
0.02 mg/L30 ind./L
0.06 mg/L57 ind./L
1.14 mg/L35 ind./L
1.05 mg/L1982年蚌埠闸上水体中耐污
能力弱的桡足和枝角类的数
量分别为35 ind./L和57 ind./L2005年3月—
2005年5月淮河蚌
埠段[32]N N 2属3种 5属5种 浮游甲壳动物密度为1.75—
67.51 ind./L, 水体为重污染,
水质很差2006年5月 淮河干流
蚌埠闸[31]10 ind./L
0.0005 mg/L9 ind./L
0.001 mg/L0.10 ind./L
0.001 mg/L0.02 ind./L
0.004 mg/L2006年耐污能力弱的桡足和枝
角类的这两类种群数量分别为
0.10 ind./L和0.02 ind./L2007年1月—
2007年12月淮河中
游[33]N N 8属13种 9属11种 浮游甲壳动物密度季节变动范
围为7.9—58.0 ind./L, 年均值为
13.2 ind./L2011年3月—
2011年12月淮河干
流[8]27属69种1.7×
106 ind./L 87 mg/L35属104种1.7×
106 ind./L
2048 mg/L12属27种
3410 ind./L
68 mg/L5属6种7.7×
104 ind./L
249 mg/L淮河干流上游水质为轻污染,
中下游为中污染或重污染2011年3月—
2012年2月阜阳城
区[34]23属30种4138—7856 ind./L
0.15—0.35 mg/L23属58种1061—3356 ind./L
1.26—3.87 mg/L15属34种56—136 ind./L
0.9—1.7 mg/L6属12种12—
84 ind./L
1.56—
10.92 mg/L阜阳城区各水体均处于富营养
状态, 部分位点为重污染2011年4月—
2012年1月沙颍河阜
阳段[14]N N 7属11种 8属9种 浮游甲壳动物密度春季最高为
51.8 ind./L, 冬季最低为4.5 ind./L,
水体为轻度-中度污染2012年6月—
2013年8月淮河流域
信阳段[35]4属5种 8属17种 6属9种 3属5种 轮虫、枝角类和桡足类种类和数
量较多, 并且枝角类和桡足类生
物量也较大2012年12月—
2013年7月淮河中
上游[36]N 19属46种
3.3—391.2 ind./LN N 水质从冬季到夏季有恶化趋势,
营养化程度有增加的趋势
水体为轻度-中度富营养2013年8月—
2014年1月沙颍河
流域[37]N 85种 9种 7种 轮虫密度明显高于甲壳类浮游
动物的密度2014年冬季 淮河淮
南段[38]22种 6种 3种 7种 浮游动物密度为1313.17 ind./L,
生物量为0.23 mg/L, 水体为
中度污染注: N表示无数据 Note: N means no data 3. 讨论
3.1 沙颍河流域浮游动物群落结构变化特征
近年来国内外越来越多的学者关注于利用浮游动物评价水体生态系统是否健康[26, 27], 这是由于浮游动物个体小, 具有较短的世代交替周期, 对环境变化反应迅速。随着水体富营养化程度提高, 浮游动物群落结构趋于简单, 种类变少, 丰度增加, 多样性指数下降, 群落的稳定性降低[28]。因此, 浮游动物群落结构变化也成为了水生态系统中调查的热点。例如甬江干流[29]调查报道了轮虫24属72种, 枝角类5属20种, 桡足类11属13种, 湘江干流[30]报道了轮虫23属45种, 枝角类9属17种, 桡足类3属5种, 并利用浮游动物对水质进行了评价, 这些均说明浮游动物研究在水生态系统监测方面具有重要意义。
通过对比分析淮河流域浮游动物的历年调查研究发现(表 3), 淮河流域浮游动物研究主要集中在蚌埠、信阳、阜阳、淮河中上游沙颍河流域及干流, 浮游动物的种类数以小型浮游动物为主。此次, 沙颍河流域的20个采样点调查检测到浮游动物36属78种(不包括原生动物), 其中轮虫20属60种, 枝角类10属12种, 桡足类6属6种, 表明沙颍河流域浮游动物主要以轮虫为主。国内关于河流流域浮游动物调查结果也均显示浮游动物以小型浮游动物为主[8, 33, 39]。吴利等[8]2011年对淮河干流调查发现轮虫194种, 枝角类27种, 桡足类6种。左其亭等[28]2012年对淮河中上游轮虫群落结构的调查发现轮虫有46种。Zhao等[37]2013年对沙颍河流域调查发现轮虫85种, 枝角类9种, 桡足类7种。这与此次2016年沙颍河调查结果相比, 轮虫种类数有所下降, 枝角类种类数有所增加, 桡足类种类数没有太大变化。
3.2 沙颍河流域浮游动物密度和生物量空间变化特征
2011年淮河干流[8]轮虫密度高达1.7×106 ind./L, 生物量为2048 mg/L, 枝角类密度为3410 ind./L, 生物量为68 mg/L, 桡足类密度为7.7×104 ind./L, 生物量为249 mg/L; 同年, 沙颍河流域下游的阜阳城区[34]轮虫密度为1061—3356 ind./L, 生物量为1.26—3.87 mg/L, 枝角类密度为56—136 ind./L, 生物量为0.90—1.70 mg/L, 桡足类密度为12—84 ind./L, 生物量为1.56—10.92 mg/L。本研究各采样点浮游动物密度为125—13701 ind./L, 浮游动物的总生物量范围为0.15—9.32 mg/L。与2011年淮河干流浮游动物的调查结果相比较, 沙颍河流域浮游动物的密度和生物量相对偏低, 但与国内其他河流流域[29, 30]相比较而言, 此次沙颍河流域浮游动物的密度和生物量相对较高。有研究表明[40]轮虫喜欢富营养环境, 水体营养状态的提升有利于轮虫种类密度的提高, 枝角类喜还原性的中营养或贫营养的清洁水体, 枝角类对环境的响应能力更为敏感, 而桡足类受环境因子的影响较少。沙颍河流域结果显示浮游动物密度主要是由轮虫占大多数比例, 浮游甲壳类密度所占比例较低。同时, 通过表 3可明显发现该流域水体富营养污染严重, 这可能就是导致浮游动物密度较高的原因。此外, 浮游动物密度和生物量峰值出现在贾鲁河和双洎河流域, 地理位置处于沙颍河流域的上游, 从整体上而言, 随着水流汇入淮河过程中浮游动物的密度和生物量空间变化趋势相一致, 呈大致下降趋势。
3.3 沙颍河流域浮游动物对水质的总体评价
在本研究中, 沙颍河流域各采样点浮游动物优势种为1—7种, 常见优势种为长肢多肢轮虫、裂痕龟纹轮虫、角突臂尾轮虫、象鼻溞和微型裸腹溞, 沙颍河上游浮游动物优势种还分布有萼花臂尾轮虫和圆形盘肠溞等富营养化指示种, 中下游分布出现曲腿龟甲轮虫等喜清洁环境的指示种[22, 41]。沙颍河流域大多数优势种为喜富营养化水体的种类。综合分析各采样点的种群反映出沙颍河流域上游水体富营养化较为严重, 中下游水体富营养化有所缓解。
Shannon-Wiener多样性指数(H′)是用来反映生物群落结构的复杂性和稳定性, 表示群落物种内部及物种间分配均匀性的整体性指标, 在一定程度上可以指示水体环境。同时, Pielou指数(J′)描述群落中各种间个体分布均匀程度。沙颍河流域采样位点Shannon-Wiener多样性指数H′为1.03—3.51, Pielou指数J′的变化范围为0.26—0.70。参照水生生物评价水质的标准沙颍河流域水质总体呈现出中污染-重污染。
自1982年以来, 淮河流域浮游动物变化详见表 3, 淮河蚌埠段浮游动物变化明显, 耐污性浮游动物种类数降低, 水体质量变差[31, 32]; 淮南段[38]和信阳段[35]水体为轻污染-中污染; 淮河上游流域水体处于轻污染状态, 中下游水体为中-重污染状态[8, 33, 36]。此外, 有研究表明整个沙颍河流域属颍河周口段及其支流贾鲁河的重金属污染问题突出[42], 这说明水体污染状况很严重。此次调查结果显示贾鲁河上游(即1至3号采样点)水体为中-重污染, 贾鲁河下游(即4至5号采样点)水体为轻污染或无污染, 周口段水体为中污染, 表明周口段及贾鲁河水体污染问题依然需要严峻对待和治理。2011年沙颍河流域阜阳段[14]调查研究浮游甲壳动物发现水体为轻度-中度污染, 但阜阳城区[34]水体均处于富营养状态, 城区部分位点为重污染。历经5年之后, 本研究利用浮游动物评价水体状况, 发现阜阳段水体为中污染状态, 阜阳的水体污染状况依旧需要认真对待。据统计[43], 在我国的大多数河流中, 上游生物多样性指数较高, 下游多样性降低, 但是沙颍河流域却是上游个别采样点多样性指数低, 下游多样性指数相对较高。这可能是因为沙颍河上游城市人口聚集及生活污水排放导致水质污染严重, 多样性指数低; 下游人口稀少, 人为影响较少, 故水体状况相对良好。
-
![]()
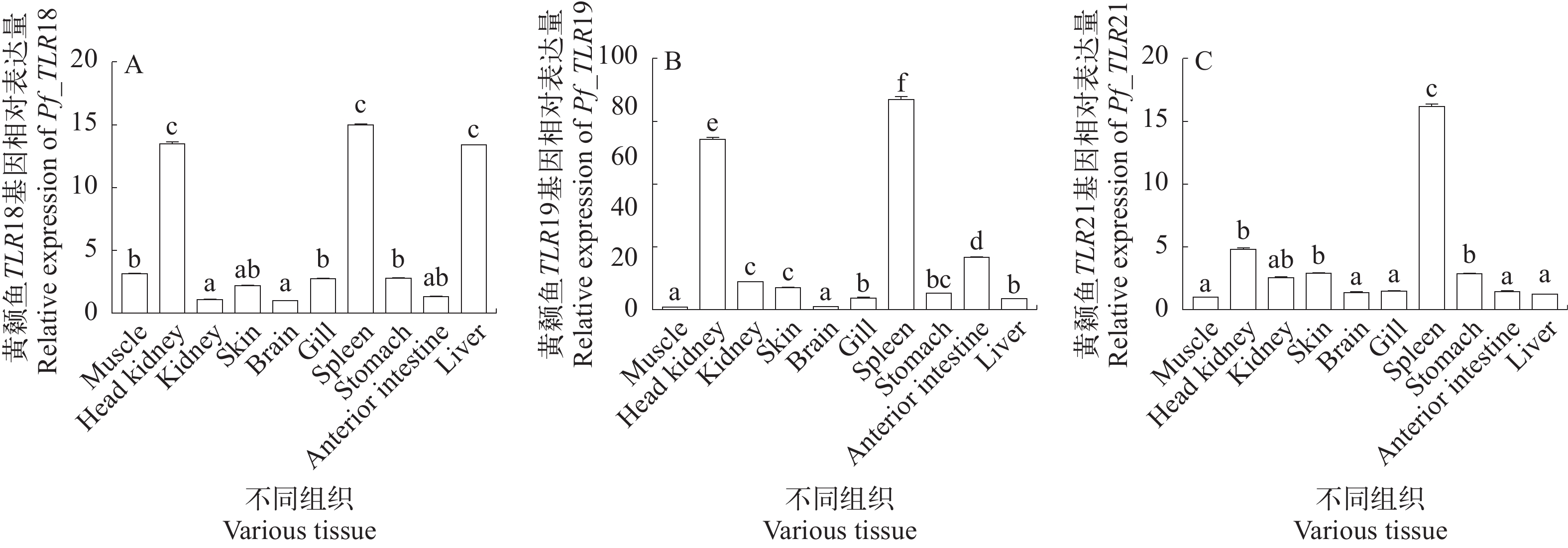
图 1 黄颡鱼TLR18 (A)、TLR19 (B)和TLR21 (C)基因mRNA组织表达分布
Muscle. 肌肉; Head kidney. 头肾; Kidney. 肾脏; Skin. 皮肤; Brain. 脑; Gill. 鳃; Spleen. 脾脏; Stomach. 胃; Anterior intestine. 前肠; Liver. 肝脏. 字母代表显著性差异(Duncan检验, P<0.05). Letters indicate significant difference (Duncan test, P<0.05)
Figure 1. Tissue distribution of TLR18(A), TLR19(B), and TLR21(C) mRNA in Pelteobagrus fulvidraco
![]()
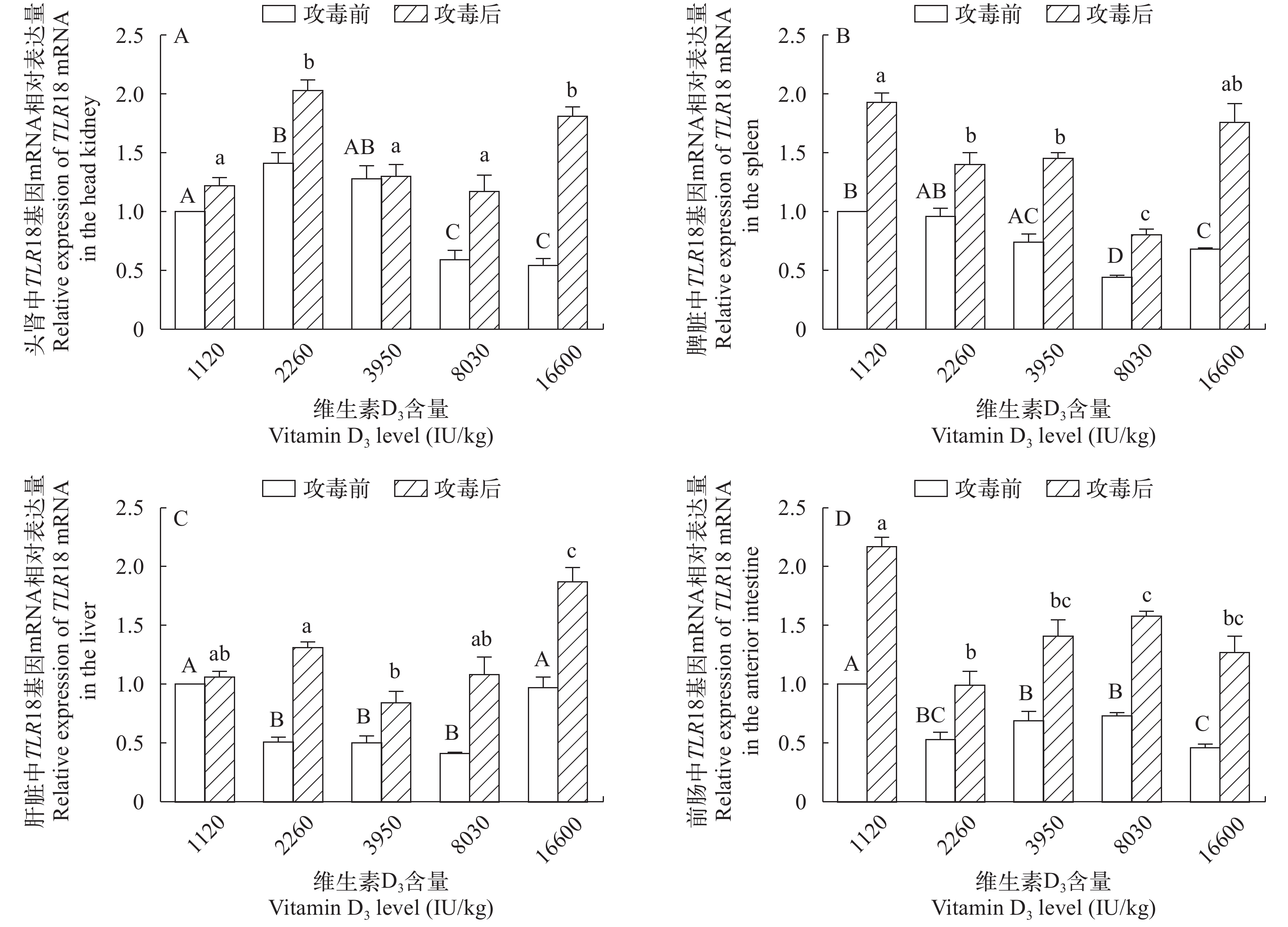
图 2 攻毒前后TLR18 mRNA在黄颡鱼头肾(A)、脾脏(B)、肝脏(C)和前肠(D)中相对表达量的变化
Figure 2. Changes in the relative mRNA expression levels of the TLR18 gene in the head kidney, spleen, liver and anterior intestine after the Edwardsiella ictaluri stimulation
![]()
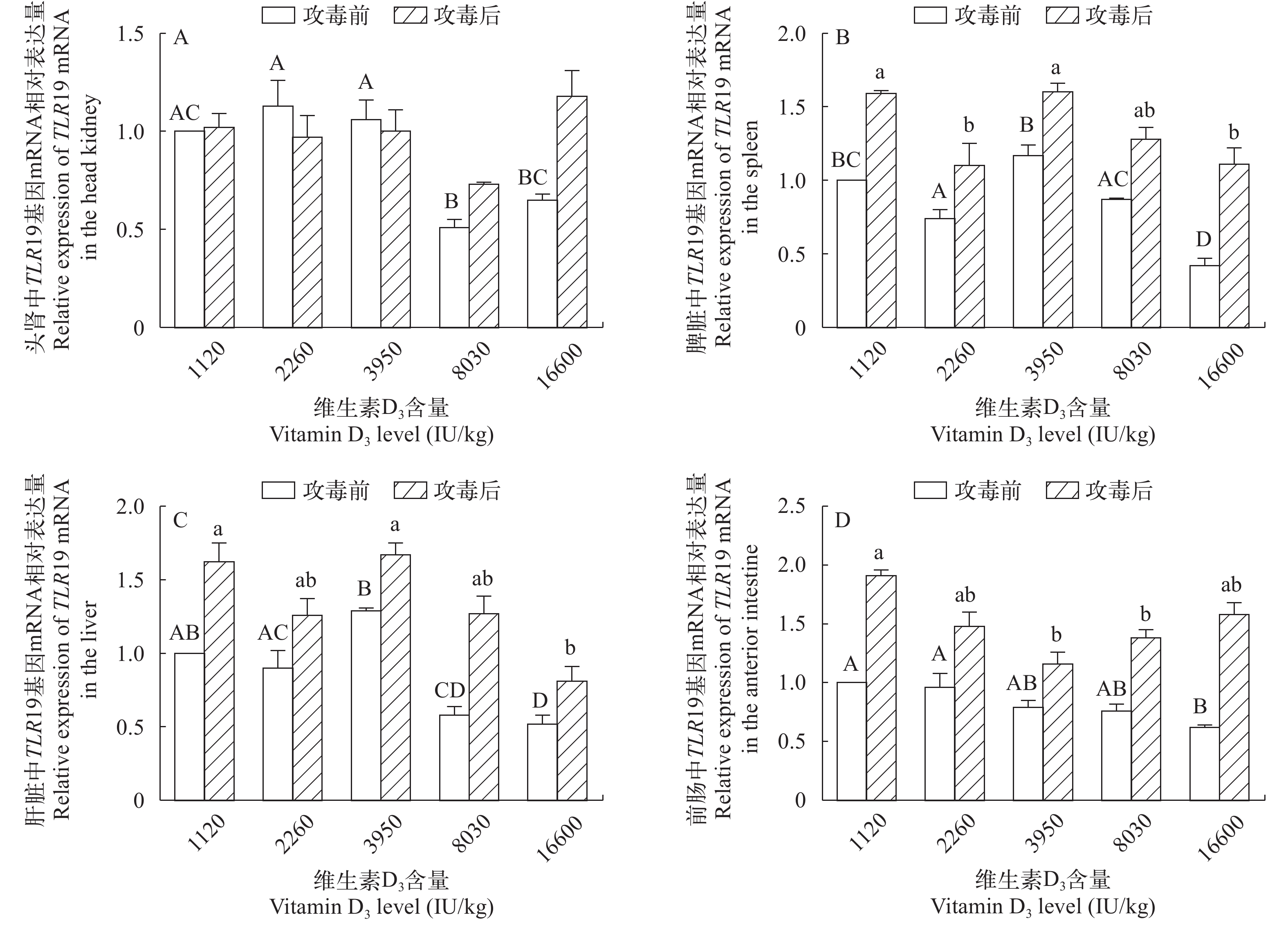
图 3 攻毒前后TLR19 mRNA在黄颡鱼头肾(A)、脾脏(B)、肝脏(C)和前肠(D)中相对表达量的变化
Figure 3. Changes in the relative mRNA expression levels of the TLR19 gene in the head kidney, spleen, liver and anterior intestine after the Edwardsiella ictaluri stimulation
![]()
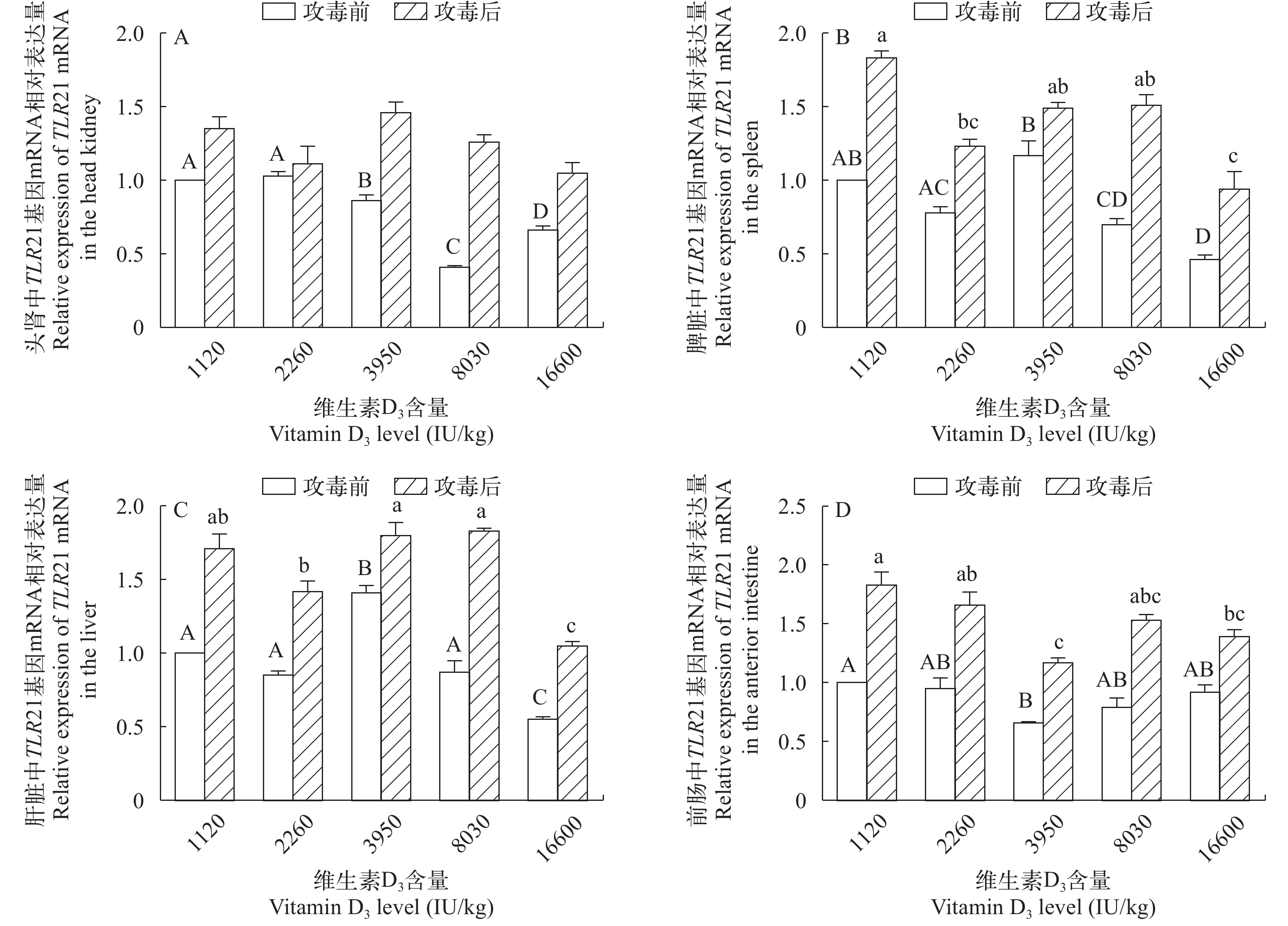
图 4 攻毒前后TLR21 mRNA在黄颡鱼头肾(A)、脾脏(B)、肝脏(C)和前肠(D)中相对表达量的变化
Figure 4. Changes in the relative mRNA expression levels of the TLR21 gene in the head kidney, spleen, liver and anterior intestine after the Edwardsiella ictaluri stimulation
表 1 实验饲料组成和营养成分
Table 1 Formulation and proximate composition of experimental diets
项目Item 饲料Ⅰ Diet I 饲料Ⅱ Diet II 饲料Ⅲ Diet III 饲料Ⅳ Diet IV 饲料Ⅴ Diet V 原料(%饲料干重)Ingredients (% dry matter) 血粉 10 10 10 10 10 脱脂奶粉 5 5 5 5 5 玉米蛋白粉 2 2 2 2 2 面粉 10 10 10 10 10 酪蛋白 30 30 30 30 30 豆粕 10 10 10 10 10 菜粕 10 10 10 10 10 大豆油 10 10 10 10 10 赖氨酸 1 1 1 1 1 蛋氨酸 0.1 0.1 0.1 0.1 0.1 氯化胆碱 0.5 0.5 0.5 0.5 0.5 维生素C 0.5 0.5 0.5 0.5 0.5 三氧化二钇 0.1 0.1 0.1 0.1 0.1 抗氧化剂 0.02 0.02 0.02 0.02 0.02 矿物盐 2 2 2 2 2 无VD3-维生素预混料 0.5 0.5 0.5 0.5 0.5 维生素D3 0.2 0.4 0.8 1.6 3.2 麸皮 8.08 7.88 7.48 6.68 5.08 营养成分(%饲料干重)Proximate composition (% dry matter) 水分 5.45 6.00 5.23 5.52 5.55 粗蛋白 47.60 47.25 46.44 46.96 46.12 粗脂肪 4.53 4.57 4.56 4.40 4.48 灰分 3.30 3.45 3.47 3.36 3.37 维生素D 1120 2260 3950 8030 16600 注: (1) 矿物盐预混料(每千克饲料): 氯化钠198 mg; 硫酸镁2995 mg; 硫酸钠4560 mg; 硫酸钾4098.5 mg; 氯化钙2340 mg; 硫酸亚铁499.5 mg; 乳酸钙700.5 mg; 硫酸锌70.6 mg; 硫酸镁32.5 mg; 硫酸铜6.2 mg; 硫酸钴0.2 mg; 碘化钾0.6 mg; 麸皮4498.4 mg; (2) 无VD3的维生素预混物(每千克饲料): 维生素A 2750 IU, 维生素E 25 IU; 维生素K3 5 mg, 维生素B1 10 mg; 维生素B6 10 mg; 维生素B12 10 mg; 尼克酸50 mg; 核黄素10 mg; 泛酸钙25 mg; 叶酸5 mg; 肌醇50 mg; 抗坏血酸50 mg; 脱脂米糠1757.43 mg. 由广州市联鲲生物科技有限公司定制; (3) 维生素D3预混物: 包膜饲料级别的维生素D3 (500000 IU/kg). 由广州市联鲲生物科技有限公司提供Note: (1) Mineral premix (mg per kg diet): NaCl 198 mg; MgSO4·7H2O 2995 mg; Na2SO4 4560 mg; K2SO4 4098.5 mg; CaCl2·2H2O 2340 mg; FeSO4 499.5 mg; Calcium lactate 700.5 mg; ZnSO4·7H2O 70.6 mg; MnSO4·4H2O 32.5 mg; CuSO4·5H2O 6.2 mg; CoSO4 0.2 mg; KI 0.6 mg; Bran 4498.4 mg; (2) Vitamin premix (per kg diet): Vitamin A 2750 IU; Vitamin E 25 IU; Vitamin K3 15 mg; Vitamin B1 20 mg; Vitamin B6 20 mg; Vitamin B12 20 mg; Niacin 50 mg; Riboflavin 10 mg; D-calcium pantothenate 25 mg; Biotin 5 mg; Inositol 100 mg; Ascorbic acid 500 mg; Bran 1757.43 mg. This premix was specially made by Nutriera; (3) Vitamin D3 premix: Coated vitamin D3 (500000 IU/kg), which was provided by Nutriera  下载: 导出CSV
下载: 导出CSV
表 2 Real-time PCR所用引物
Table 2 Primers used for real-time PCR
名称Name 序列Sequence (5′-3′) TLR18 Forward GGCACAGCTAAGGAGTGTCATGG TLR18 Reverce GGCCAGCTCAATCACAGACAGAG TLR19 Forward TCCGAGGCTGGAGGAACAAGG TLR19 Reverce GCACCAGTCGCTTCGCAGATAC TLR21 Forward CCAGGCCGCAACATAGTCGATAAC TLR21 Reverce AGCACCACTCACTACGGAGGAAG β-actin Forward GGATTCGCTGGAGATGATG β-actin Reverce TCGTTGTAGAAGGTGTGATG
下载: 导出CSV
表 3 在饲料中添加维生素D3对黄颡鱼幼鱼生长的影响
Table 3 Effects of dietary vitamin D3 levels on the growth performance of juvenile P.fulvidraco
指标Index 饲料维生素D3含量Dietary vitamin D3 level (IU/kg) 1120 2260 3950 8030 16600 初始体重IBW (g) 5.53±0.08 5.51±0.07 5.55±0.03 5.56±0.06 5.32±0.15 终末体重FBW (g) 26.15±0.63 24.00±0.66 24.93±0.60 25.91±0.99 26.99±1.37 增重率WG (%) 373.2±9.2ab 335.9±12.6b 349.2±11.6ab 365.5±16.0ab 408.3±27.0a 热积温生长系数TGC (%) 0.060±0.001 0.056±0.001 0.058±0.001 0.060±0.002 0.063±0.003 饲料效率FE (%) 0.63±0.02ab 0.58±0.01b 0.62±0.02ab 0.65±0.35ab 0.71±0.42a 饲料系数FCR 1.60±0.05ab 1.74±0.28b 1.63±0.05ab 1.55±0.08ab 1.43±0.09a 存活率SUR (%) 93.33±1.36 95.00±0.96 95.00±0.96 95.84±0.84 95.84±0.84 注: 同一行中数值后的上标字母不同代表差异显著(P<0.05)Note: Means with different letters in the same row are significantly different (P<0.05)
下载: 导出CSV
表 4 在饲料中维生素D3添加水平对黄颡鱼疾病抗性的影响
Table 4 Effects of vitamin D3 supplementation on the disease resistance of P. fulvidraco
项目Item 时间Time (d) 饲料维生素D3含量Dietary vitamin D3 level (IU/kg) 1120 2260 3950 8030 16600 攻毒后死亡数(尾)Death number after challenge tail 1 12 10 9 8 9 2 5 5 5 4 3 3 2 2 1 2 1 累积死亡总数(尾)Cumulative death number tail 19 17 15 14 13 累积死亡率Cumulative mortality (%) 34.54 30.90 27.27 25.45 23.63 免疫保护率Immune protective rate (%) 10.54 21.05 26.32 31.59
下载: 导出CSV
-
[1] 张铜, 徐奇友. 鱼类对维生素D需要量的研究进展 [J]. 饲料研究, 2010, (11): 62-65. Zhang T, Xu Q Y. Advances in research on vitamin D requirements in fish [J]. Feed Research, 2010, (11): 62-65.
[2] Mostafa W Z, Hegazy R A. Vitamin D and the skin: focus on a complex relationship: A review [J]. Journal of Advanced Research, 2015, 6(6): 793-804. doi: 10.1016/j.jare.2014.01.011
[3] Lock E J, Waagb R, Bonga S W, et al. The significance of vitamin D for fish: a review [J]. Aquaculture Nutrition, 2010, 16(1): 100-116. doi: 10.1111/j.1365-2095.2009.00722.x
[4] Calton E K, Keane K N, Newsholme P, et al. The impact of vitamin D levels on inflammatory status: a systematic review of immune cell studies [J]. PLoS One, 2015, 10(11): 141-147.
[5] Schwartz Z, Ehland H, Sylvia V L, et al. 1α, 25-dihydroxyvitamin D3 and 24 R, 25-dihydroxyvitamin D3 modulate growth plate chondrocyte physiology via protein kinase C-dependent phosphorylation of extracellular signal-regulated kinase 1/2 mitogen-activated protein kinase [J]. Endocrinology, 2002, 143(7): 2775-2786. doi: 10.1210/endo.143.7.8889
[6] Fleet J C. Rapid, membrane-initiated actions of 1,25-dihydroxyvitamin D: what are they and what do they mean [J]? Journal of Nutrition, 2004, 134(12): 3215-3218. doi: 10.1093/jn/134.12.3215
[7] Beard J A, Bearden A, Striker R. Vitamin D and the antiviral state [J]. Journal of Clinical Virology, 2011, 50(3): 194-200. doi: 10.1016/j.jcv.2010.12.006
[8] 范泽军, 邹鹏飞, 姚翠鸾. 鱼类Toll样受体及其信号传导的研究进展 [J]. 水生生物学报, 2015, 39(1): 173-184. doi: 10.7541/2015.22 Fan Z J, Zou P F, Yao C L. Advances in fish Toll-like receptors and signal transduction [J]. Acta Hydrobiologica Sinica, 2015, 39(1): 173-184. doi: 10.7541/2015.22
[9] Kwmar H, Kuwai T, Akwa S. Toll-like receptors and innate immunity [J]. Biochemical and Biophysical Research Communications, 2009, 388(4): 621-625. doi: 10.1016/j.bbrc.2009.08.062
[10] Rebl A, Goldammer T, Seyfert H. Toll-like receptor signaling in bony fish [J]. Veterinary Immunology and Immunopathology, 2010, 134(3-4): 139-150. doi: 10.1016/j.vetimm.2009.09.021
[11] Taro K, Shizuo A. Pathogen recognition with Toll-like receptors [J]. Current Opinion in Immunology, 2005, 17(4): 338-344. doi: 10.1016/j.coi.2005.02.007
[12] 李春艳, 黄贝, 熊静, 等. 日本鳗鲡TLR21基因的鉴定、免疫应答与启动子分析 [J]. 水生生物学报, 2017, 41(2): 296-305. Li C Y, Huang B, Xiong J, et al. Molecular cloning and characterization of TLR21 gene from japanese eel, Anguilla japonica [J]. Acta Hydrobiologica Sinica, 2017, 41(2): 296-305.
[13] Takano T, Hwang S D, Kondo H, et al. Evidence of molecular Toll-like receptor mechanisms in teleosts [J]. Fish Pathology, 2010, 245(1): 1-16.
[14] 沈建忠. 黄颡鱼的生物学特性及其养殖技术 [J]. 养殖与饲料, 2002, (3): 25-28. Shen J Z. Biological characteristics and farming technology of Pelteobagrus fulvidraco [J]. Aquaculture and Feed, 2002, (3): 25-28.
[15] 隗黎丽, 吴华东, 阮记明. 黄颡鱼鮰爱德华氏菌的分离及鉴定 [J]. 江西农业大学学报, 2014, 36(1): 187-192. doi: 10.3969/j.issn.1000-2286.2014.01.029 Kui L L, Wu H D, Ruan J M, et al. Isolation and identification of the Edwardsiella ictaluri in Pelteobagrus fulvidraco [J]. Journal of Jiangxi Agricultural University, 2014, 36(1): 187-192. doi: 10.3969/j.issn.1000-2286.2014.01.029
[16] Zuo R T, Ai Q H, Mai K S, et al. Effects of dietary N3- highly unsaturated fatty acids on growth, nonspecific immunity, expression of some immune related genes and disease resistance of large yellow croaker (Larmichthys crocea) following natural infestation of parasites (Cryptocaryon irritans) [J]. Fish & Shellfish Immunology, 2012, 32(2): 249.
[17] Zhao F, Li Y W, Pan H J, et al. Expression profiles of toll-like receptors in channel catfish (Ictalurus punctatus) after infection with Ichthyophthirius multifiliis [J]. Fish & Shellfish Immunology, 2013, 35(6): 993-997.
[18] Lee P T, Zou J, Holland J W, et al. Identification and characterization of TLR18-21 genes in Atlantic samlon (Salmo salar) [J]. Fish & Shellfish Immunology, 2014, 41(2): 549-555.
[19] Wang W, Shen Y, Pandit N P, et al. Molecular cloning characterization and immunological response analysis of Toll-like receptor (TLR21) gene in grass carp, Ctenopharyngodon idella [J]. Developmental and Comparative Immunology, 2013, 40(3-4): 227-231. doi: 10.1016/j.dci.2013.03.003
[20] Wang K L, Ji W, Wei K J, et al. Molecular characterization and expression analysis of three TLR genes in yellow catfish (Pelteobagrus fulvidraco): Responses to stimulation of Aeromonas hydrophila and TLR ligands [J]. Fish & Shellfish Immunology, 2017, (66): 466-479.
[21] Osamu T, Shizuo A. Pattern recognition receptors and inflammation [J]. Cell, 2010, 140(6): 805-820. doi: 10.1016/j.cell.2010.01.022
[22] Palti Y. Toll-like receptors in bony fish: from genomics to function [J]. Developmental and Comparative Immunology, 2011, 35(12): 1263-1272. doi: 10.1016/j.dci.2011.03.006
[23] Zhang J, Kong X H, Zhou C J, et al. Toll-like receptor recognition of bacteria in fish: Ligand specificity and signal pathways [J]. Fish & Shellfish Immunology, 2014, 41(2): 380-388.
[24] Wang J L, Zhang Z, Fu H, et al. Structural and evolutionary characteristics of fish specific TLR19 [J]. Fish & Shellfish Immunology, 2015, 47(1): 271-279.
[25] Huang W J, Shen Y, Xu X Y, et al. Identification and characterization of the TLR18 gene in grass carp, Ctenopharyngodon idella [J]. Fish & Shellfish Immunology, 2015, (47): 681-688.
[26] Zhao F, Li Y W, Pan H J, et al. Expression profiles of toll-like receptors in channel catfish (Ictalurus punctatus) after infection with Ichthyophthirius multifiliis [J]. Fish & Shellfish Immunology, 2013, 35(3): 993-997.
[27] Gao H, Wu L, Sun J S, et al. Molecular characterization and expression analysis of Toll-like receptor 21 cDNA from Paralichthys olivaceus [J]. Fish & Shellfish Immunology, 2013, (35): 1138-1145.
[28] Li Y W, Luo X C, Dan X M, et al. Molecular cloning of orange-spotted grouper (Epinephelus coioides) TLR21 and expression analysis post Cryptocaryon irritans infection [J]. Fish & Shellfish Immunology, 2012, 32(3): 476-481.
[29] Bell J K, Mullen G E D, Leifer C A, et al. Leucine-rich repeats and pathogen recognition in Toll-like receptors [J]. Trends in immunology, 2003, 24(10): 528-533. doi: 10.1016/S1471-4906(03)00242-4
[30] Zhang J, Liu S, Liu Z, et al. Pathogen recognition receptors in channel catfish: Ⅲ phylogeny and expression analysis of Toll-like receptors [J]. Developmental and Comparative Immunology, 2013, 40(1): 185-194.
[31] Naim M, Maalouf M D. The noncalciotropic actions of vitamin D3 recent clinial developments [J]. Current Opinion in Nephrology and Hypentension, 2008, 17(4): 408-415. doi: 10.1097/MNH.0b013e3283040c99
[32] Rebeca C, Alberto C, Lee J, et al. Effects of dietary vitamin D3 administration on innate immune parameters of seabream (Sparus aurata L) [J]. Fish & Shellfish Immunology, 2009, 26(2): 243-248.
[33] 黎德兵, 李超, 周定刚, 等. 饲料中维生素D3水平对黄鳝抗菌肽hepcidin基因表达的影响 [J]. 动物营养学报, 2014, 24(8): 1520-1526. Li D B, Li C, Zhou D G, et al. Effects of dietary vitamin D3 level on expression of antimicrobial peptide hepcidin gene in Monopterus albus [J]. Chinese Journal of Animal Nutrition, 2014, 24(8): 1520-1526.
[34] Cerezuela R, Cuesta A, Meseguer J, et al. Effects of dietary vitamin D3 administration on innate immune parameters of seabream (Sparus aurata L) [J]. Fish & Shellfish Immunology, 2008, 26(2): 243-245.
-
期刊类型引用(28)
1. 孙亚乔,蒋婕,段磊,杨思月,周艾诗,朱思衡,曹永翔,孙佳乾,寇思飞. 浮游生物对渭河水环境的响应机理及生态效应. 水利水电技术(中英文). 2024(04): 121-136 .  百度学术
百度学术
2. 李刚,潘保柱,丁一桐,孙长顺,王晓涛,杨子杰. 无定河流域浮游动物群落结构特征及水质生物评价. 中国环境监测. 2024(03): 181-196 . 百度学术
3. 胡威,谢蓉蓉,李家兵,蔡金傍,赵志强. 春夏过渡期调度型水库“河-库-河”连续体浮游生物群落结构的联动变化及多样性评价. 福建轻纺. 2024(09): 10-17 . 百度学术
4. 刘玉飞,葛丹,施银燕,蓝春燕,施思,殷燕,许海. 分水江流域秋季浮游动物群落特征分析. 环境生态学. 2024(09): 55-59+64 . 百度学术
5. 吕梦茹,丁奕帆,张瑞,刘洋,阚冬奇,张真,刘其根,杨家新. 紧水滩水库后生浮游动物群落结构及水质营养状态评价. 长江流域资源与环境. 2023(02): 354-364 . 百度学术
6. 张浩坤,闵奋力,崔慧荣,彭雪,张心怡,张淑娴,李竹栖,葛芳杰,张璐,吴振斌,刘碧云. 武汉市3种类型湖泊浮游植物群落特点及关键影响因子. 环境科学. 2023(04): 2093-2102 . 百度学术
7. 赵晨,孙佳伟,尹增强,王慧博,王乐,宋聃,都雪,赵玉财,孙洋,霍堂斌. 大罗圈沟河水生生物多样性及水质评价. 大连海洋大学学报. 2023(03): 504-514 . 百度学术
8. 徐珊,于莉,徐秋云,陈聪,白海锋. 济南市雪野湖浮游生物群落结构特征及多样性分析. 安徽农业科学. 2023(24): 56-61 . 百度学术
9. 白海锋,王怡睿,宋进喜,孔飞鹤,张雪仙,李琦. 渭河浮游生物群落结构特征及其与环境因子的关系. 生态环境学报. 2022(01): 117-130 . 百度学术
10. 刘俏,刘智暘,王江滨,廖传松,李俊锋,郭超,王靖斌,熊满堂,张堂林,郭传波,刘家寿. 福建山美水库浮游动物群落结构时空特征及其影响因子分析. 湖泊科学. 2022(06): 2039-2057 . 百度学术
11. 孙枭琼,崔铁峰,方楠,王印开,王宏伟. 白洋淀枝角类和桡足类调查及水环境因子分析. 河北大学学报(自然科学版). 2021(03): 290-296 . 百度学术
12. 陈佳林,余海军,王茜. 大清河流域浮游动物的群落特征研究. 水生态学杂志. 2021(03): 72-78 . 百度学术
13. 徐雪,姚文佳,邢雨辉,张振华,时应征,胡超超,常青,陈建琴. 苏州工业园区湖泊后生浮游动物群落结构及影响因子. 生态学报. 2021(10): 4023-4035 . 百度学术
14. 王腾,刘永,全秋梅,林琳,肖雅元,李纯厚,李鸿. 广东江门市主要淡水河流浮游动物群落结构特征. 南方水产科学. 2021(04): 9-17 . 百度学术
15. 梁建,路伟亭. 涡河中下游浮游动物群落结构及水生态评价. 水生态学杂志. 2021(04): 50-58 . 百度学术
16. 杨丹丹,李君轶,吴金明,王丰,问思恩,熊冬梅,邵俭. 千河流域秋季水生生物资源调查研究. 水产学杂志. 2021(04): 49-58 . 百度学术
17. 白海锋,孔飞鹤,王怡睿,宋进喜,曹叶琳,蒋晓辉. 北洛河流域浮游动物群落结构时空特征及其与环境因子相关性. 大连海洋大学学报. 2021(05): 785-795 . 百度学术
18. 严航,夏霆,陈宇飞,姚春雨,王浩. 太湖流域平水期水生态功能区浮游动物群落结构特征. 长江流域资源与环境. 2021(11): 2641-2650 . 百度学术
19. 相华,朱中竹,商书芹,殷旭旺,白海锋. 小清河济南段浮游动物群落结构空间变化特征. 河北渔业. 2020(01): 38-43 . 百度学术
20. 汪中华,王帅帅,韩晓,殷旭旺,白海锋. 小清河济南段浮游动物多样性及水质评价. 黑龙江水产. 2020(01): 18-22 . 百度学术
21. 陆颖,陈宇顺. 河流生境的评估方法及应用的研究进展. 水生生物学报. 2020(03): 670-684 . 本站查看
22. 王雨路,袁丹妮,袁国庆,冯伟松,龚迎春. 武汉东湖夏冬两季浮游动物物种多样性及群落结构研究. 水生生物学报. 2020(04): 877-894 . 本站查看
23. 丁婷婷,杜士林,王宏亮,张亚辉,王一喆,何连生. 沙颍河流域硝基苯水质基准推导及生态风险评估. 生态毒理学报. 2020(04): 256-267 . 百度学术
24. 卫向东,郭匿春,丁瑞睿,李静. 巢湖双桥河底泥疏浚过程中轮虫群落结构研究. 环境科学与技术. 2020(09): 190-197 . 百度学术
25. 刘麟菲,徐宗学,殷旭旺,李福林,王汨. 济南市不同区域水生生物与水环境因子的响应关系. 湖泊科学. 2019(04): 998-1011 . 百度学术
26. 王硕,杨涛,李小平,陈佳. 渭河流域浮游动物群落结构及其水质评价. 水生生物学报. 2019(06): 1333-1345 . 本站查看
27. 林海,王源,李冰. 北京市妫水河浮游动物群落结构与水质评价. 生态学报. 2019(20): 7583-7591 . 百度学术
28. 毕孟飞,虞洋,陈佳琪,赵坤,涂俊. 夏季率水浮游动物群落类型及影响因素. 上海师范大学学报(自然科学版). 2019(05): 511-520 . 百度学术
其他类型引用(19)





计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 47