
IDENTIFICATION AND DENITRIFICATION CHARACTERISTICS OF THE AEROBIC DENITRIFYING BACTERIA PSEUDOMONAS FURUKAWAII ZS1
-
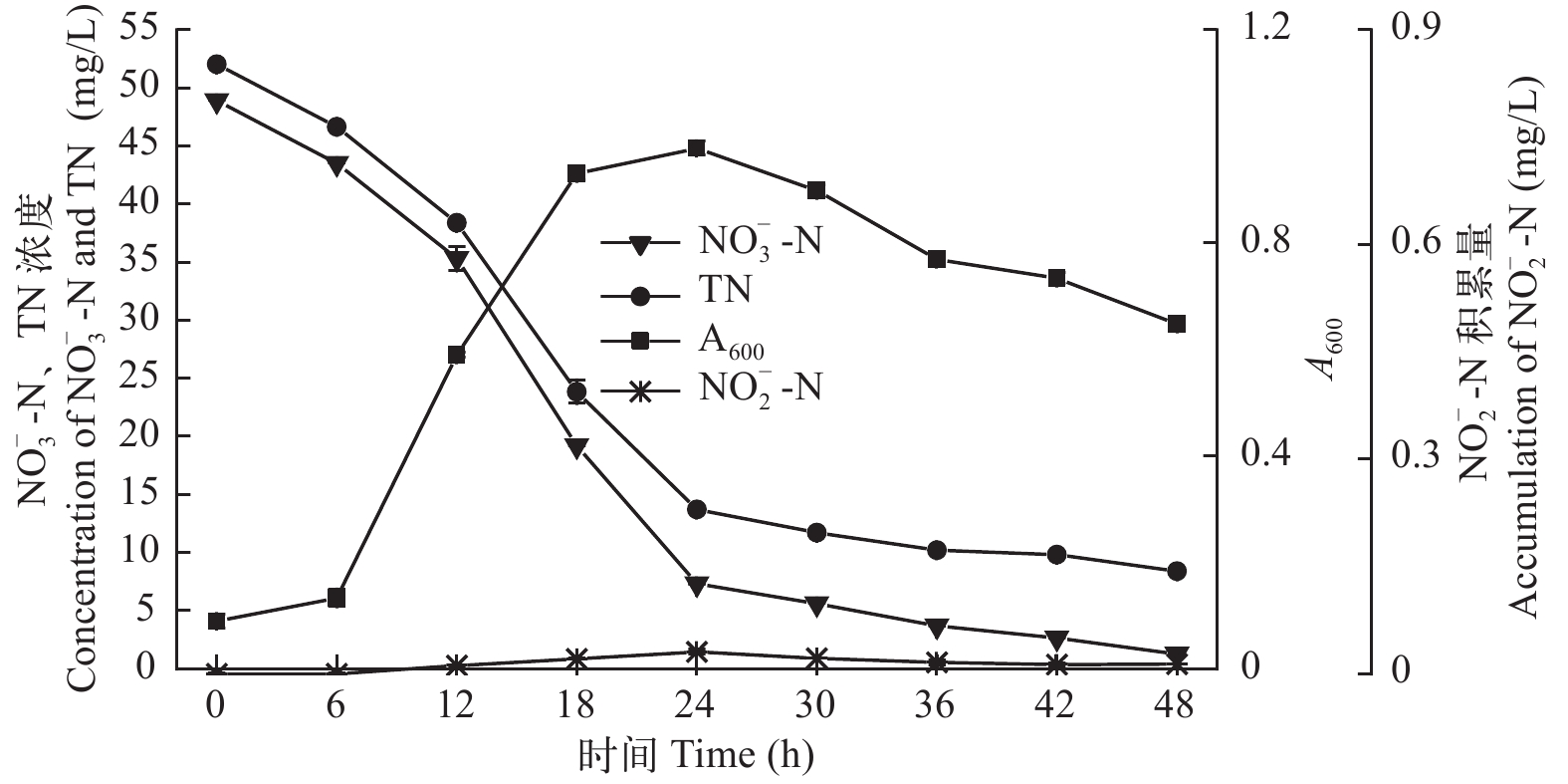
摘要: 2018年3月, 在广东省中山市的草鱼(Ctenopharyngodon idellus)养殖池塘水体中分离筛选出一株高效好氧反硝化细菌, 结合菌株的形态观察、生理生化特性和16S rDNA基因序列分析鉴定为Pseudomonas furukawaii, 命名为ZS1。并进一步研究了该菌株的脱氮特性, 同时采用单因素实验方法探究了不同碳源种类、温度、pH、C/N和摇床转速对菌株ZS1脱氮效率的影响。结果表明, ZS1菌株在好氧条件下具有高效的脱氮效率。经过48h的培养, 硝酸盐氮(
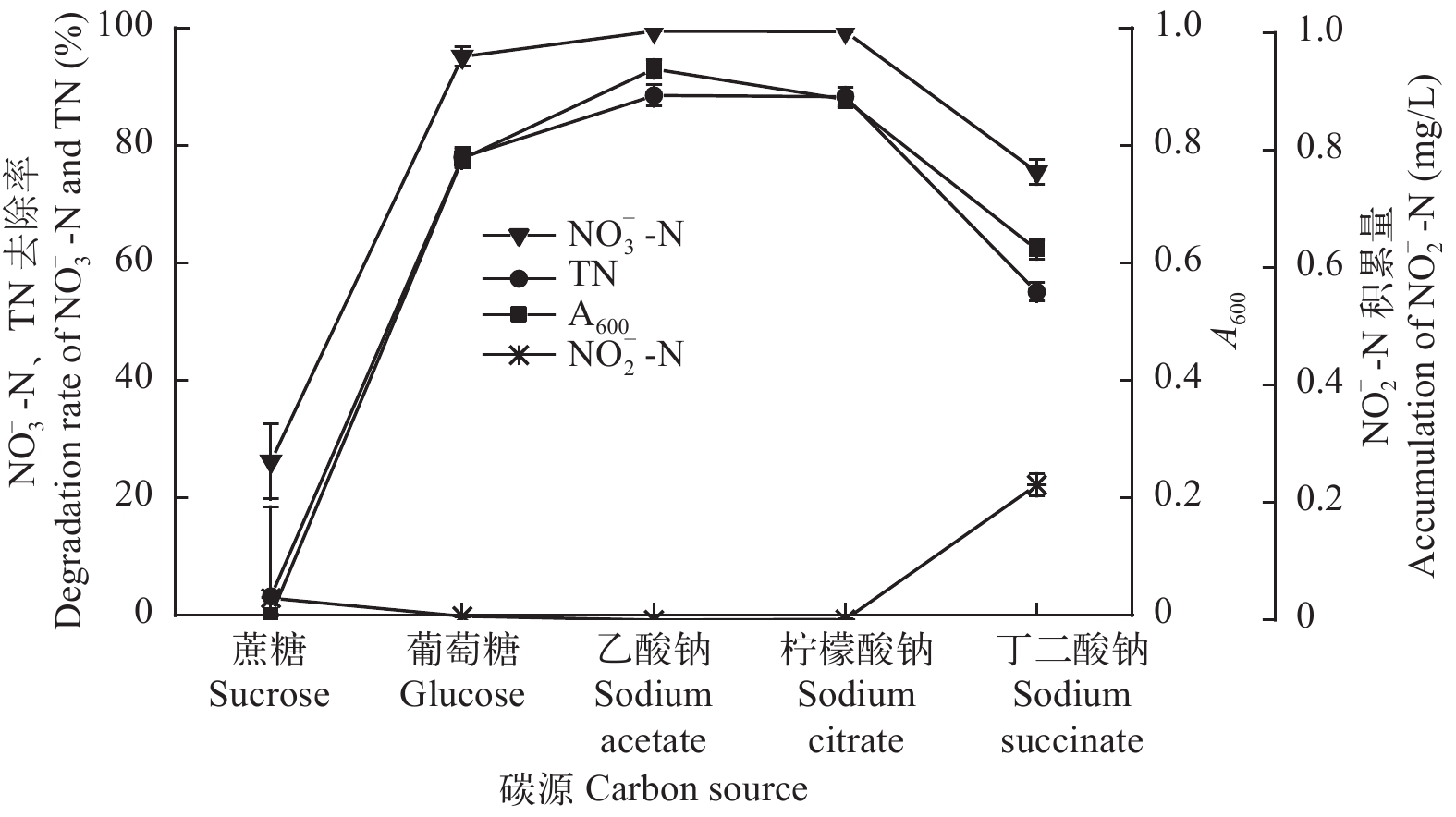
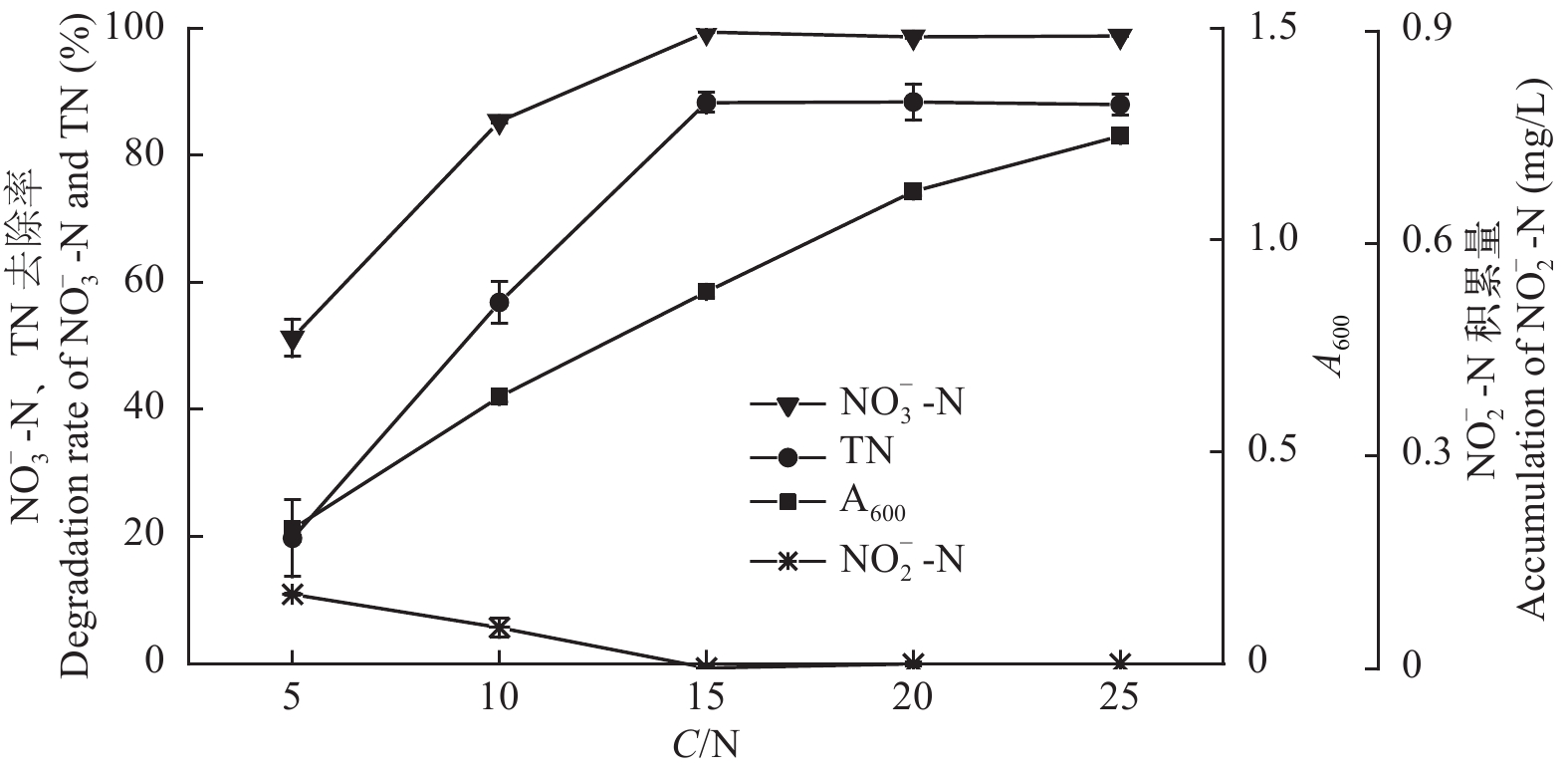
$ {\rm{NO}}_{\rm{3}}^{\rm{ - }}$ -N)的含量从48.93降低到1.27 mg/L, 去除率为97.40%, 去除速率达0.993 mg/(L·h); 总氮(TN)的含量从52.04降低到8.40 mg/L, 去除率为83.86%, 去除速率达0.909 mg/(L·h), 且无亚硝酸盐氮($ {\rm{NO}}_{\rm{2}}^{\rm{ - }}$ -N)积累。以$ {\rm{NO}}_{\rm{3}}^{\rm{ - }}$ -N为唯一氮源, 菌株ZS1发挥最佳好氧反硝化性能的碳源为乙酸钠、柠檬酸钠和葡萄糖、温度为25—35℃、pH为7.0—10.0、C/N为15—25、转速为100—200 r/min。上述结果显示, 菌株P. furukawaii ZS1具有良好的好氧反硝化性能, 将为池塘养殖尾水处理应用生物脱氮技术提供理论依据及成为初步的候选菌株。Abstract: An efficient aerobic denitrifying bacteria species was isolated from water in a Ctenopharyngodon idellus culture pond in Zhongshan City, Guangdong Province in March 2018. The bacteria were identified as Pseudomonas furukawaii by morphological observation, physiological and biochemical characteristics, and 16S rDNA sequence analysis. This strain was named Pseudomonas furukawaii ZS1. The denitrification characteristics of the strain and the effects of the carbon source, temperature, pH, C/N ratio and shaking speed on the denitrification ability of strain ZS1 were investigated. The results showed that P. furukawaii ZS1 was an efficient denitrifier under aerobic conditions. The content of nitrate nitrogen ($ {\rm{NO}}_{\rm{3}}^{\rm{ - }}$ -N) decreased from 48.93 mg/L to 1.27 mg/L, the removal efficiency of$ {\rm{NO}}_{\rm{3}}^{\rm{ - }}$ -N was 97.40%, and the removal rate was 0.993 mg/L/h; the content of total nitrogen (TN) decreased from 52.04 mg/L to 8.40 mg/L, the removal efficiency was 83.86%, and the removal rate was 0.909 mg/L/h. Single-factor experiments indicated that the optimal conditions for denitrification activity in P. furukawaii ZS1 were a carbon source of sodium acetate, sodium citrate or glucose; temperatures ranging from 25 to 35℃; a pH of 7.0 to 10.0; a C/N ratio of 15 to 25; and a shaking speed of 100 to 200 r/min. The above results showed that the strain could quickly and effectively remove nitrogen through aerobic denitrification. Therefore, the P. furukawaii ZS1 strain can be used as a candidate strain for the treatment of pond culture wastewater through biological denitrification. -
大银鱼(Protosalanx hyalocranius Abbott, 1901)和太湖新银鱼(Neosalanx taihuensis Chen, 1956)分别隶属于鲑形目(Salmoniformes)银鱼科(Salangidae)的大银鱼属和新银鱼属, 主要分布在我国黄河、淮河和长江中下游及其附属湖泊, 是我国重要的经济鱼类[1]。20世纪70年代以来, 2种银鱼被引种移植到全国众多湖泊和水库[2]。大银鱼主要被移植到长江以北[3], 太湖新银鱼则多被移植到黄河以南[4]。2种银鱼的生命周期均为一年, 个体小、生长快、世代离散, 种群波动显著 [1, 5]。
过度捕捞是影响鱼类资源可持续利用的重要因素, 分为生长型过度捕捞和补充型过度捕捞[6]。由于银鱼世代离散, 补充型过度捕捞导致剩余产卵群体数量不足, 进而使种群下一世代数量减少, 被认为是其资源量下降的重要原因[7]。单位补充量产卵群体生物量(Spawning Biomass-recruitment, SBR)模型根据种群的生长、死亡参数和种群资源动态, 估算捕捞后产卵群体的残存量, 进而评估种群的资源利用状况[8, 9]。当SBR残存量低于未开发状态的20%时, 通常视为补充型过度捕捞, 可能导致种群崩溃[10, 11]。平衡产量模型是渔业资源评估最常用的Beverton-Holt模型的简化形式[6, 12], 常用于分析开捕时间和捕捞强度对渔获产量的影响, 并据此制定合理的渔业管理方案[13, 14]。
洪泽湖(33°06—40′N, 118°10—52′E)是中国第四大淡水湖, 水域面积约2069 km2。大银鱼和太湖新银鱼是洪泽湖最主要的经济鱼类之一, 2010年以来年渔产量最高曾达106 kg[15], 但年际波动较大, 且近年来呈现下降趋势(江苏省洪泽湖渔业管理委员会办公室, 未发表数据)。洪泽湖银鱼捕捞采用银鱼拖网进行, 每年1月1日至8月8日为禁捕期, 其中5月21日左右允许捕捞1周。据渔民反映, 在每年12月下旬捕捞期的最后阶段2种银鱼均已很难捕捞、渔获量极低。本研究对洪泽湖大银鱼和太湖新银鱼的周年生长和资源量动态进行了分析, 评估种群捕捞利用状况, 进而优化渔业管理方式, 以期为洪泽湖银鱼的资源可持续利用管理提供参考。
1. 材料与方法
1.1 采样水域与采样方法
2015年7月至2016年6月每月中旬, 对洪泽湖大银鱼和太湖新银鱼进行周年采样(图 1)。采样使用渔民用的银鱼拖网进行, 拖网网口宽11.6 m, 高2.3 m; 网体长约25 m, 由4个网片组成, 侧面网片网目为1.2 cm×1.2 cm, 上下网片网目为1.5 cm×1.5 cm; 末端网袋长1.3 m, 网目为0.3 cm×0.3 cm。采样时在机动渔船左右两侧各设置一个拖网, 拖网上沿恰好没入水中, 渔船在固定起点(A, 33°17′19″N, 118°47′17″E)和终点(B, 33°19′58″N, 118°46′50″E)间拖行5 km, 拖速为4.0 km/h。采样在上午9:00—12:00间进行。样品采集后立即置于冰水保温箱中, 带回实验室计数, 进行生物学测定。每次采样2种银鱼生物学测定数量均在100尾以上, 当样品数量不足时, 增加拖行时间。
![]() 图 1 洪泽湖, 示大银鱼和太湖新银鱼采集时拖网拖行区间起点(A)和终点(B)Figure 1. Hongze Lake, showing the trawling area (from A to B) for P. hyalocranius and N. taihuensis collection
图 1 洪泽湖, 示大银鱼和太湖新银鱼采集时拖网拖行区间起点(A)和终点(B)Figure 1. Hongze Lake, showing the trawling area (from A to B) for P. hyalocranius and N. taihuensis collection1.2 数据获取与分析
体长-体重关系和生长率 在实验室对大银鱼和太湖新银鱼的渔获物进行计数和称重。测定个体体长(L, 0.1 mm), 称体重(W, 0.01 g), 解剖鉴定性别, 繁殖季节肉眼确定雌性性腺发育分期[16, 17]。体长-体重关系采用W=a×Lb进行拟合。采用单因子方差分析(ANOVA)检验各月2种银鱼体长和体重差异显著性, 并采用Bonferroni方法进行多重比较。分别求出2种银鱼各月的平均体长和体重, 计算相邻月份间体长和体重瞬时生长率(gl, gw)[18]:
${g_l}{\rm{ = ln}}{L_{n + 1}}{\rm{ - ln}}{L_n}$

${g_w}={\rm{ln}}{W_{n + 1}} - {\rm{ ln}}{W_n}$

式中, Ln 和Wn 分别为某月(n)大银鱼或太湖新银鱼的平均体长和体重; Ln+1 和Wn+1则为下一月份的平均体长和体重。
最适生长方程 洪泽湖大银鱼的繁殖季节为12至次年2月, 盛期为1月中旬; 太湖新银鱼春群的繁殖季节为3—4月, 盛期为3月(未发表数据)。据此, 我们将1月25日和3月25日分别设为大银鱼和太湖新银鱼的出生时间, 即0龄, 并推算2种银鱼各月采样时间对应的年龄。采用Logistic生长方程(Logistic growth function, Logistic GF)、Gompertz生长方程(Gompertz growth function, Gompertz GF)和von Bertalanffy生长方程(Von Bertalanffy Growth Function, VBGF)拟合2种银鱼的年龄与体长生长关系, 使用最小二乘法估算各生长方程的参数, 依据赤井信息准则(Akaike’s information criterion, AIC)选择最适生长方程[19]。3种生长方程分别为:
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
式中, t表示样本年龄; t0为理论生长起点, 即体长Lt=0时的起始年龄; L∞为渐近体长; K为生长系数。生长方程拟合采用R-3.3.1软件进行[20]。
种群死亡系数 定义单位捕捞努力量渔获量(Catch per Unit Effort, CPUE)为拖网拖行1 km2水域面积的渔获物个体数量和生物量。使用CPUE逐月数量变动数据, 结合洪泽湖当前的银鱼捕捞管理方式(8月9日—12月31日为捕捞期, 同时5月21—28日可对银鱼资源进行为期一周的捕捞), 求解以下方程组, 计算捕捞死亡系数F和自然死亡系数M [21]:
${N_{{{t_2}}}} = \left\{ {\begin{array}{*{20}{c}}\!\!\!\! {{N_{{{t_1}}}}{{\rm{e}}^{ - M({{t_2}} - {{t_1}})}}\;\;\left( \text{当不存在人为捕捞作用时} \right)}\\\!\!\!\! {{N_{{{t_1}}}}{{\rm{e}}^{ - Z({{t_2}} - {{t_1}})}}\;\;\left( \text{当存在人为捕捞作用时} \right)}\end{array}} \right.$

This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
模型构建 使用平衡产量模型(The equilibrium output model)估算种群单位补充量渔产量随开捕年龄和捕捞强度的变化[6, 13]:
$\begin{aligned}Y = & F \times {N_0} \times {{\rm{e}}^{ - M \times {{r}}}} \times {W_\infty } \times \\ & \left( {\frac{1}{Z} - \frac{{3{{\rm{e}}^{ - K \times {{r}}}}}}{{Z + K}} + \frac{{3{{\rm{e}}^{ - 2K \times {{r}}}}}}{{Z + 2K}} - \frac{{{{\rm{e}}^{ - 3K \times {{r}}}}}}{{Z + 3K}}} \right)\end{aligned}$

This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
使用SBR模型对洪泽湖当前银鱼捕捞管理方式和依据平衡产量模型制定的新的管理方式进行评估。假定2种方案下银鱼的捕捞死亡系数和自然死亡系数不变, 将模型中的时间步长设置为1周, 即0.0192 year[10], SBR模型公式为[8]:
$S \! B \! R \!\!=\!\! \sum\limits_{t = 0}^{{t_{\max }}} \!{\exp [( \!-\! (F \! \times \!\! {S_t}} \! \times \! {A_t}) \!-\! M) \!\! \times \!\! t] \!\! \times \!\! a \!\! \times \!\! {({L_t})^b} \!\! \times\! \! {G_t}$

式中, tmax表示样本的最大年龄; St和Gt分别表示刀刃式选择假设下的网具和性成熟选择性, 当t<50%性成熟体长时,Gt=0, 反之, Gt=1; 当t<初次开捕年龄时,St=0, 反之, St=1[22]; At对应于t龄时是否处于渔业开捕状态, 若是, 则At=1, 否则At=0。
对于SBR模型中使用的50%性成熟体长, 针对2种银鱼1年生且1次性成熟的特点, 以繁殖期性成熟的最小10尾雌性个体的体长均值Lm表示[23], 将其代入最适生长方程中, 计算对应的性成熟年龄Tm。
2. 结果
2.1 种群结构与生长
共测定了洪泽湖1250尾大银鱼和1818尾太湖新银鱼的体长和体重。大银鱼体长和体重分别为29.0—182.6 mm和0.10—34.79 g; 太湖新银鱼体长和体重分别为20.4—82.7 mm和0.04—3.40 g。银鱼的体长-体重关系方程分别为:
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
![]() 图 2 洪泽湖大银鱼(a)和太湖新银鱼(b)的体长频率分布逐月变化Figure 2. Monthly variations in body length frequency distribution for P. hyalocranius (a) and N. taihuensis (b) in Hongze Lake
图 2 洪泽湖大银鱼(a)和太湖新银鱼(b)的体长频率分布逐月变化Figure 2. Monthly variations in body length frequency distribution for P. hyalocranius (a) and N. taihuensis (b) in Hongze Lake大银鱼一个世代在渔获物中出现的时间从4月到翌年2月, 9月以后大银鱼个体间体长差异逐渐增大、体长范围逐渐增加, 12月后大个体在种群中所占比例下降(图 2a)。世代周期中存在4—6月和8—11月2个快速生长阶段。这2个阶段相邻月份的体长和体重均差异显著(P<0.05)。体长和体重均在11月达到最大值。体长和体重瞬时生长率总体均呈下降趋势, 12月后为负值(图 3、图 4)。
太湖新银鱼一个世代在渔获物中出现的时间从5月到翌年4月, 为春季繁殖群体(春群)的后代; 9月也采集到少量性成熟个体(秋群), 其后代于10月开始出现在渔获物中, 但数量较少(图 2b)。本研究仅考虑春群。太湖新银鱼整个世代周期中持续生长, 无明显的阶段性。5—12月间相邻月份间体长差异显著(P<0.05), 但体重差异不显著(P>0.05)。太湖新银鱼体长和体重瞬时生长率变化较大银鱼平稳, 总体呈下降趋势, 12月后近乎于零(图 3、图 4)。
![]() 图 3 洪泽湖大银鱼和太湖新银鱼(不包括秋群)的体长与其瞬时生长率的逐月变化(误差线代表标准差; *表示两个相邻月份间体长差异显著, P<0.05)Figure 3. Monthly fluctuations in body length and its instantaneous growth rate for P. hyalocranius and N. taihuensis in Hongze Lake (Error bars represent ± standard deviation; * indicates significant difference in body length between two adjacent months, P<0.05)
图 3 洪泽湖大银鱼和太湖新银鱼(不包括秋群)的体长与其瞬时生长率的逐月变化(误差线代表标准差; *表示两个相邻月份间体长差异显著, P<0.05)Figure 3. Monthly fluctuations in body length and its instantaneous growth rate for P. hyalocranius and N. taihuensis in Hongze Lake (Error bars represent ± standard deviation; * indicates significant difference in body length between two adjacent months, P<0.05)![]() 图 4 洪泽湖大银鱼和太湖新银鱼(不包括秋群)的体重与其瞬时生长率的逐月变化(误差线代表标准差; *表示两个相邻月份间体重差异显著, P<0.05)Figure 4. Monthly fluctuations in body weight and its instantaneous growth rate for P. hyalocranius and N. taihuensis in Hongze Lake (Error bars represent ± standard deviation; * indicates significant difference in body weight between two adjacent months, P<0.05)
图 4 洪泽湖大银鱼和太湖新银鱼(不包括秋群)的体重与其瞬时生长率的逐月变化(误差线代表标准差; *表示两个相邻月份间体重差异显著, P<0.05)Figure 4. Monthly fluctuations in body weight and its instantaneous growth rate for P. hyalocranius and N. taihuensis in Hongze Lake (Error bars represent ± standard deviation; * indicates significant difference in body weight between two adjacent months, P<0.05)2.2 最适生长方程与性成熟大小
在3种生长方程中, VBGF对大银鱼生长拟合的AIC值最小, Logistic GF对太湖新银鱼生长拟合的AIC值最小(表 1), 分别为2种鱼类的最适生长方程(图 5)。大银鱼和太湖新银鱼卵巢发育至Ⅳ期的10尾最小个体的体长均值Lm分别为99.9和61.1 mm, 代入对应的生长方程, 得出大银鱼和太湖新银鱼的Tm分别为0.528 year和0.564 year。
表 1 三种生长方程对大银鱼和太湖新银鱼生长拟合的方程参数Table 1. Estimated parameters of three different growth functions for P. hyalocranius and N. taihuensis in Hongze Lake参数Parameter 大银鱼P. hyalocranius 太湖新银鱼N. taihuensis Logistic GF Gompertz GF VBGF Logistic GF Gompertz GF VBGF L∞ (mm) 145.86 153.63 173.35 66.82 67.24 67.87 K 5.013 3.536 1.972 5.386 4.569 3.748 t0 (year) 0.376 0.290 0.092 0.124 0.054 –0.042 AIC 7237.47 7220.83 7202.66 12424.96 12428.04 12432.73 ![]() 图 5 洪泽湖大银鱼(a)和太湖新银鱼(b)的最适生长方程曲线Figure 5. The most fitted growth functions for P. hyalocranius (a) and N. taihuensis (b) in Hongze Lake
图 5 洪泽湖大银鱼(a)和太湖新银鱼(b)的最适生长方程曲线Figure 5. The most fitted growth functions for P. hyalocranius (a) and N. taihuensis (b) in Hongze Lake2.3 CPUE逐月变动与种群死亡系数
单位水域面积(1 km2)中大银鱼和太湖新银鱼渔获物数量均在5月最高, 分别为31809和78098 ind./km2。经历了5月中旬为期1周的捕捞后, 6月2种银鱼的资源数量均明显下降; 6月至8月间下降相对平缓; 8月捕捞开始后下降明显, 到12月时渔获物数量均降至500 ind./km2以下。单位水域面积(1 km2)大银鱼和太湖新银鱼渔获物生物量的最高值分别出现在6月和8月, 为73.07和49.33 kg/km2。大银鱼生物量在8—9月之间下降最快,太湖新银鱼生物量在9—10月间下降最快。
根据洪泽湖现行的捕捞管理方式, 5—6月间和8月以后, 2种银鱼的CPUE下降受捕捞和自然死亡的共同影响; 6—8月间, CPUE的下降只受自然死亡的影响。据此求出大银鱼F和M分别为8.583/year和3.292/year,太湖新银鱼F和M分别为7.006/year和1.146/year。
![]() 图 6 洪泽湖大银鱼和太湖新银鱼(春群)单位水域面积(1 km2)渔获物数量(a)和生物量(b)的逐月变动4月之前2种银鱼的个体小、渔获数量少, 均未计算CPUEFigure 6. Monthly variations in number of individuals (a) and biomass (b) for P. hyalocranius and N. taihuensis harvested across an area of 1 km2 by trawl in Hongze LakeIn April, size of both species was small, and numbers of catch for both species was very low. Thus, data in April was not included
图 6 洪泽湖大银鱼和太湖新银鱼(春群)单位水域面积(1 km2)渔获物数量(a)和生物量(b)的逐月变动4月之前2种银鱼的个体小、渔获数量少, 均未计算CPUEFigure 6. Monthly variations in number of individuals (a) and biomass (b) for P. hyalocranius and N. taihuensis harvested across an area of 1 km2 by trawl in Hongze LakeIn April, size of both species was small, and numbers of catch for both species was very low. Thus, data in April was not included2.4 平衡产量模型分析
使用以上获得的生长、繁殖和死亡等相关参数(表 2), 分别构建了大银鱼和太湖新银鱼单位补充量渔产量随开捕年龄(图 6a)和捕捞死亡系数(图 6b)变化的曲线。结果显示, 大银鱼和太湖新银鱼的单位补充量渔产量均随开捕年龄的增大呈先升后降的倒U型曲线关系。当大银鱼开捕年龄tc=0.525 year时, 能够取得相应最大的单位补充量渔产量Ymax=1.078 kg, 开捕年龄过早或过晚均不能获得足够的渔产量。就洪泽湖当前渔业管理模式来说, 大银鱼的开捕年龄约为0.538 year, 略大于tc, 此时的单位补充量渔产量Y=1.077 kg, 接近Ymax。当太湖新银鱼开捕年龄tc=0.525 year时, 能够取得相应最大的单位补充量渔产量Ymax=0.684 kg。就当前渔业管理模式来说, 太湖新银鱼的开捕年龄约为0.365 year, 此时的单位补充量渔产量Y=0.607 kg<Ymax。将开捕时间向后推移20d (即8月29日), 即大银鱼和太湖新银鱼的开捕年龄分别为0.593 year和0.420 year时, 能够获得最大的总单位补充量渔产量1.708 kg。
![]() 图 7 平衡产量模型估算的洪泽湖大银鱼和太湖新银鱼单位补充量渔产量随开捕年龄(a)和捕捞死亡系数(b)变化的曲线垂直虚线表示当前捕捞管理模式下2种银鱼的开捕年龄和捕捞死亡系数Figure 7. Relationships of yield per-recruitment to the age at first capture (a) and fishing mortality coefficient (b) estimated from the equilibrium output mode for P. hyalocranius and N. taihuensis in Hongze LakeThe dotted vertical lines locate the current age at first capture (a) and fishing mortality (b) for both species
图 7 平衡产量模型估算的洪泽湖大银鱼和太湖新银鱼单位补充量渔产量随开捕年龄(a)和捕捞死亡系数(b)变化的曲线垂直虚线表示当前捕捞管理模式下2种银鱼的开捕年龄和捕捞死亡系数Figure 7. Relationships of yield per-recruitment to the age at first capture (a) and fishing mortality coefficient (b) estimated from the equilibrium output mode for P. hyalocranius and N. taihuensis in Hongze LakeThe dotted vertical lines locate the current age at first capture (a) and fishing mortality (b) for both species大银鱼和太湖新银鱼的单位补充量渔产量均先随捕捞死亡系数的增加而快速增高, 当捕捞死亡系数增大到一定程度后, 渔产量上升速度减慢, 之后不再升高甚至出现下降趋势。大银鱼捕捞死亡系数F=24.38/year时, 获得最大单位补充量渔产量Ymax=1.114 kg; 当前洪泽湖大银鱼捕捞死亡系数为8.583/year, 相应渔产量Y=1.077 kg, 接近Ymax且尚未开发过度。太湖新银鱼捕捞死亡系数F=4.72/year时, 获得最大的单位补充量渔产量Ymax=0.616 kg; 而当前洪泽湖太湖新银鱼捕捞死亡系数为7.006/year, 已大于最适捕捞强度, 且相应的渔产量Y=0.607 kg, 小于Ymax。
2.5 基于SBR模型分析的资源状况评价
使用以上获得的生长、繁殖和死亡等相关参数(表 2), 分别构建了2种不同渔业管理方案下大银鱼和太湖新银鱼种群的SBR随捕捞死亡系数变动的曲线:
方案1: 洪泽湖现行的渔业管理方式。8月9日—12月31日为捕捞期, 同时5月可对银鱼资源进行为期1周的捕捞。
方案2: 假定对两种银鱼的捕捞强度不发生变化(即F恒定), 将开捕时间向后推迟20d, 8月29日—12月31日为捕捞期, 同时取消5月为期1周的捕捞。
大银鱼在当前渔业管理方案1下, 相对于未开发状态(100%), SBR残存量为20.234%, 即3.326 g, 接近于补充型过度捕捞评价阈值20%, 种群有可能因产卵亲体不足而导致资源衰退; 在方案2下, SBR残存量相当于未开发状态的36.721%, 即6.036 g, 远大于补充型过度捕捞评价阈值20%, 大银鱼渔业资源和产卵群体得到了较好的保护(图 7)。在2种不同管理方案下, 太湖新银鱼SBR残存量分别相当于未开发状态的7.503%和12.862%, 即0.972 g和1.666 g (图 8)。在优化的管理方案2下, 太湖新银鱼的产卵亲体资源虽然得到了一定程度的保护, 但其渔业资源仍处于补充型过度捕捞水平。
表 2 洪泽湖大银鱼和太湖新银鱼平衡产量模型和单位补充量产卵群体生物量模型参数Table 2. Growth and mortality parameters fitted into the equilib-rium output model and SBR model for P. hyalocranius and N. taihuensis in Hongze Lake参数
Parameter大银鱼
P. hyalocranius太湖新银鱼
N. taihuensisa 1.082×10–6 2.113×10–6 b 3.279 3.215 K 1.972 5.386 L∞ (mm) 173.35 66.82 W∞ (g) 23.75 1.56 t0 (year) 0.092 0.124 Z (/year) 11.875 8.152 M (/year) 3.292 1.146 F (/year) 8.583 7.006 tmax (year) 1 1 Lm (mm) 99.9 61.1 Tm (year) 0.528 0.564 tc (year) 0.538 0.365 ![]() 图 8 两种不同捕捞管理方案下洪泽湖大银鱼(a)和太湖新银鱼(b)单位补充量产卵群体生物量随捕捞死亡系数F变化的曲线当F=0时, 大银鱼和太湖新银鱼的单位补充量产卵群体生物量分别为16.44 g和12.95 g。捕捞管理方案1: 洪泽湖现行的渔业管理方式, 即8月9日—12月31日为捕捞期, 同时5月可对银鱼资源进行为期1周的捕捞; 捕捞管理方案2: 假定对两种银鱼的捕捞强度不变, 将开捕时间向后推迟20d, 即8月29日—12月31日为捕捞期, 同时取消5月为期1周的捕捞。虚线示两种管理方案下对应的捕捞死亡系数Figure 8. Relationships of spawner biomass per-recruitment to fishing mortality coefficients for P. hyalocranius (a) and N. taihuensis (b) under two different fishery management schemes in Hongze LakeThe spawner biomass per-recruitment at F=0 is 16.44 g for P. hyalocranius and 12.95 g for N. taihuensis. Fishery management scheme 1 is the current status, with fishing season of one week in Mid-May and between August 9 and December 31; the scheme 2 is an adjusted status, with fishing season between August 29 and December 31. The dotted vertical lines represent the corresponding fishing mortality coefficients under these two fishery management schemes
图 8 两种不同捕捞管理方案下洪泽湖大银鱼(a)和太湖新银鱼(b)单位补充量产卵群体生物量随捕捞死亡系数F变化的曲线当F=0时, 大银鱼和太湖新银鱼的单位补充量产卵群体生物量分别为16.44 g和12.95 g。捕捞管理方案1: 洪泽湖现行的渔业管理方式, 即8月9日—12月31日为捕捞期, 同时5月可对银鱼资源进行为期1周的捕捞; 捕捞管理方案2: 假定对两种银鱼的捕捞强度不变, 将开捕时间向后推迟20d, 即8月29日—12月31日为捕捞期, 同时取消5月为期1周的捕捞。虚线示两种管理方案下对应的捕捞死亡系数Figure 8. Relationships of spawner biomass per-recruitment to fishing mortality coefficients for P. hyalocranius (a) and N. taihuensis (b) under two different fishery management schemes in Hongze LakeThe spawner biomass per-recruitment at F=0 is 16.44 g for P. hyalocranius and 12.95 g for N. taihuensis. Fishery management scheme 1 is the current status, with fishing season of one week in Mid-May and between August 9 and December 31; the scheme 2 is an adjusted status, with fishing season between August 29 and December 31. The dotted vertical lines represent the corresponding fishing mortality coefficients under these two fishery management schemes3. 讨论
3.1 大银鱼和太湖新银鱼的阶段性生长差异
鱼类的生长具有阶段性, 生活史的不同阶段, 个体的生理特征、食性以及环境因素等差异, 会引起生长速度的改变[24, 25]。本研究发现, 洪泽湖大银鱼存在2个快速生长阶段, 分别在4—6月和8—11月间。鱼类一般在早期生活史阶段和性成熟前体长生长较快[26], 这一阶段的快速生长可能是鱼类维持种群数量的一种生态适应方式, 有利于降低其被捕食的压力、快速达到性成熟体长[27, 28]。洪泽湖大银鱼繁殖期在12—2月(作者未发表数据), 4—6月间的第一个快速生长阶段, 在个体性成熟之前; 同时, 该时期水体温度逐步上升, 初级生产力升高, 水体中浮游动物等大银鱼的主要饵料生物丰富, 是这一阶段大银鱼快速生长的营养保障[29]。8—11月间第二阶段快速生长可能主要源于大银鱼的食性转化。唐富江发现兴凯湖大银鱼的体长大于60 mm以后, 逐渐由摄食浮游动物转为摄食鱼虾类[30]; 本文作者也发现, 自8月起虾类和鱼类逐渐取代枝角类和桡足类成为洪泽湖大银鱼的主要食物(作者未发表数据)。鱼虾类相对于浮游动物而言具有体积大、能量高、动物蛋白丰富等特点, 更易于被鱼类吸收和利用, 作为生长和繁殖的能量供应[31]。因此, 大银鱼在8—11月的快速生长很可能是因为有更多的能量来源投入到生长中。12月后, 大银鱼种群体长和体重均表现为负增长, 这可能与其进入繁殖期后大个体较早的繁殖和死亡有关[32], 需进一步研究确证。
太湖新银鱼个体相对较小, 其体长在5—10月呈快速持续生长状态, 无明显阶段性。太湖新银鱼终生摄食浮游动物, 与大银鱼不同, 没有明显的食性转化[33]。太湖新银鱼这一食性特点可能是其生长过程没有明显阶段性的主要原因。11月以后, 太湖新银鱼个体体长无明显的增长趋势。洪泽湖太湖新银鱼繁殖季节主要在3—4月, 11月之后, 繁殖季节前, 是性腺发育能量积累的主要阶段[34]。因此, 能量投入到性腺发育中, 可能是太湖新银鱼11月后体长停止增长的重要原因。
3.2 洪泽湖银鱼资源利用评估与管理
鱼类种群可提供渔获量的多少取决于种群数量、生长、存活、开捕年龄和捕捞强度等, 其中仅有捕捞强度(捕捞死亡系数)和初次捕捞年龄(初次开捕体长)能够被人为控制, 因此渔业管理中常通过调整这2个因素以获取最大可持续渔产量[6]。依据平衡产量模型分析, 洪泽湖大银鱼和太湖新银鱼在当前捕捞强度下的渔产量均已接近其资源群体所能提供的最大渔产量, 调整捕捞强度对进一步提高渔产量的影响不大; 而调整2种银鱼的初次开捕年龄, 更有利于获得较高的总渔产量。
在洪泽湖当前的捕捞管理方式下, 大银鱼和太湖新银鱼的SBR残存量分别为未开发状态的20.23%和7.50%, 表明对2种银鱼的捕捞均处于补充型过度捕捞水平, 且太湖新银鱼捕捞后的SBR残余量已远低于种群崩溃的警戒线。由此看来, 较大的捕捞强度和补充型过度捕捞, 可能是导致近年来洪泽湖大银鱼和太湖新银鱼资源波动明显的重要原因。因此, 很有必要对洪泽湖当前银鱼资源捕捞管理方式进行优化, 避免过度捕捞的发生。
本研究构建平衡产量模型时发现, 将洪泽湖银鱼开捕时间推迟20d可获得大银鱼和太湖新银鱼的最大总渔产量。在渔业管理中, 这种开捕时间的推迟也将影响最终捕捞后的产卵亲体生物量[8, 9]。对洪泽湖的银鱼捕捞管理方式进行优化后, 大银鱼的SBR残存量提高至未开发状态的36.72%, 既满足了获得最高渔产量的需求, 又不会带来太大的渔业风险; 优化管理后太湖新银鱼的SBR残存量亦提高至未开发状态的12.86%。可见, 优化的渔业管理方式实现了获取最高渔产量和保护产卵亲体资源上的“双赢”。同时也应注意到, 优化的渔业管理方案对大银鱼产卵群体的保护作用更为显著; 在未改变当前捕捞强度的情况下, 尽管太湖新银鱼在捕捞后的产卵亲体资源量也得到了一定程度的提升, 但依然低于补充型过度捕捞评价阈值20%, 故还需减小对太湖新银鱼的捕捞强度。因此, 针对洪泽湖太湖新银鱼的渔业资源管理, 在延迟开捕时间的基础上, 建议还可以通过限制渔获物的起捕体长、增大捕捞网具的网目规格等方式控制捕捞强度, 同时可采取人工放流等措施提高其下一代的资源补充能力, 以避免种群资源的进一步衰退。
-
![]()
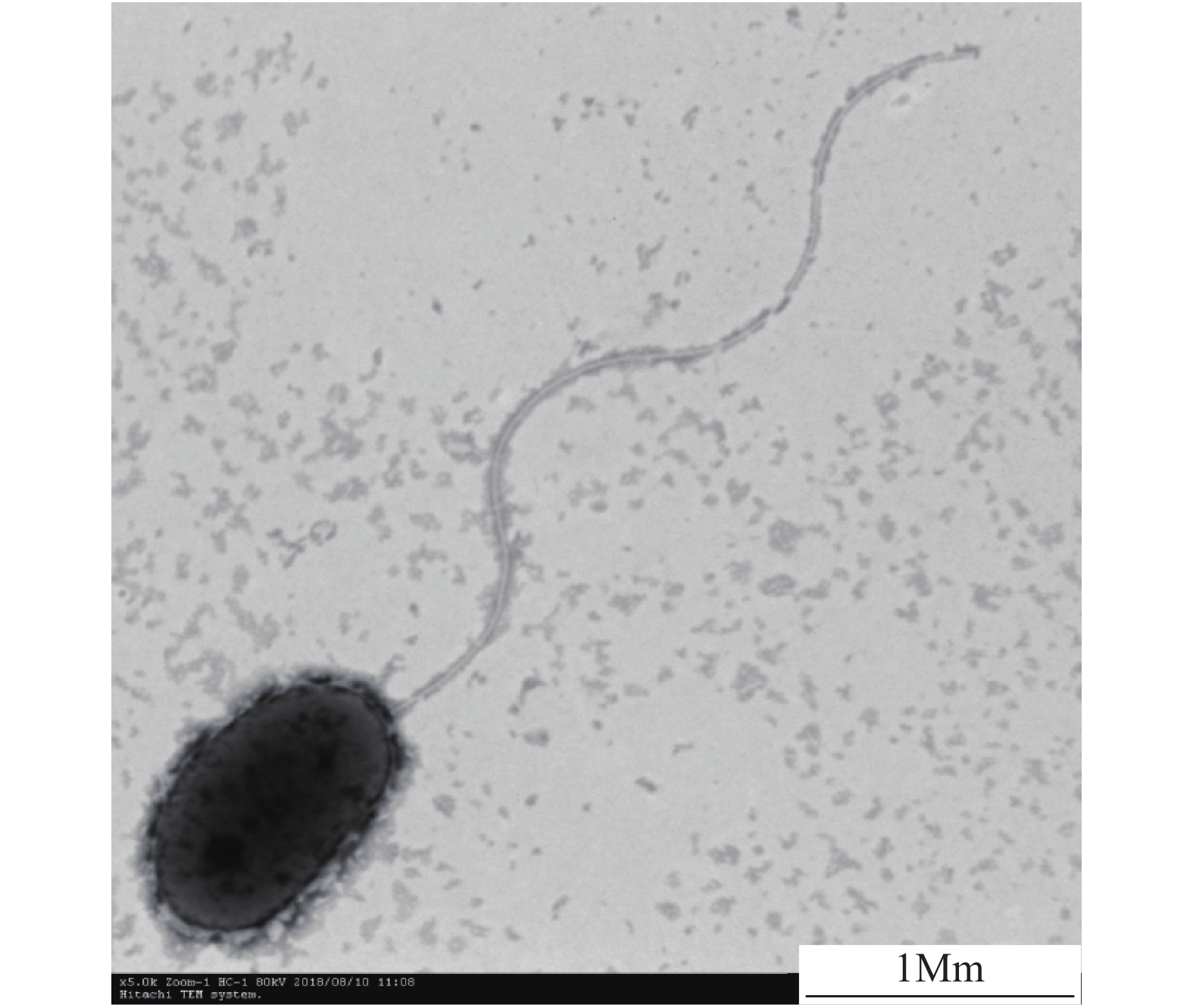
图 1 扫描电镜下菌株ZS1的形态特征
Figure 1. Morphological characteristics of strain ZS1 under a scanning electron microscope
![]()
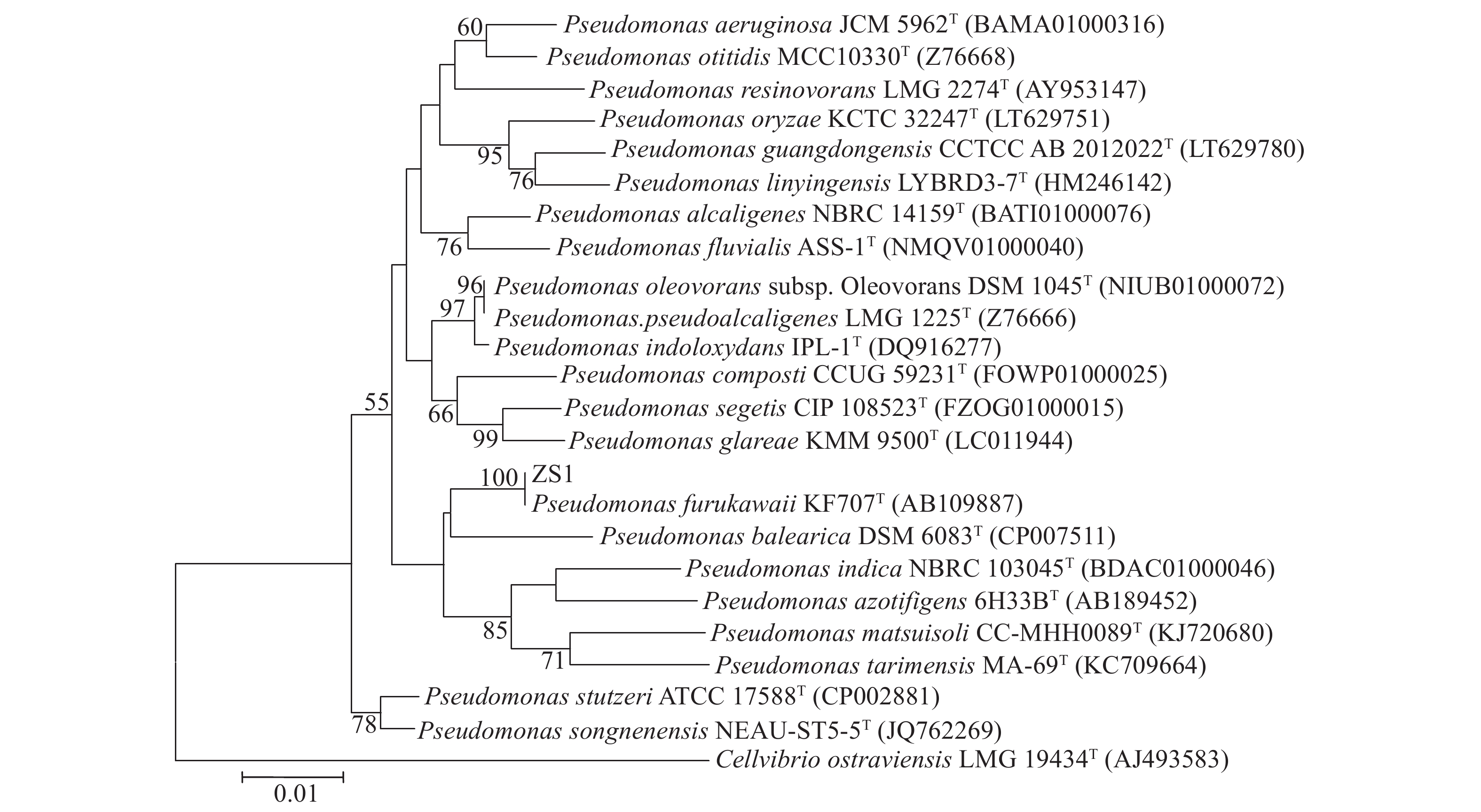
图 2 基于16S rDNA分析的菌株ZS1系统发育树
Figure 2. Phylogenetic tree of strain ZS1 based on 16S rDNA analysis
![]()
图 3 菌株ZS1好氧反硝化特征及生长曲线
Figure 3. Aerobic denitrification performance and growth curve of the strain ZS1
![]()
图 4 碳源对菌株ZS1反硝化性能的影响
Figure 4. Effect of the carbon source on the denitrification ability of strain ZS1
![]()
图 5 C/N对菌株ZS1反硝化性能的影响
Figure 5. Effect of the C/N ratio on the denitrification ability of strain ZS1
![]()
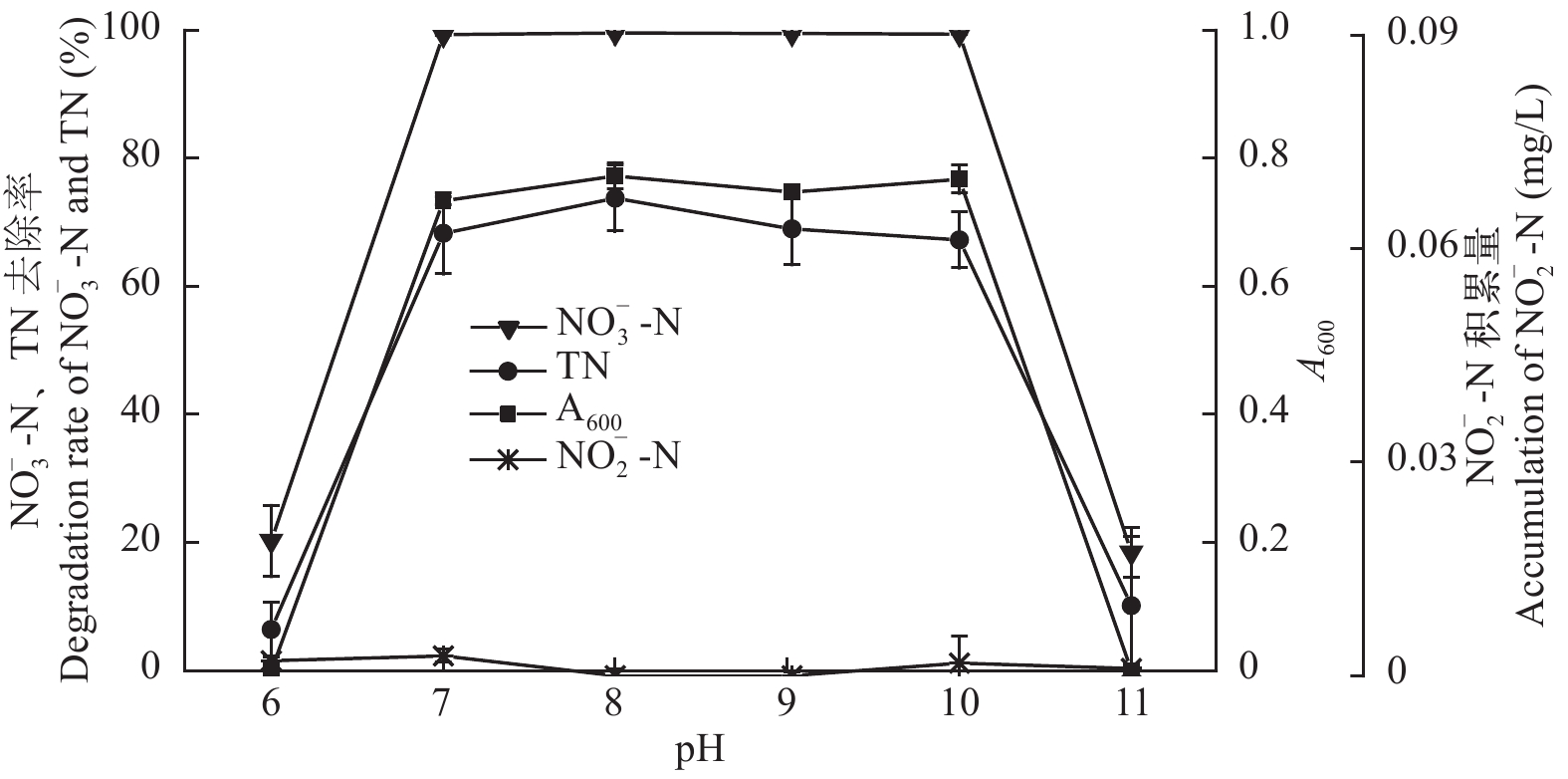
图 6 pH对菌株ZS1反硝化性能的影响
Figure 6. Effect of pH on the denitrification ability of strain ZS1
![]()
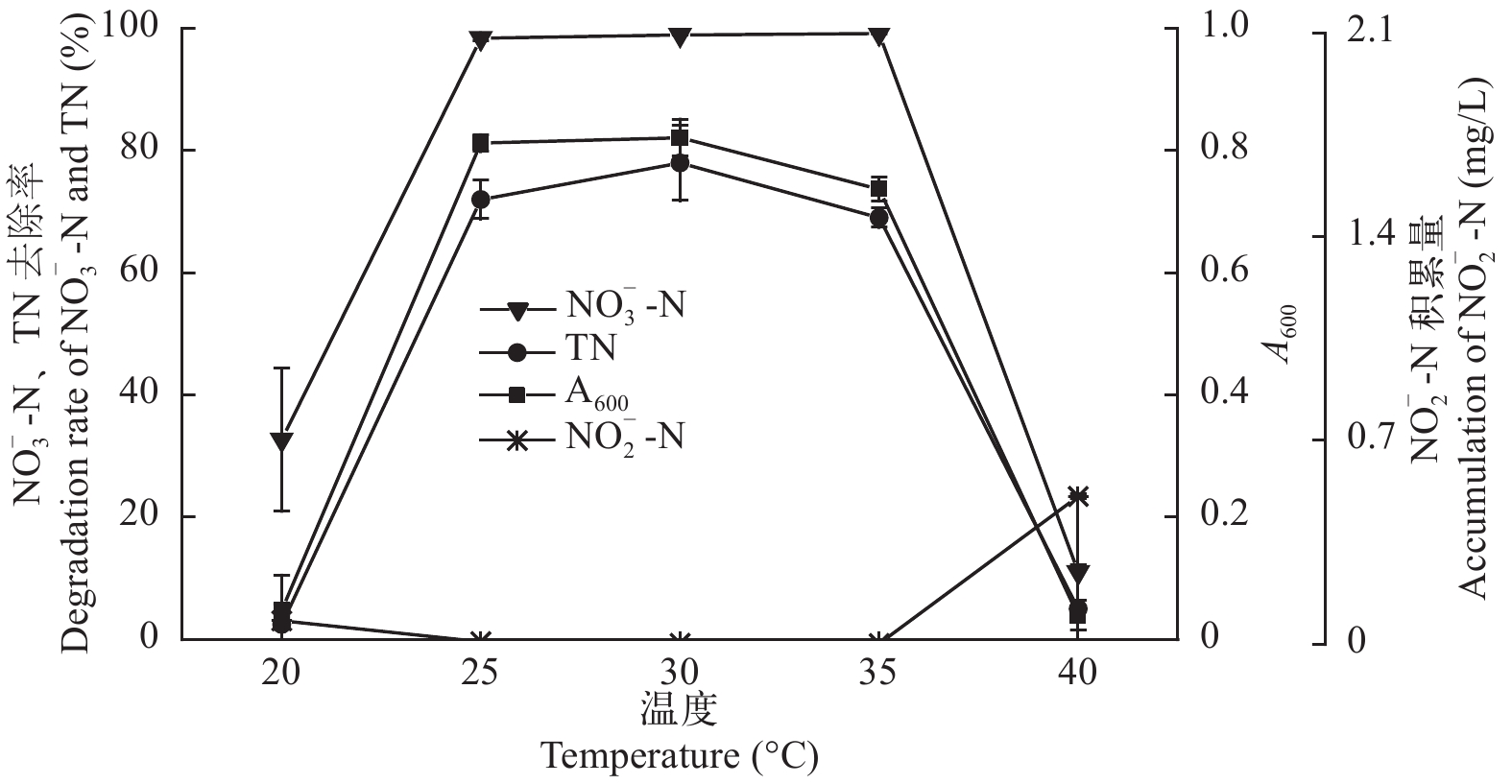
图 7 温度对菌株ZS1反硝化性能的影响
Figure 7. Effect of temperature on the denitrification ability of strain ZS1
![]()
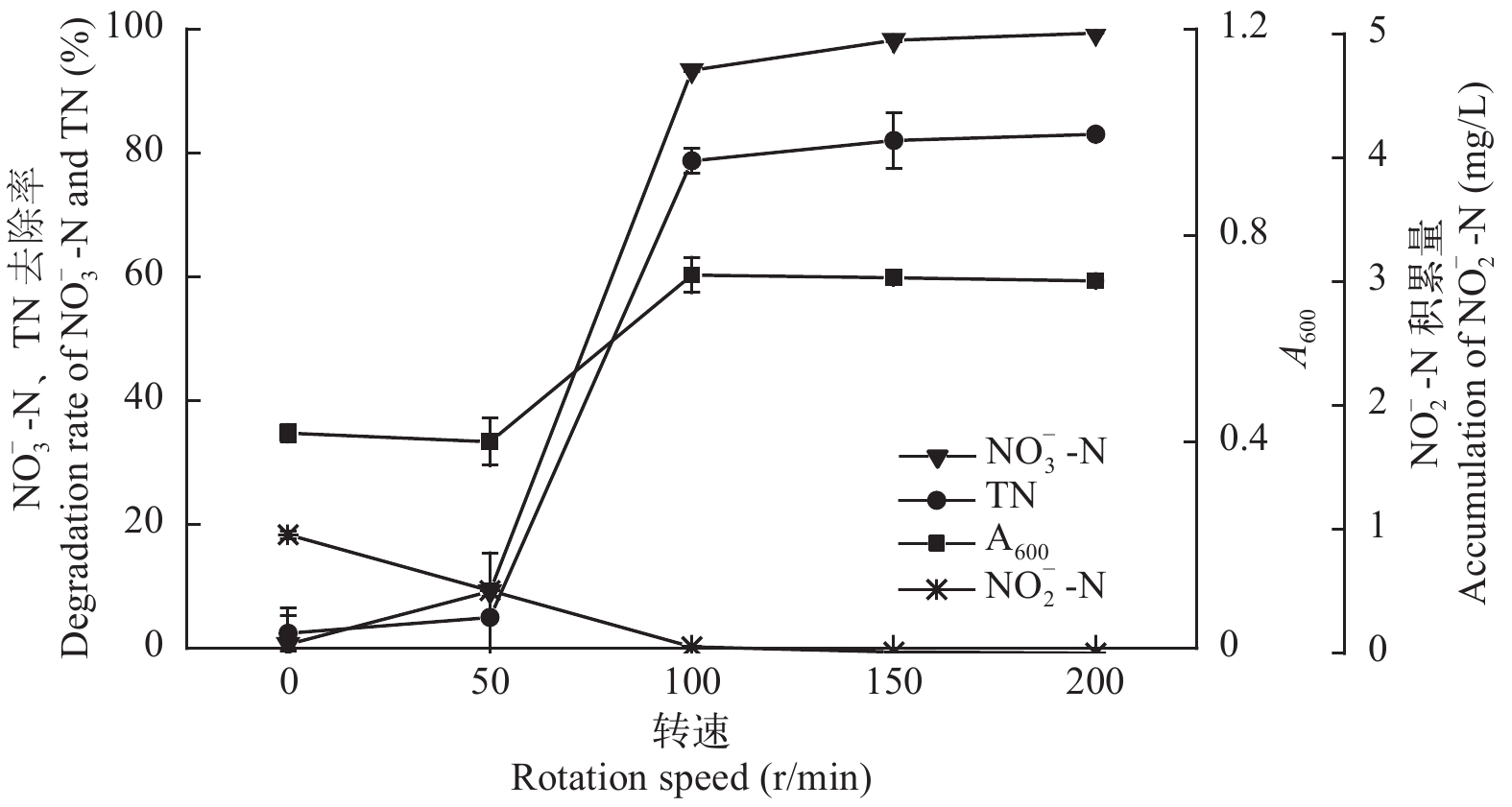
图 8 转速对菌株ZS1反硝化性能的影响
Figure 8. Effect of rotation speed on the denitrification ability of strain ZS1
表 1 菌株ZS1的生理生化特性
Table 1 Physiological and biochemical characteristics of strain ZS1
生理生化试验Physiological and biochemical test 结果Results 生理生化试验Physiological and biochemical test 结果Results 接触酶Contact enzyme + 42℃生长42℃ growth + 氧化酶Oxidase + 45℃生长45℃growth — ONPG测定ONPG determination — 酪素水解Casein hydrolysis — 精氨酸双水解酶Arginine dihydrolase — 脲酶Urease — 硝酸盐还原Nitrate reductase + 厌氧生长Anaerobic growth — 柠檬酸盐利用Citrate utilization + 淀粉水解Starch hydrolysis — 明胶液化Gelatin liquefaction + V-P测定V-P determination — 吲哚Indole — 七叶苷水解Heptapeptide hydrolysis + 注: “+”表示为阳性反应, “—”表示为阴性反应Note: “+” indicates a positive reaction and “—” indicates a negative reaction  下载: 导出CSV
下载: 导出CSV
-
[1] 丁文, 马茵驰, 李文通. 鱼类个体快速识别系统的设计和实现 [J]. 科学养鱼, 2015(1): 75-78. Ding W, Ma Y C, Li W T. Design and implementation of rapid identification system for fish individuals [J]. Scientific Fish Farming, 2015(1): 75-78.
[2] 周娜. 贫营养好氧反硝化菌的筛选、鉴定及脱氮性能研究 [D]. 西安: 西安建筑科技大学, 2014: 7 Zhou N. Isolation and identification of oligotrophic aerobic denitrifying bacteria and denitrification characteristics [D]. Xi’an: Xi’an University of Architecture and Technology, 2014: 7
[3] 杨希. 好氧反硝化细菌应用研究及nosZ基因克隆表达 [D]. 武汉: 华中师范大学, 2008: 7 Yang X. Application of aerobic denitrifying bacteria and cloning expression of nosZ gene [D]. Wuhan: Central China Normal University, 2008: 7
[4] Robertson L A, Kuenen J G. Aerobic denitrification-a controversy revived [J]. Archives of Microbiology, 1984, 139(4): 351-354. doi: 10.1007/BF00408378
[5] Bell L C, Richardson D J, Ferguson S J. Periplasmic and membrane-bound respiratory nitrate reductases in Thiosphaera pantotropha: The periplasmic enzyme catalyzes the first step in aerobic denitrification [J]. FEBS Letters, 1990, 265(1): 85-87.
[6] 孔庆鑫. 一株新型脱氮微生物的分离鉴定及其脱氮机制 [D]. 北京: 中国人民解放军军事医学科学院, 2004: 6 Kong Q X. Screening and identification of a novel denitrogenation microorganism and study on its nitrogen removal mechanism [D]. Beijing: Academy of Military Medical Sciences, 2004: 6
[7] 林敏, 尤崇杓. 粪产碱菌(Alcaligenes faecalis)的反硝化及固氮作用 [J]. 核农学报, 1987, 1(1): 3-10. Lin M, You C Z. Denitrification and nitrogen fixation by Alcaligenes faecalis [J]. Journal of Nuclear Agricultural Sciences, 1987, 1(1): 3-10.
[8] 修海峰, 朱仲元, 丁爱中, 等. 好氧反硝化菌种DF2的分离鉴定及生理生化特性分析 [J]. 生态环境学报, 2011, 20(z2): 1307-1314. Xiu H F, Zhu Z Y, Ding A Z, et al. Isolation and identification of the aerobic denitrifying strain DF2 and its physiological and biochemical analysis [J]. Ecology and Environmental Sciences, 2011, 20(z2): 1307-1314.
[9] 颜薇芝, 张汉强, 余从田, 等. 1株异养硝化好氧反硝化不动杆菌的分离及脱氮性能 [J]. 环境工程学报, 2017, 11(7): 4419-4428. doi: 10.12030/j.cjee.201604200 Yan W Z, Zhang H Q, Yu C T, et al. Isolation of Acinetobacter sp. YN3 and its heterotrophic nitrification-aerobic denitrification characters [J]. Chinese Journal of Environmental Engineering, 2017, 11(7): 4419-4428. doi: 10.12030/j.cjee.201604200
[10] 汪苹, 项慕飞, 翟茜. 从不同反应器筛选、鉴别好氧反硝化菌 [J]. 环境科学研究, 2007, 20(4): 120-124. doi: 10.3321/j.issn:1001-6929.2007.04.021 Wang P, Xiang M F, Zhai Q. Selection and identification of aerobic denitrifiers from different reactors [J]. Research of Environmental Sciences, 2007, 20(4): 120-124. doi: 10.3321/j.issn:1001-6929.2007.04.021
[11] 汤丽娟, 罗固源, 许晓毅, 等. 好氧反硝化菌T7用于废水脱氮特性的研究 [J]. 三峡环境与生态, 2008, 1(3): 24-27. Tang L J, Luo G Y, Xu X Y, et al. Study on nitrogen removal characteristics of an aerobic denitrifying bacterium Rhodococcus strain T7 [J]. Environment and Ecology in the Three Gorges, 2008, 1(3): 24-27.
[12] 李泳洪, 陈小岚, 许旭萍. 异养硝化-好氧反硝化菌Bacillus sp. JB4的分离鉴定试验 [J]. 绿色科技, 2016(16): 23-25. doi: 10.3969/j.issn.1674-9944.2016.16.007 Li Y H, Chen X L, Xu X P. Isolation and identification of heterotrophic nitrification-aerobic denitrifying Bacillus sp. JB4 [J]. Journal of Green Science and Technology, 2016(16): 23-25. doi: 10.3969/j.issn.1674-9944.2016.16.007
[13] 刘燕, 甘莉, 黄哲强, 等. 脱氮副球菌YF1的反硝化特性研究 [J]. 水处理技术, 2010(10): 61-65. Liu Y, Gan L, Huang Z Q, et al. Study on denitrification characteristics of Paracoccus YF1 [J]. Water Treatment Technology, 2010(10): 61-65.
[14] 李慧颖, 黄少斌, 范利荣. 一株好氧反硝化菌的反硝化性能研究 [J]. 环境科学与技术, 2009, 32(8): 9-12. doi: 10.3969/j.issn.1003-6504.2009.08.003 Li H Y, Huang S B, Fan L R. Denitrification characteristics of an aerobic denitrifying bacterium Pseudomonas putida strain A1 [J]. Environmental Science & Technology, 2009, 32(8): 9-12. doi: 10.3969/j.issn.1003-6504.2009.08.003
[15] 蔡亚君, 桂震, 李锋, 等. 好氧反硝化菌Pseudomonas aeruginosa NO62筛选分离与性质鉴定 [J]. 长江大学学报(自科版), 2011, 8(1): 240-243. Cai Y J, Gui Z, Li F, et al. Identification and characterization of an anoxic denitrifying strain Pseudomonas aeruginosa NO62 [J]. Journal of Yangtze University (
Natural Science Edition ) , 2011, 8(1): 240-243. [16] 何腾霞, 徐义, 李振轮. 耐冷亚硝酸盐型反硝化菌Pseudomonas tolaasii Y-11的鉴定及其脱氮特性 [J]. 微生物学报, 2015, 55(8): 991-1000. He T X, Xu Y, Li Z L. Identification and characterization of a hypothermia nitrite bacterium Pseudomonas tolaasii Y-11 [J]. Acta Microbiologica Sinica, 2015, 55(8): 991-1000.
[17] 周国勤, 陈兵, 吴伟. 假单胞菌Pseudomonas stutzeri的生长特性及其反硝化活性 [J]. 广东海洋大学学报, 2004, 24(1): 46-50. doi: 10.3969/j.issn.1673-9159.2004.01.008 Zhou G Q, Chen B, Wu W. Growth characteristic and denitrification activity of Pseudomonas stutzeri [J]. Journal of Guangdong Ocean University, 2004, 24(1): 46-50. doi: 10.3969/j.issn.1673-9159.2004.01.008
[18] 马放. 污染控制微生物学实验 [M]. 哈尔滨: 哈尔滨工业大学出版社, 2002: 42 Ma F. Pollution Control Microbiology Experiment [M]. Harbin: Harbin Institute of Technology Press, 2002: 42
[19] 郭淑清, 田锋. 菌种的甘油冷冻保存法 [J]. 山西医科大学学报, 2000(4): 382-382. Guo S Q, Tian F. Glycerol cryopreservation method for strains [J]. Journal of Shanxi Medical University, 2000(4): 382-382.
[20] 牟东阳, 靳鹏飞, 彭永臻, 等. 1株异养硝化-好氧反硝化细菌DK1的分离鉴定及其脱氮特性 [J]. 环境科学, 2017, 38(11): 4763-4773. Mou D Y, Jin P F, Peng Y Z, et al. Identification and nitrogen removal characteristics of the heterotrophic nitrification and aerobic denitrification bacterial strain DK1 [J]. Environmental Science, 2017, 38(11): 4763-4773.
[21] 孙庆花, 于德爽, 张培玉, 等. 1株海洋异养硝化-好氧反硝化菌的分离鉴定及其脱氮特性 [J]. 环境科学, 2016, 37(2): 647-654. Sun Q H, Yu D S, Zhang P Y, et al. Identification and nitrogen removal characteristics of a heterotrophic nitrification-aerobic denitrification strain isolated from marine environment [J]. Environmental Science, 2016, 37(2): 647-654.
[22] 张婷月, 丁钰, 黄民生. 异养硝化-好氧反硝化细菌的筛选及其脱氮性能研究 [J]. 华东师范大学学报(自然科学版), 2018(6): 22-31. Zhang T Y, Ding Y, Huang M S. Screening of heterotrophic nitrification-aerobic denitrifying bacteria and its nitrogen removal characteristics [J]. Journal of East China Normal University (
Natural Science Edition ) , 2018(6): 22-31. [23] 蔡茜, 何腾霞, 冶青, 等. 耐冷嗜碱蒙氏假单胞菌H97的鉴定及其好氧反硝化特性 [J]. 环境科学, 2018(7): 3314-3320. Cai Q, He T X, Ye Q, et al. Identification and characterization of a hypothermic alkaliphilic aerobic denitrifying bacterium Pseudomonas monteilii strain H97 [J]. Environmental Science, 2018(7): 3314-3320.
[24] 赵惊鸿. 一株耐高温好氧反硝化菌的筛选及特性研究 [J]. 环境科学与技术, 2015, 38(1): 6-10. Zhao J H. Isolation and characteristics of a thermophilic aerobic denitrifier [J]. Environmental Science & Technology, 2015, 38(1): 6-10.
[25] 苑宏英, 孙烨怡, 李原玲, 等. 不同碳源对低温投加氧化还原介体污水生物反硝化脱氮过程的影响 [J]. 化工进展, 2018, 37(2): 783-788. Yuan H Y, Sun Y Y, Li Y L, et al. Effects of different carbon sources on biological denitrification of wastewater at low temperature with adding redox mediator [J]. Chemical Industry and Engineering Progress, 2018, 37(2): 783-788.
[26] 易立, 黄少斌, 潘梦岚. 垃圾渗滤液中好氧反硝化菌的筛选及其反硝化条件优化 [J]. 微生物学杂志, 2017, 37(2): 41-46. doi: 10.3969/j.issn.1005-7021.2017.02.006 Yi L, Huang S B, Pan M L. Screening of aerobic denitrification bacteria in landfill leachate and its denitrification conditions optimization [J]. Journal of Microbiology, 2017, 37(2): 41-46. doi: 10.3969/j.issn.1005-7021.2017.02.006
[27] 田雪雪, 程玉立, 张圆圆, 等. 一株异养硝化-好氧反硝化功能菌的分离鉴定及其脱氮特性 [J]. 环境工程学报, 2017, 11(2): 1269-1275. doi: 10.12030/j.cjee.201509183 Tian X X, Cheng Y L, Zhang Y Y, et al. Isolation, identification and nitrogen removal characteristics of a heterotrophic nitrification-aerobic denitrification bacterium [J]. Chinese Journal of Environmental Engineering, 2017, 11(2): 1269-1275. doi: 10.12030/j.cjee.201509183
[28] 王田野, 魏荷芬, 胡子全, 等. 一株异养硝化好氧反硝化菌的筛选鉴定及其脱氮特性 [J]. 环境科学学报, 2017, 37(3): 945-953. Wang T Y, Wei H F, Hu Z Q, et al. Isolation and identification of a heterotrophic nitrifying and aerobic denitrifying strain and its denitrification characteristics [J]. Acta Scientiae Circumstantiae, 2017, 37(3): 945-953.
[29] 赵丹, 于德爽, 李津, 等. 菌株ZD8的分离鉴定及其异养硝化和缺氧/好氧反硝化特性研究 [J]. 环境科学学报, 2013, 33(11): 3007-3016. Zhao D, Yu D S, Li J, et al. Isolation and identification of a bacterial strain ZD8 and its characteristics of heterotrophic nitrification and anoxic/oxic denitrification [J]. Acta Scientiae Circumstantiae, 2013, 33(11): 3007-3016.
[30] Joong K K, Kyoung J P, Kyoung S C, et al. Aerobic nitrification-denitrification by heterotrophic Bacillus strains [J]. Bioresource Technology, 2005, 96(17): 1897-1906. doi: 10.1016/j.biortech.2005.01.040
[31] Duan J, Fang H, Su B, et al. Characterization of a halophilic heterotrophic nitrification-aerobic denitrification bacterium and its application on treatment of saline wastewater [J]. Bioresource Technology, 2015(179): 421-428.
[32] 王兆阳, 陈国耀, 姜珂, 等. 1株耐冷兼性嗜碱好氧反硝化菌的分离鉴定及反硝化特性 [J]. 环境科学, 2014(6): 2341-2348. Wang Z Y, Chen G Y, Jiang K, et al. Identification and denitrification characteristics of a psychrotolerant facultative basophilic aerobic denitrifier [J]. Environmental Science, 2014(6): 2341-2348.
[33] Gupta A B. Thiosphaera pantotropha: a sulphur bacterium capable of simultaneous heterotrophic nitrification and aerobic denitrification [J]. Enzyme & Microbial Technology, 1997, 21(8): 589-595.
[34] 黄菲菲. 异养硝化-好氧反硝化菌的筛选与脱氮性能研究 [D]. 南京: 南京理工大学, 2013: 32 Huang F F. Identification and nitrogen removal characteristic of two heterotrophic nitrification-aerobic denitrification strains [D]. Nanjing: Nanjing University of Science and Technology, 2013: 32
[35] 张淑楠, 王心义, 夏大平, 等. 地热水中一株好氧反硝化菌的分离鉴定及最优反硝化条件的研究 [J]. 环境污染与防治, 2016, 38(2): 34-39. Zhang S N, Wang X Y, Xia D P, et al. Isolation, identification and denitrification optimization of an aerobic denitrifying bacterium from geothermal water [J]. Environmental Pollution Control, 2016, 38(2): 34-39.
[36] 黄廷林, 张丽娜, 张海涵, 等. 一株贫营养异养硝化-好氧反硝化菌的筛选及脱氮特性 [J]. 生态环境学报, 2015(1): 113-120. Huang T L, Zhang L N, Zhang H H, et al. Screening and nitrogen removal characteristics of a heterotrophic nitrification-aerobic denitrification strain [J]. Ecology and Environmental Sciences, 2015(1): 113-120.
[37] 高喜燕, 刘鹰, 郑海燕, 等. 一株海洋好氧反硝化细菌的鉴定及其好氧反硝化特性 [J]. 微生物学报, 2010, 50(9): 1164-1171. Gao X Y, Liu Y, Zheng H Y, et al. Identification and characteristics of a marine aerobic denitrifying bacterium [J]. Acta Microbiologica Sinica, 2010, 50(9): 1164-1171.
[38] Qin W, Li W G, Zhang D Y, et al. Ammonium removal of drinking water at low temperature by activated carbon filter biologically enhanced with heterotrophic nitrifying bacteria [J]. Environmental Science and Pollution Research, 2016, 23(5): 4650-4659. doi: 10.1007/s11356-015-5561-9
[39] Ye J, Zhao B, An Q, et al. Nitrogen removal by, Providencia rettgeri, strain YL with heterotrophic nitrification and aerobic denitrification [J]. Environmental Technology, 2016, 37(17): 1-8.
[40] 李卫芬, 傅罗琴, 邓斌, 等. 1株好氧反硝化菌的分离鉴定及反硝化特性研究 [J]. 环境科学, 2011, 32(8): 2403-2408. Li W F, Fu L Q, Deng B, et al. Identification and denitrification characteristics of an aerobic denitrifier [J]. Environmental Science, 2011, 32(8): 2403-2408.
-
期刊类型引用(8)
1. 公莉,过龙根,尹成杰,杨姣姣,杨雅兰. 洱海西太公鱼和太湖新银鱼生长特性及种群调控效果研究. 水生态学杂志. 2022(01): 117-123 .  百度学术
百度学术
2. 李大命,唐晟凯,刘燕山,谷先坤,刘小维,殷稼雯,张彤晴,潘建林. 基于Cytb基因的江苏省大银鱼种群遗传多样性和遗传结构分析. 上海海洋大学学报. 2021(03): 416-425 . 百度学术
3. 赵旭昊,徐东坡,任泷,徐跑. 基于Ecopath模型的太湖鲢鳙生态容量评估. 中国水产科学. 2021(06): 785-795 . 百度学术
4. 王煜恒,陈军,李华. 大银鱼仔鱼的人工培育技术探究. 南方农业. 2021(29): 178-179 . 百度学术
5. 李大命,唐晟凯,刘燕山,谷先坤,刘小维,李强,高军,张伟,穆新武,张彤晴. 骆马湖大银鱼、太湖新银鱼线粒体Cytb和COⅠ基因序列多态性分析. 江苏农业科学. 2020(11): 47-52 . 百度学术
6. 李大命,唐晟凯,刘燕山,谷先坤,刘小维,殷稼雯,张彤晴,潘建林. 江苏省4个太湖新银鱼种群遗传多样性和遗传结构分析. 渔业科学进展. 2020(05): 52-60 . 百度学术
7. 唐富江,高文燕,李慧琴,刘伟. 大银鱼生物学与渔业生态学研究进展. 水产学报. 2020(12): 2100-2111 . 百度学术
8. 毛志刚,谷孝鸿,龚志军,曾庆飞,陈辉辉,李红敏,张胜宇,穆欢. 洪泽湖鱼类群落结构及其资源变化. 湖泊科学. 2019(04): 1109-1119 . 百度学术
其他类型引用(7)











计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 15