
RESEARCH ON THE REVISION OF GRATELOUPIA CONSTRICATA LI ET DING BASED ON MORPHOLOGICAL OBSERVATIONS AND EARLY DEVELOPMENT OF SPORES AND MOLECULAR ANALYSES
-
摘要: 通过形态结构和早期发育过程的观察, 结合分子分析, 对采自青岛(模式标本产地)的缢基蜈蚣藻(Grateloupia constricata Li et Ding)进行了重新鉴定。结果表明: (1)藻体直立, 单生或丛生, 呈红褐色或紫红色, 质地黏滑, 软骨质, 高10—30 cm, 主枝扁平; 1—3回羽状分枝, 小枝对生、互生或偏生, 基部缢缩; 藻体由皮层及髓部构成, 皮层由7—10层细胞组成, 厚80—120 μm, 髓部由不规则排列的髓丝组成, 雌配子体的果胞枝生殖枝丛主枝和辅助细胞生殖枝丛主枝分别由6个和5个细胞构成, 为典型的Grateloupia(6cpb—5auxb)型; 四分孢子囊经十字分裂形成。上述特征均与亚洲蜈蚣藻(G. asiatica Kawaguchi et Wang)一致。(2)缢基蜈蚣藻孢子的早期发育类型为“间接盘状体”型, 其早期发育过程与亚洲蜈蚣藻一致。(3)基于rbcL基因序列构建的系统树显示, 8个缢基蜈蚣藻样本之间无碱基差异, 与青岛和大连的亚洲蜈蚣藻均无碱基差异, 形成独立的进化支, 与韩国和日本的亚洲蜈蚣藻碱基差异分别为2 bp(0.17%)和3 bp(0.25%), 均属于种内差异; 基于COⅠ基因序列构建的系统树显示, 8个缢基蜈蚣藻样本之间无碱基差异, 与产于韩国的亚洲蜈蚣藻无碱基差异, 形成独立的进化支; 通过形态、结构及基因序列分析, 确定缢基蜈蚣藻与亚洲蜈蚣藻为同一种, 根据优先法则, 将缢基蜈蚣藻作为亚洲蜈蚣藻的同物异名。Abstract: Through morphological and early development observation followed by molecular analysis, we re-examined Grateloupia constricata Li et Ding collected from Qingdao, Shandong Province. The results showed that: (1) The thalli were upright, brown-red or purple-red, single or clustered, up to 10—30 cm high fleshy and gelatinous in texture. The first branch was flat with pinnately branched 1—3 orders. The branches were reversed, alternate or partial and usually constricted or tapered at the base. Thallus were composed of cortex and medulla. The cortex had 7—10 cell layers and the medulla was consisted of many rhizoidal filaments, 80—120 microns thick, The medulla is consisted of an irregular arrangement of medullary filament. Carpogonial branch ampullae had six cells, auxiliary cell ampullae had five cells (6cpb-5auxb type). Gametophytes dioecious, mature tetrasporangia originated from inner cortex cells and splited into cross. The above characteristics were similar to those of G. asiatica Kawaguchi et Wang. (2) The early developmental type of G. constricata Li et Ding was ‘mediate discal type’, which is consistent with G. asiatica. (3) Based on the phylogenetic tree constructed by rbcL sequences, there was no sequence divergence between the eight samples of G. constricata and G. asiatica from Qingdao and Dalian, which formed a single monophyletic subclade within the large Grateloupia clade of Halymeniaceae. The rbcL sequences differences between G. constricata and G. asiatica from South Korea and Japan were 2 bp (0.17%) and 3 bp (0.25%), which were intraspecific difference. Based on the phylogenetic tree constructed by gene (COⅠ), there was no sequence divergence between the eight samples of G. constricata and G. asiatica from South Korea, which formed a single monophyletic subclade. Consideringmorphological observations and molecular analysis, G. constricata Li et Ding and G. asiatica Kawaguchi et Wang were proved to be the same species. Accroding to the priority rule, G. constricata Li et Ding was the synonym of G. asiatica.
-
随着DNA分子序列分析在藻类分类中的广泛应用, 藻类分类学进入了分子分析的纪元, 分子分析与精细的形态学观察相结合纠正了早期因只根据传统的形态学依据导致的分类错误。2000年Wang等[1]对蜈蚣藻中空变型G. filicina var. lomentaria Howe、蜈蚣藻节荚变型G. filicina var. porracea f. lomentaria (Howe) Okamura和管型藻Sinotibimorpha porracea (Martens ex Kützing) Li et Ding进行了形态观察和rbcL基因序列分析, 结果表明此3种藻类均不是蜈蚣藻G. filicina的变型也不是新种, 而是蜈蚣藻属Grateloupia中的一个独立的种, 即链状蜈蚣藻Grateloupia catenata Yendo, 因此将蜈蚣藻中空变型、蜈蚣藻节荚变型及管型藻作为链状蜈蚣藻的同物异名。Kawaguchi等[2]通过形态学观察和基因序列分析, 发现产自亚洲的蜈蚣藻与产自意大利的模式种蜈蚣藻[G. filicina (Lamouroux) C. Agardh]存在一定程度的序列差异, 因此认为产自亚洲的蜈蚣藻可作为一个新种而独立存在, 并将其命名为: 亚洲蜈蚣藻(G.asiatica Kawaguchi et Wang)。2016年, 李芳等[3]通过形态观察和rbcL基因序列分析, 认为帚状蜈蚣藻G. fastigiata Li et Ding与亚洲蜈蚣藻G. asiatica为同一种, 将帚状蜈蚣藻G. fastigiata作为亚洲蜈蚣藻G. asiatica的同物异名。2017年, 刘芳等[4]运用形态学观察与rbcL基因序列相结合的方法, 对对枝蜈蚣藻G. didymecladia Li et Ding
进行分析, 认为对枝蜈蚣藻与亚栉状蜈蚣藻为同一种, 并将对枝蜈蚣藻作为亚栉状蜈蚣藻的同物异名。由此可见, 在藻类分类学的研究中, 分子分析为区分外部形态相似, 易造成混淆的藻类提供重要的判断依据。其中, 最为常用的分子序列为rbcL基因和COⅠ基因。 本研究的缢基蜈蚣藻记载于《中国海藻志》(夏邦美主编)第二卷第三册中, 是基于简单的形态、结构等特征建立的新种, 该种的主要特征为: 藻体丛生, 紫红色, 主枝圆形、椭圆形或亚扁形, 高5—10 cm, 宽1—1.5 mm。1—2回羽状分枝, 一般分枝基部缢缩或渐细, 小分枝对生、互生或偏生, 呈棍棒状或长锥形。关于缢基蜈蚣藻(G. constricata Li et Ding)的研究仅限于《中国海藻志》中简单的外部形态记载, 而关于它的详细的生殖器官的结构、发育及分子分析等方面的研究均是空白, 因此本研究对缢基蜈蚣藻G. constricata的形态、结构、孢子早期发育和DNA分子分析进行详细的研究, 从而判断缢基蜈蚣藻G. constricata作为新种是否成立, 明确缢基蜈蚣藻G. constricata在蜈蚣藻属(Grateloupia C. Agardh)中的分类地位。本研究将为我国蜈蚣藻属的分类修订、物种多样性及《中国海藻志》的再版提供新信息、新资料。
1. 材料与方法
1.1 材料的采集与处理
标本的采集: 缢基蜈蚣藻样本于2017年6月5日, 由王惠娜和丁媛媛采自模式标本产地山东省青岛市的第一海水浴场、鲁迅公园和麦岛。
标本的处理: 将采集的海藻去除杂质和附生藻类。少数雌配子体和四分孢子体置于解剖镜下仔细观察, 反复清理至藻体干净后, 置于灭菌的海水中备用; 其余的海藻进行编号(表 1), 制成腊叶标本, 用于观察外部形态结构; 新鲜标本制成冰冻切片, 用于观察内部结构; 硅胶干燥标本, 提取DNA, 用于后续的分子分析。
表 1 缢基蜈蚣藻的标本号、采集地点和基因登录号Table 1. Specimen No., collection information and GenBank accession number标本编号Specimen No. 采集地点Collection information 基因登录号GenBank accession number rbcL COⅠ LNU2017060501 The First Swimming Beach, Qingdao, China MN412709 MN412717 LNU2017060502 The First Swimming Beach, Qingdao, China MN412710 MN412718 LNU2017060503 The First Swimming Beach, Qingdao, China MN412711 MN412719 LNU2017060504 Luxun Park, Qingdao, China MN412712 MN412720 LNU2017060505 Luxun Park, Qingdao, China MN412713 MN412721 LNU2017060506 Luxun Park, Qingdao, China MN412714 MN412722 LNU2017060507 Maidao, Qingdao, China MN412715 MN412723 LNU2017060508 Maidao, Qingdao, China MN412716 MN412724 1.2 实验方法
外部形态的观察: 对标本的外部形态特征进行详细的观察和记录, 并用Canon EOS 650D拍摄照片。选取合适的腊叶标本制成临时装片, 置于光学显微镜和解剖镜下观察生殖结构。
内部结构的观察: 选取具备生殖器官(囊果或四分孢子囊)的新鲜标本, 一般切割为1 cm左右的小段置于冰冻切片机(YD-335Ⅲ)的刀台上, 用OCT包埋剂(SAKURA Tissue-Tek O.C.T. Compound 4583)使其完全被包埋, 制作成冰冻切片, 观察内部结构。
培养方法: 首先, 用75%的酒精浸泡脱脂棉, 清洁玻璃培养容器并用脱脂棉擦拭, 晾干后用灭菌的海水冲洗3次。而后, 在玻璃培养容器底部铺一层载玻片(7个), 将之前备用的雌配子体或四分孢子体及灭菌的空气泵放入容器中, 加入灭菌的海水, 至海水没过藻体。空气泵通电, 待孢子释放。对光照培养箱(LRH-250-GB)的培养条件进行设置: 温度20℃, 光照强度80 μmol/(m2·s), 光周期(Light﹕ Dark)12L﹕12D。24h后, 对载玻片进行观察, 孢子的释放量为15—30时(10×10倍视野), 将这些载玻片置于配置好的培养液中, 放入光照培养箱(LRH-250-GB)。每天观察孢子萌发情况, 我们采用了Olympus BX53荧光显微镜观察萌发管中原生质体的流动方向, 并拍照记录, 每2天更换1次培养液。使用的培养液为20 mL PES培养液, 2 mg GeO2, 980 mL灭菌海水。
基因序列分析: 使用植物基因组DNA提取试剂盒(TIAGEN, Valencia, CA, Beijing)提取藻体样本的DNA, 并进行rbcL和COⅠ基因扩增, 具体实验流程参考王宏伟等[5]改进的方法, PCR扩增产物经1%琼脂糖凝胶电泳检测, 检测后由上海生工生物公司代为纯化和测序(表 1)。引物设计: rbcL基因引物组合为 F29-R1150, F765-R1381, 共4个引物, 2对组合; COⅠ基因引物组合为 COIF1-COIR1, 2个引物, 1对组合。系统树构建: 从GenBank网站下载并选取蜈蚣藻属40个种[6—11]及2个外群种的rbcL基因序列, 15个种[12, 13]及2个外群种的COⅠ基因序列。(外群选取的原则是选取与内群具最近亲缘关系的物种[5])将两组基因序列分别与本研究中得到的8个缢基蜈蚣藻的rbcL和COⅠ基因序列进行对比。序列比对和校正软件采用Clustalx(1.83)[14], 碱基差异度的分析和系统发育树的构建使用软件PAUP 4.0[15]和MEGA 5.0[16]; 算法为Maximum likelihood (ML)法, 计算遗传距离的模型为Number of differences和Kimura 2-Parameter, Bootstrap值为1000。
2. 结果
2.1 缢基蜈蚣藻的外部形态
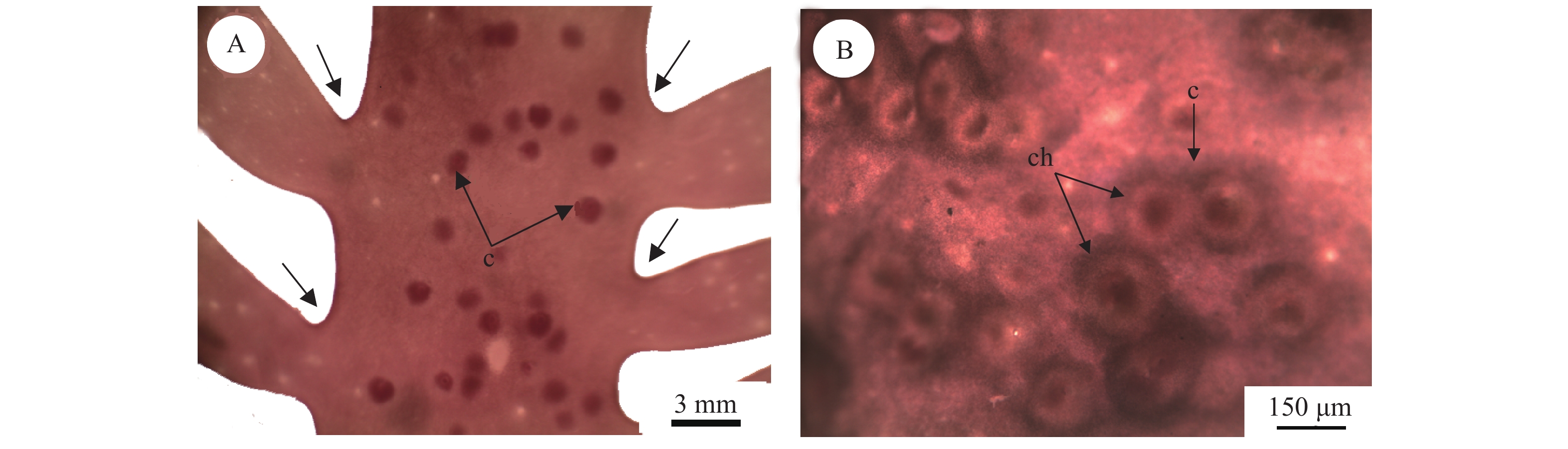
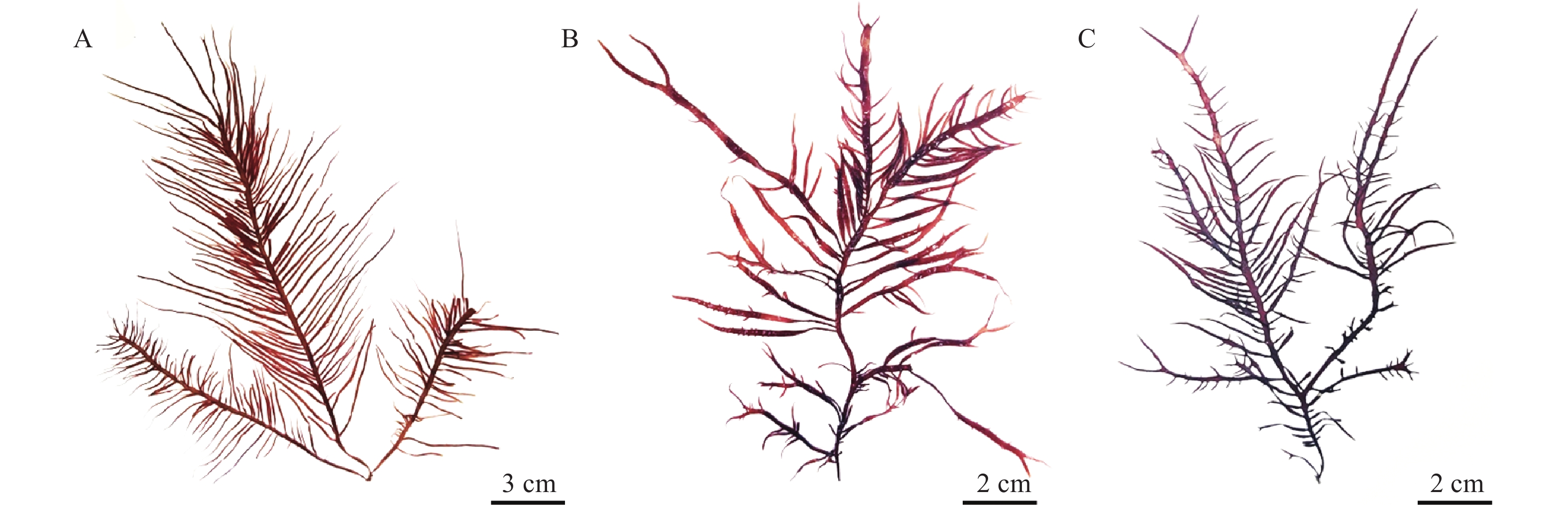
图 1为缢基蜈蚣藻的外部形态, 颜色呈红褐色或紫红色, 丛生, 直立, 高10—30 cm, 质地黏滑, 软骨质, 主枝扁平; 1—3回羽状分枝, 对生、互生或偏生, 小羽枝生长在主枝上或主枝边缘, 基部缢缩(图 2A)。囊果近球形, 分布于藻体的主枝和小枝, 稍突出藻体表面(图 2B)。
![]() 图 1 缢基蜈蚣藻的外部形态A. 四分孢子体(LNU17060512); B. 雌配子体(LNU17060506); C. 雄配子体(LNU17060565)Figure 1. External morphology of G. constricata Li et DingA. Tetrasporangia (LNU17060512); B. Female gametophyte (LNU17060506); C. Male gametophyte (LNU17060565)
图 1 缢基蜈蚣藻的外部形态A. 四分孢子体(LNU17060512); B. 雌配子体(LNU17060506); C. 雄配子体(LNU17060565)Figure 1. External morphology of G. constricata Li et DingA. Tetrasporangia (LNU17060512); B. Female gametophyte (LNU17060506); C. Male gametophyte (LNU17060565)![]() 图 2 缢基蜈蚣藻囊果A. 缢基蜈蚣藻羽枝基部存在缢缩(小箭头所示), 囊果在主枝和小枝上均有分布(c, 囊果); B. 主枝上的囊果及囊果孔展示(c, 囊果; ch, 囊果孔)Figure 2. Branches of G. constricata Li et DingA. Branches of G. constricata Li et Ding with constricted at bases (small arrowheads) and cystocarps distributed on axes and lateral branches of G. constricata Li et Ding (c, cystocarp); B. Cystocarp distribution on axes(c, cystocarp; ch, cystocarp hole)
图 2 缢基蜈蚣藻囊果A. 缢基蜈蚣藻羽枝基部存在缢缩(小箭头所示), 囊果在主枝和小枝上均有分布(c, 囊果); B. 主枝上的囊果及囊果孔展示(c, 囊果; ch, 囊果孔)Figure 2. Branches of G. constricata Li et DingA. Branches of G. constricata Li et Ding with constricted at bases (small arrowheads) and cystocarps distributed on axes and lateral branches of G. constricata Li et Ding (c, cystocarp); B. Cystocarp distribution on axes(c, cystocarp; ch, cystocarp hole)2.2 缢基蜈蚣藻的营养结构
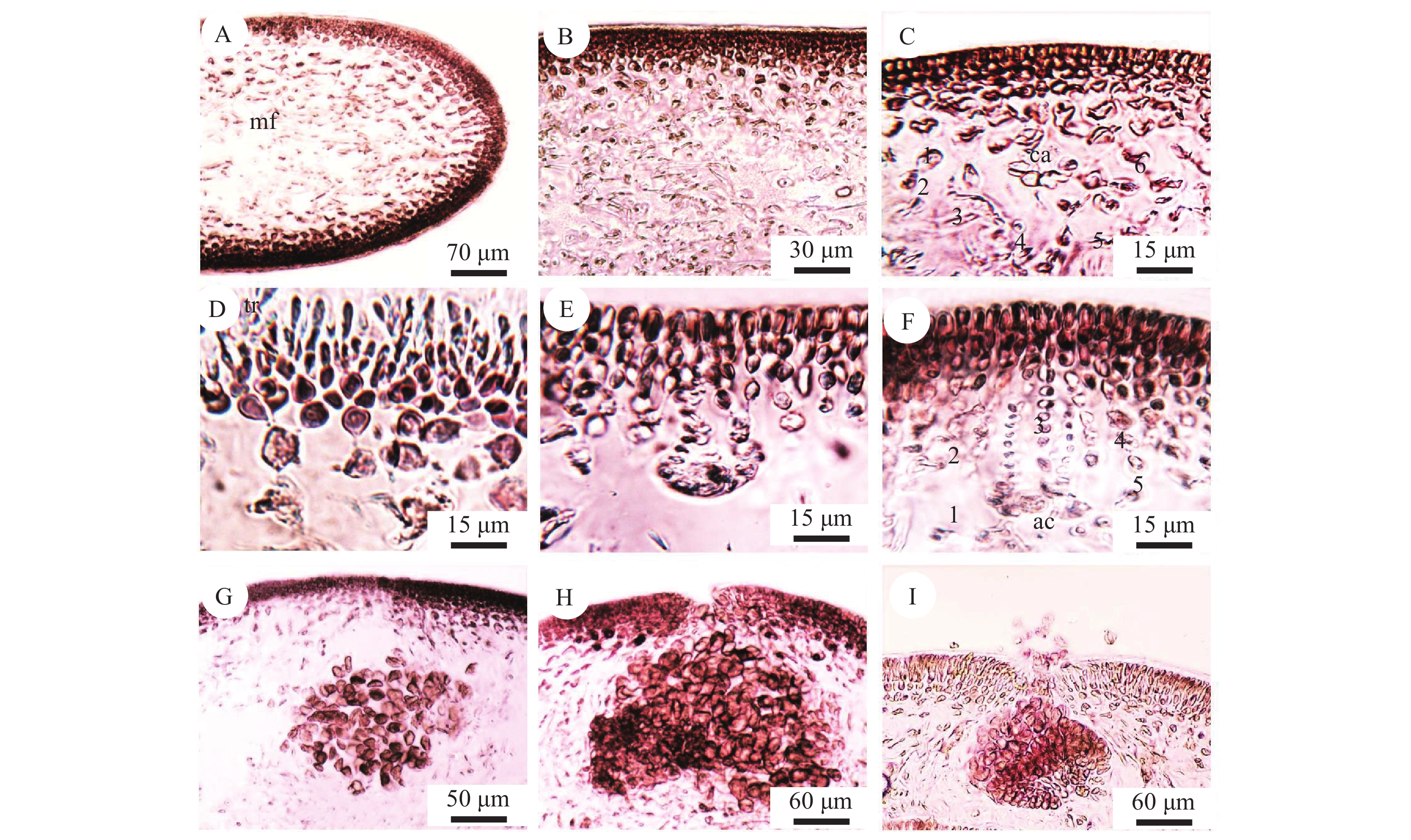
缢基蜈蚣藻横切面观, 厚300—400 μm。如图 3A所示, 皮层位于横切面两端, 厚80—120 μm, 共由7—10层细胞组成, 包括外皮层和内皮层。外皮层细胞略小, 通常3—5层细胞; 与外皮层相连的内皮层, 细胞较大, 呈不规则的星形或长椭圆形, 通常4—5层细胞。中央丝状区域为髓部, 由细长的髓丝细胞组成, 排列不规则(图 3A、3B)。
![]() 图 3 缢基蜈蚣藻内部结构A、B. 藻体的横切面观(mf, 髓丝); C. 雌配子体的果胞枝生殖枝丛横切面观(1—6. 果胞枝生殖枝丛主枝; ca. 果胞); D. 雄配子体的精子囊; E、F. 雌配子体的辅助细胞生殖枝丛横切面观(1—5, 辅助细胞生殖枝丛主枝; ac, 辅助细胞); G、H. 囊果连续的发育过程; G. 幼小囊果; H. 成熟囊果; I. 果孢子放散; 切片标本号: LNU17060506、LNU17060516、LNU17060536、LNU17060565Figure 3. The internal structure of G. constricata Li et DingA, B. Cross-section of the thallus (mf, medullary filament); C. Transection of carpogonial branch ampullae from female gametophyte (1—6, leader branch carpogonial branch ampullae; ca, carpogonium); D. Spermatangium of male gametophyte; E, F. Transection of auxiliary cell ampullae from female gametophyte (1—5, leader branch of auxiliary cell ampullae; ac, auxiliary cell); G, H. Cystocarp development; G. Little cystocarp; H. Developing cystocarp; I. Release of mature carpospores. Section number: LNU17060506, LNU17060516, LNU17060536, LNU17060565
图 3 缢基蜈蚣藻内部结构A、B. 藻体的横切面观(mf, 髓丝); C. 雌配子体的果胞枝生殖枝丛横切面观(1—6. 果胞枝生殖枝丛主枝; ca. 果胞); D. 雄配子体的精子囊; E、F. 雌配子体的辅助细胞生殖枝丛横切面观(1—5, 辅助细胞生殖枝丛主枝; ac, 辅助细胞); G、H. 囊果连续的发育过程; G. 幼小囊果; H. 成熟囊果; I. 果孢子放散; 切片标本号: LNU17060506、LNU17060516、LNU17060536、LNU17060565Figure 3. The internal structure of G. constricata Li et DingA, B. Cross-section of the thallus (mf, medullary filament); C. Transection of carpogonial branch ampullae from female gametophyte (1—6, leader branch carpogonial branch ampullae; ca, carpogonium); D. Spermatangium of male gametophyte; E, F. Transection of auxiliary cell ampullae from female gametophyte (1—5, leader branch of auxiliary cell ampullae; ac, auxiliary cell); G, H. Cystocarp development; G. Little cystocarp; H. Developing cystocarp; I. Release of mature carpospores. Section number: LNU17060506, LNU17060516, LNU17060536, LNU170605652.3 缢基蜈蚣藻的生殖结构
雌配子体的生殖结构由其内皮层细胞形成。果胞枝生殖枝丛与辅助细胞生殖枝丛符合典型的Grateloupia (6cpb-5auxb)型。果胞枝生殖枝丛生长在较大的支持细胞上, 与藻体表面相邻处连接一条受精丝, 受精丝位于藻体外侧, 呈伸直或弯曲状, 该受精丝与果胞枝生殖枝丛中的壶形果胞相连。果胞周围有6个细胞, 包括下位细胞、亚下位细胞、基细胞及3个圆形细胞。6个细胞均带有一条侧枝, 形成果胞枝生殖枝丛, 即瓶状体(图 3C)。辅助细胞生殖枝丛的基底细胞为辅助细胞, 成熟的辅助细胞呈圆球形(图 3E—F)。在其周围可观察到5个细胞, 每个细胞均带有一条侧枝, 当侧枝完全发育并且扩大, 直到辅助细胞易于识别时, 每条侧枝的长度可达10—12个细胞, 这5条侧枝将辅助细胞包围在中心, 形成辅助细胞生殖枝丛, 即瓶状体。
缢基蜈蚣藻雄配子体精子囊(图 3D)中的精母细胞由表皮细胞分化而来, 表皮细胞首先纵向分裂然后伸长, 随后它们横向分裂形成末端精子。精子经雌配子体表面的受精丝进入果胞中, 与果胞结合, 受精后, 二倍体细胞核可能通过联络丝从果胞枝生殖枝丛进入到瓶状体的辅助细胞中, 辅助细胞和其两个相邻的瓶状体细胞之间发生细胞融合, 形成融合复合体。该复合体上产生三级联络丝和一个初级产孢丝指向藻体表面, 在产孢丝发育的晚期阶段, 发育中的产孢丝附近的髓丝最终通过次生纹孔连接交织并互连, 形成致密的囊果皮。完全发育成熟的产孢丝由一至两个产孢瓣组成, 并且大多数产孢丝转化为球形的囊果, 成熟囊果直径范围为190—230 μm, 果孢子囊上有囊果孔, 果孢子经囊果孔逸出(图 3H)。
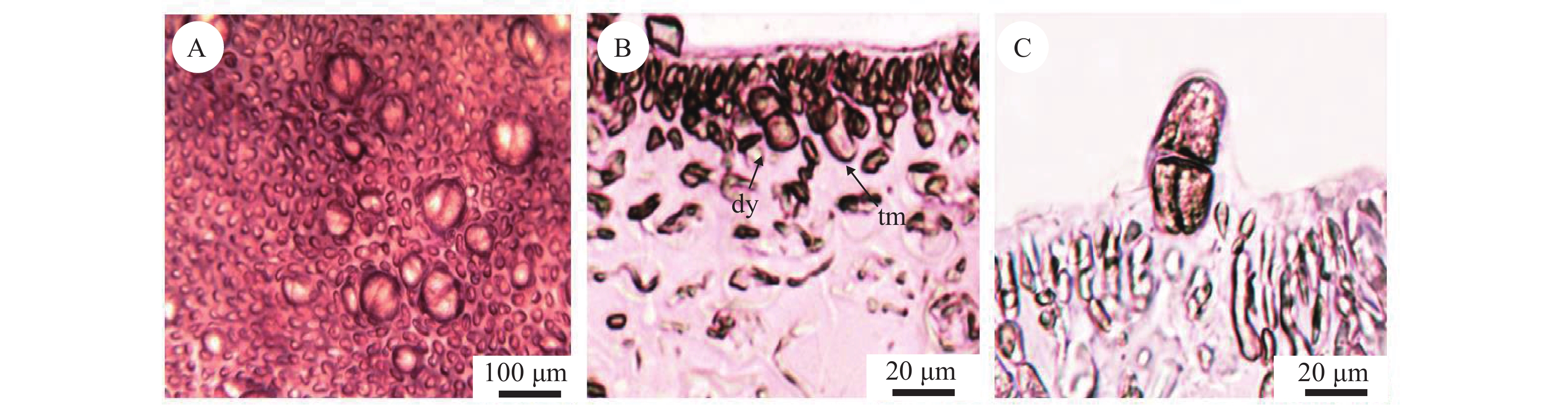
四分孢子体表面较大的突起为四分孢子囊, 由孢子体内皮层细胞产生(图 4A)。四分孢子囊在主枝与小枝上均有分布。四分孢子囊母细胞 (图 4B)经减数分裂形成成熟的四分孢子囊(图 4C), 为长椭圆形(长36—52 μm×宽14—20 μm)。
![]() 图 4 缢基蜈蚣藻四分孢子体内部结构A. 四分孢子囊表面观; B. 四分孢子囊母细胞及二分体 (tm, 四分孢子囊母细胞; dy, 二分体); C. 成熟的四分孢子囊。切片标本号: LNU17060512Figure 4. The internal structure of tetrasporophyte of G. constricata Li et DingA. Tetrasporophyte surface view; B. Tetrasporoblas and formed dyad (tm, tetrasporoblas; dy, formed dyad); C. Etrasporangia cross division, Section number: LNU17060512
图 4 缢基蜈蚣藻四分孢子体内部结构A. 四分孢子囊表面观; B. 四分孢子囊母细胞及二分体 (tm, 四分孢子囊母细胞; dy, 二分体); C. 成熟的四分孢子囊。切片标本号: LNU17060512Figure 4. The internal structure of tetrasporophyte of G. constricata Li et DingA. Tetrasporophyte surface view; B. Tetrasporoblas and formed dyad (tm, tetrasporoblas; dy, formed dyad); C. Etrasporangia cross division, Section number: LNU170605122.4 缢基蜈蚣藻孢子的早期发育
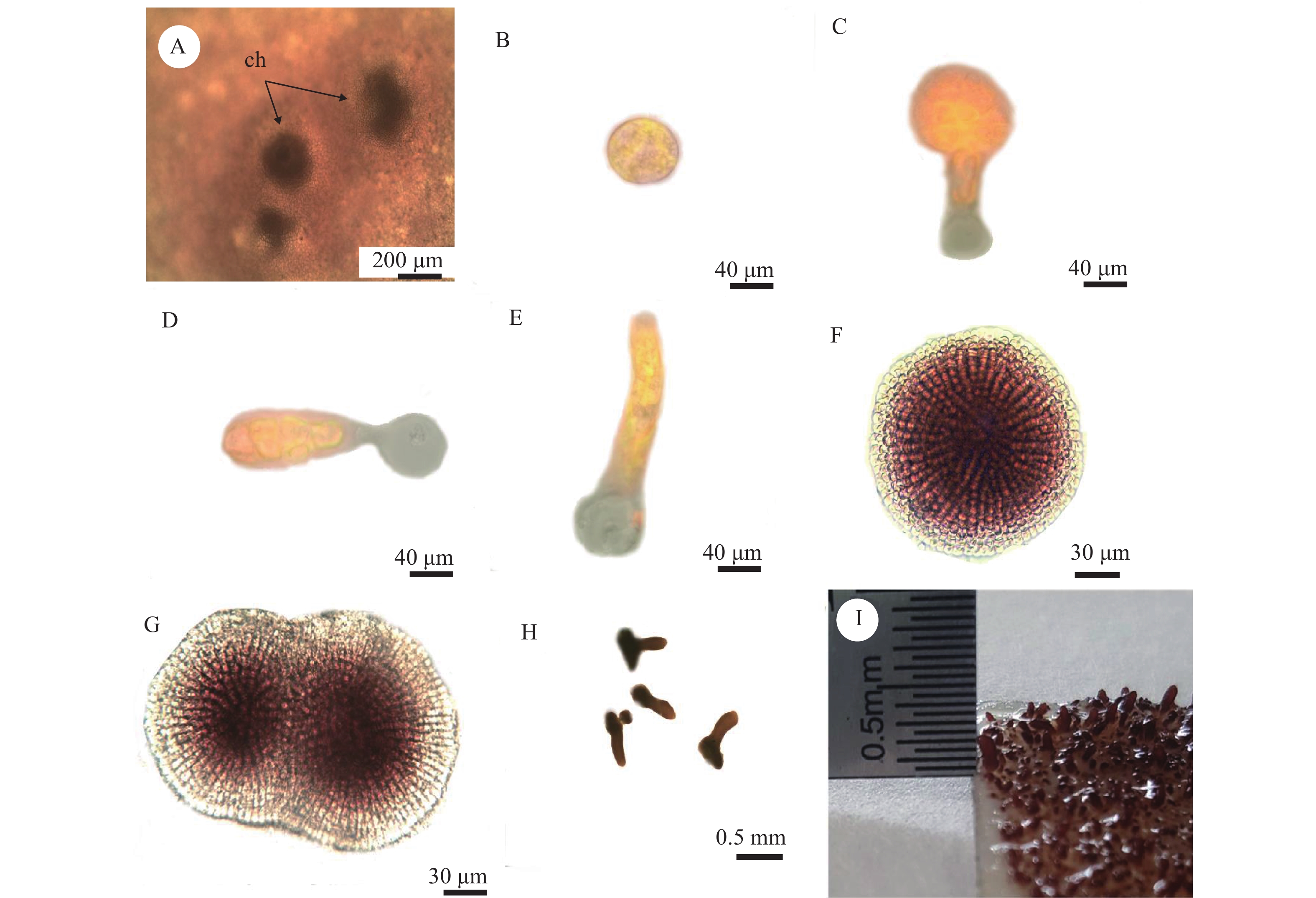
雌配子体成熟囊果中的果孢子经囊果孔逸出, 附着在载玻片上(图 5A和5B)。果孢子为球形或椭圆形, 呈浅红色, 直径约15 μm。将果孢子培养24h后, 开始萌发。通过荧光显微镜可以观察到, 果孢子的一端凸起, 形成萌发管, 萌发管伸长, 果孢子中的原生质体全部进入萌发管中(图 5C), 并在萌发管与原处的半透明胶状物质连接处形成一道隔膜。萌发管内细胞开始不断分裂(图 5D—F), 随后, 形成盘状体, 最大的盘状体直径可达310 μm左右(图 5F)。盘状体与临近的多个盘状体相互融合(图 5G), 其临近中心部位分化产生1—5个突起, 形成直立芽体(图 5H)。再经30d左右, 直立芽体分化成直立枝(图 5I)。继续培养, 直立枝发育成幼苗。
![]() 图 5 缢基蜈蚣藻果孢子的萌发过程A. 藻体表面的囊果(ch, 囊果孔); B—E为荧光显微镜拍摄结果; B. 果孢子; C. 果孢子内原生质体迁移; D—E. 萌发管内细胞分裂过程; F. 盘状体时期; G. 盘状体融合的过程; H. 直立芽体形成; I. 直立枝Figure 5. The development of carpospores in G. constricataA. Carposporophytes on the surface of thalli (ch. cystocarp hole); B—E were taken with a fluorescence microscope; B. Carpospores; C. Transformation of protoplasts in the germ tube; D—E. Cell division in the germ tube; F. Discoid crust; G. Fusion of discoid crust; H. Upright branches; I. Growth of sporeling
图 5 缢基蜈蚣藻果孢子的萌发过程A. 藻体表面的囊果(ch, 囊果孔); B—E为荧光显微镜拍摄结果; B. 果孢子; C. 果孢子内原生质体迁移; D—E. 萌发管内细胞分裂过程; F. 盘状体时期; G. 盘状体融合的过程; H. 直立芽体形成; I. 直立枝Figure 5. The development of carpospores in G. constricataA. Carposporophytes on the surface of thalli (ch. cystocarp hole); B—E were taken with a fluorescence microscope; B. Carpospores; C. Transformation of protoplasts in the germ tube; D—E. Cell division in the germ tube; F. Discoid crust; G. Fusion of discoid crust; H. Upright branches; I. Growth of sporeling四分孢子的早期发育过程与果孢子的完全一致。
2.5 缢基蜈蚣藻的rbcL和COⅠ基因序列分析结果
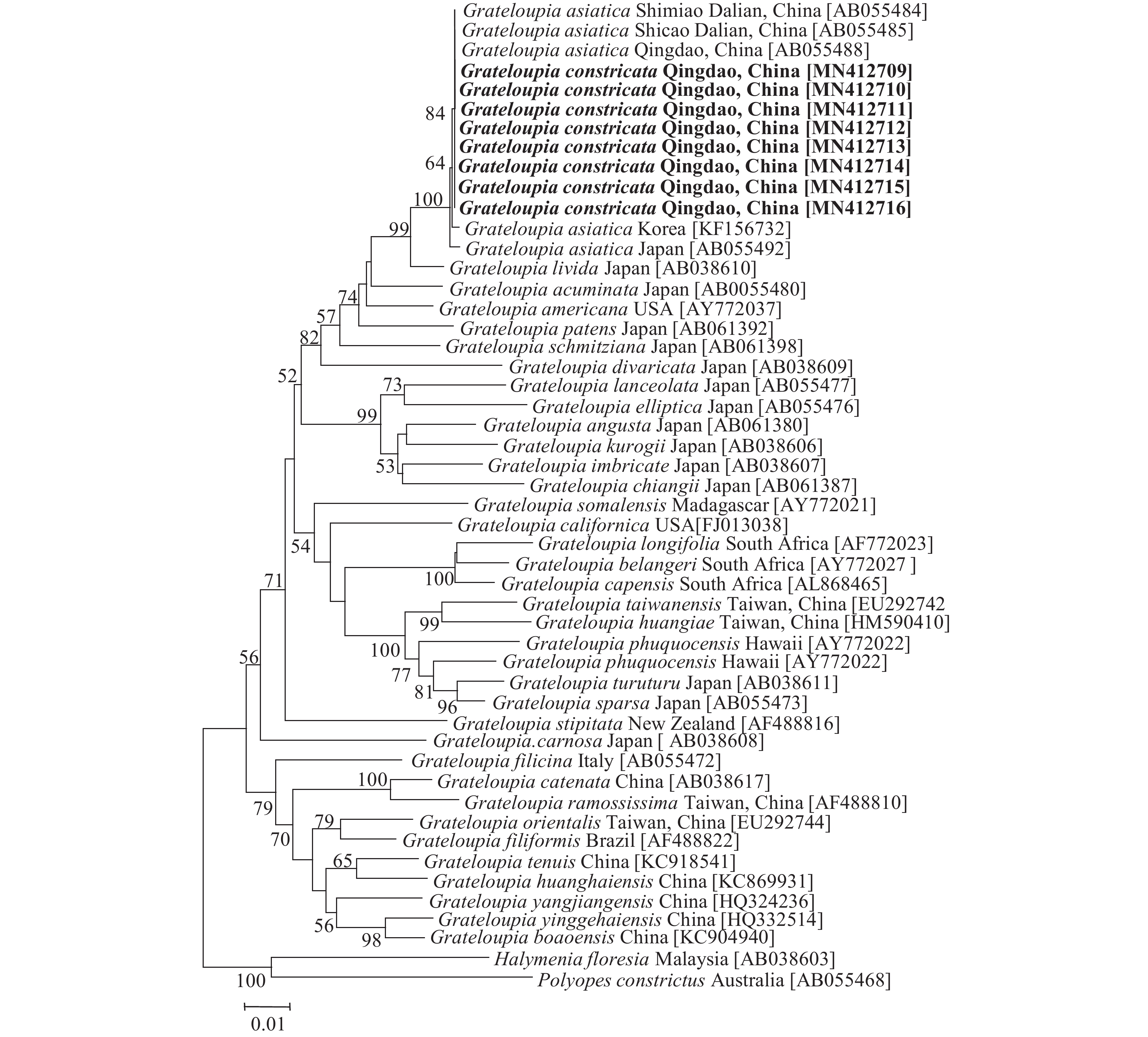
rbcL基因序列分析 缢基蜈蚣藻的8个rbcL基因序列长度均为1244 bp, 比对校正后为1198 bp。序列比对(MAGE5)结果显示如图 6, 8个缢基蜈蚣藻的基因序列间无碱基差异, 与青岛、大连的亚洲蜈蚣藻均无碱基差异, 与韩国的亚洲蜈蚣藻碱基差异为2 bp (0.17%), 与日本的亚洲蜈蚣藻碱基差异为3 bp (0.25%), 它们位于独立的发育枝中。缢基蜈蚣藻与舌状蜈蚣藻(G. livida)碱基差异为17 bp (1.42%), 与顶状蜈蚣藻(G. acuminate)碱基差异为34 bp (2.84%), 与美国蜈蚣藻(G. americana)碱基差异为34 bp (2.84%), 与伸展蜈蚣藻(G. patens)碱基差异为42 bp (3.51%), 与史氏蜈蚣藻(G. schmitziana)碱基差异为47 bp (3.92%), 与叉枝蜈蚣藻(G. divaricata)碱基差异为70 bp (5.84%)。与蜈蚣藻(G. filicina)的碱基差异为87 bp (7.26%), 均属于种间差异。与作为外群的海膜属(Halymenia C. Agardh)的H. floresia和海柏属(Polyopes J. Agardh)的P. constrictus的碱基差异分别为119 bp (9.93%)、124 bp (10.35%)。
![]() 图 6 缢基蜈蚣藻基于rbcL基因的ML系统树分支上的数字展示基于1000次重复的支持率, 只显示支持率大于50%的数值, Halymenia floresia Malaysia和Polyopes constrictus Australia作为外群, 粗体字为本文研究种; 下同Figure 6. Maximum likelihood tree based on rbcL gene of G. constricataNumerals at internal nodes are bootstrap values (1000 replicates), only values above 50% bootstrap support are shown, Halymenia floresia Malaysia and Polyopes constrictus Australia were used as outgroups. Boldfaced words are the species studied in this study. The same applies below
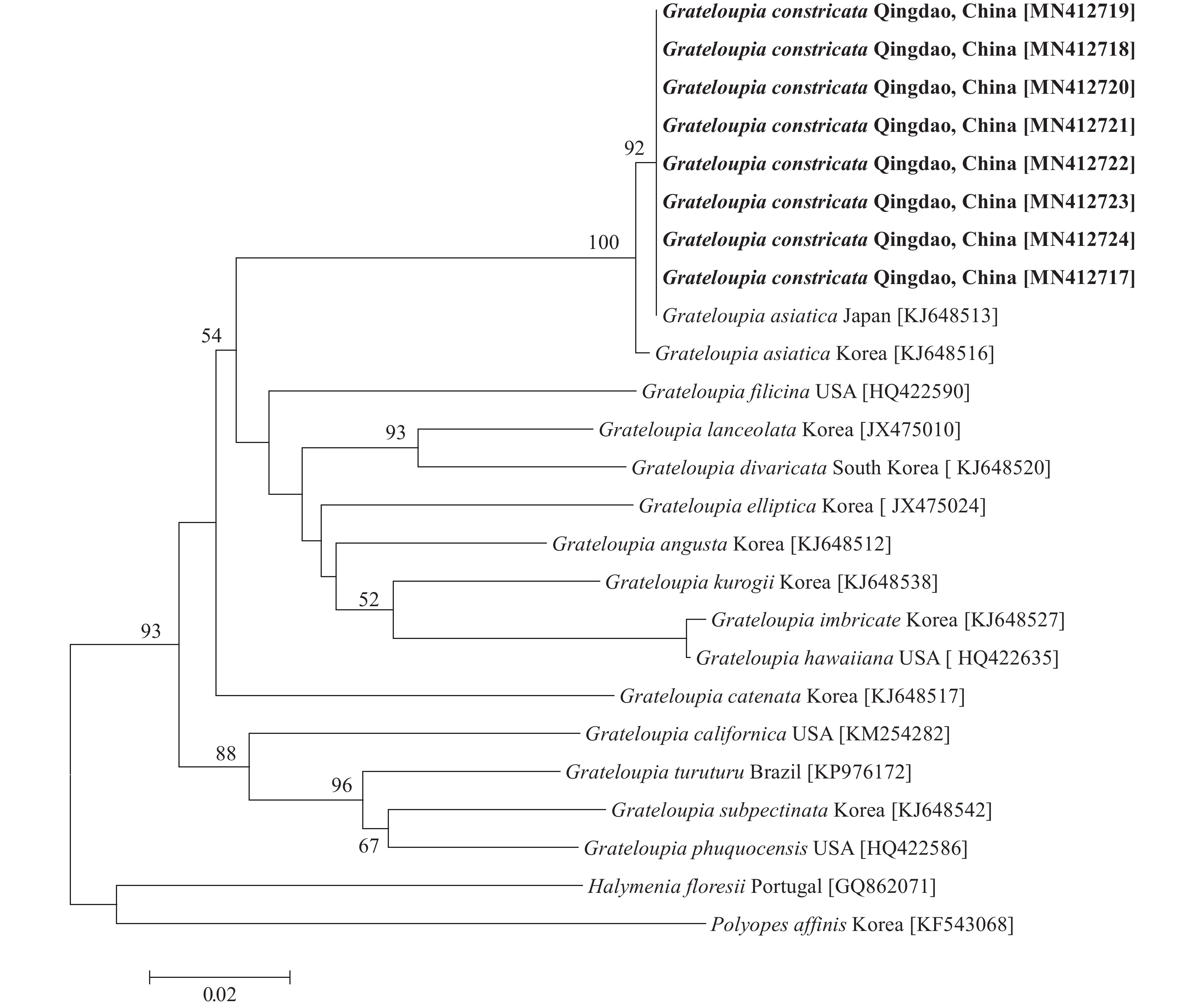
图 6 缢基蜈蚣藻基于rbcL基因的ML系统树分支上的数字展示基于1000次重复的支持率, 只显示支持率大于50%的数值, Halymenia floresia Malaysia和Polyopes constrictus Australia作为外群, 粗体字为本文研究种; 下同Figure 6. Maximum likelihood tree based on rbcL gene of G. constricataNumerals at internal nodes are bootstrap values (1000 replicates), only values above 50% bootstrap support are shown, Halymenia floresia Malaysia and Polyopes constrictus Australia were used as outgroups. Boldfaced words are the species studied in this study. The same applies belowCOⅠ基因序列分析 缢基蜈蚣藻的8个COⅠ基因序列长度均为629 bp, 比对校正后为616 bp。序列比对(MAGE5)结果显示如图 7, 8个缢基蜈蚣藻的基因序列间没有碱基差异, 与韩国的亚洲蜈蚣藻无碱基差异, 它们位于独立的发育枝中。缢基蜈蚣藻与蜈蚣藻(G. filicina)的碱基差异为65 bp (10.55%)。与作为外群的海膜属(Halymenia C. Agardh)的H. floresii和海柏属(Polyopes J. Agardh)的P. affinis的碱基差异分别为81 bp (13.15%)、89 bp (14.45%)。
![]() 图 7 缢基蜈蚣藻基于COⅠ基因的ML系统树Figure 7. Maximum likelihood tree based on COI gene of G. constricata
图 7 缢基蜈蚣藻基于COⅠ基因的ML系统树Figure 7. Maximum likelihood tree based on COI gene of G. constricata3. 讨论
对缢基蜈蚣藻的外形及生长状态进行观察, 发现其与亚洲蜈蚣藻在藻体颜色、大小、质地等方面完全一致(图 8A), 仅两者的分枝类型存在差异: 前者为1—3回羽状分枝; 后者为二叉分枝(表 2), 可能由于同种海藻因纬度、采集期及生态环境等不同, 导致二者在分枝上略有差异。1970年, Chiang[17]根据辅助细胞的位置及在其周围联络丝形成的辅助细胞瓶状体的特点, 将Grateloupia型的辅助细胞生殖枝丛结构作为蜈蚣藻的重要鉴别特征。Gargiulo等[18]认为, 根据构成辅助细胞生殖枝丛和果胞枝生殖枝丛主枝的细胞在数量上的区别, 可将Grateloupia型分为三种类型, 包括6cpb-5auxb型、5cpb-4auxb型和4cpb-3auxb型, 并且指出亚洲蜈蚣藻为典型的Grateloupia(6cpb-5auxb)型。本研究发现, 缢基蜈蚣藻为Grateloupia(6cpb-5auxb)型, 与亚洲蜈蚣藻为同一类型。二者仅在藻体分枝部分有较小的区别: 缢基蜈蚣藻1—3回羽状分枝, 小枝对生、互生或偏生, 基部缢缩; 亚洲蜈蚣藻主枝为二叉分枝, 小枝为羽状分枝, 小枝分布较稀疏(表 2)。由于同一种海藻的外部形态也因生态环境、海域及采集时间等不同而变异很大, 由此导致二者在外部形态上略有差异。
![]() 图 8 亚洲蜈蚣藻的外部形态(A)和《中国海藻志》中缢基蜈蚣藻的模式标本(B)Figure 8. External morphology of G. asiatica Kawaguchi et Wang (A) and typical specimen of G. constricata Li et Ding in Flora Algarum Marinarum Sinicarum (B)表 2 缢基蜈蚣藻和亚洲蜈蚣藻的形态比较Table 2. Comparison of morphological features between G. constricata and G. asiatic
图 8 亚洲蜈蚣藻的外部形态(A)和《中国海藻志》中缢基蜈蚣藻的模式标本(B)Figure 8. External morphology of G. asiatica Kawaguchi et Wang (A) and typical specimen of G. constricata Li et Ding in Flora Algarum Marinarum Sinicarum (B)表 2 缢基蜈蚣藻和亚洲蜈蚣藻的形态比较Table 2. Comparison of morphological features between G. constricata and G. asiatic形态学特征Morphological characteristics 缢基蜈蚣藻G. comstricata 亚洲蜈蚣藻G. asiatica 外部形态External morphology 藻体直立, 红褐色或紫红色, 高10—30 cm, 主枝扁平; 1—3回羽状分枝, 小枝对生、互生或偏生, 基部缢缩 藻体直立, 紫红色, 高10—30 cm,主枝扁压, 基部缢缩, 呈圆柱状, 宽2—5 mm;主枝二叉分枝, 小枝不规则羽状分枝 质地Texture 软骨质 软骨质 皮层数Cortex (C) 7—10层 7—10层 四分孢子囊形态和大小Tetrasporangia morphology and size 长椭圆形: 36—52 μm×14—20 μm 长椭圆形: 40—50 μm×15—20 μm 髓部Medullary structure 非中空 非中空 配子体Gametophyte 雌雄异体 雌雄异体 分布Native regions 中国青岛 中国大连、青岛 辅助细胞生殖枝丛类型Auxiliary cell ampullae Grateloupia型(6cpb-5auxb) Grateloupia型(6cpb-5auxb) 文献Reference 本研究; 夏邦美, 2004 Kawaguchi et al., 2001 本研究中详细观察并记录了缢基蜈蚣藻孢子的早期发育及生活史。缢基蜈蚣藻的孢子发育类型为“间接盘状体”型, 过程为: 在孢子逸出后, 萌发形成萌发管, 萌发管内细胞发生分裂, 进入盘状体时期。该结果与亚洲蜈蚣藻孢子发育及生活史的研究结果一致[19]。近几年的序列分析分别研究了不同基因序列在种内的遗传范围, 为藻类学分类提供了直观的证据, 研究指出COⅠ基因序列分析种内遗传变异范围为0.0—1.6%。李芳等[2]在帚状蜈蚣藻的修订研究中认为rbcL基因序列在蜈蚣藻属的种内的遗传变异范围为0.0—1.0%。基于DNA分子分析结果显示, 8个缢基蜈蚣藻样本之间均无碱基差异, 它们与大连亚洲蜈蚣藻无碱基差异,与日本和韩国的亚洲蜈蚣藻的碱基差异属于种内差异, 因此认为缢基蜈蚣藻与亚洲蜈蚣藻为同一种。
图 8B为《中国海藻志》中缢基蜈蚣藻模式标本, 李伟新等仅根据藻体形态结构和生殖结构特征建立一个新种不具有说服力。本研究通过对缢基蜈蚣藻外部形态及内部结构的观察、早期发育及DNA分子分析的研究, 确定缢基蜈蚣藻与亚洲蜈蚣藻为同一种。根据优先法则, 将缢基蜈蚣藻修订为亚洲蜈蚣藻。本研究将为我国蜈蚣藻属的分类修订、物种多样性及《中国海藻志》的再版提供新资料。
-
![]()
图 1 缢基蜈蚣藻的外部形态
A. 四分孢子体(LNU17060512); B. 雌配子体(LNU17060506); C. 雄配子体(LNU17060565)
Figure 1. External morphology of G. constricata Li et Ding
A. Tetrasporangia (LNU17060512); B. Female gametophyte (LNU17060506); C. Male gametophyte (LNU17060565)
![]()
图 2 缢基蜈蚣藻囊果
A. 缢基蜈蚣藻羽枝基部存在缢缩(小箭头所示), 囊果在主枝和小枝上均有分布(c, 囊果); B. 主枝上的囊果及囊果孔展示(c, 囊果; ch, 囊果孔)
Figure 2. Branches of G. constricata Li et Ding
A. Branches of G. constricata Li et Ding with constricted at bases (small arrowheads) and cystocarps distributed on axes and lateral branches of G. constricata Li et Ding (c, cystocarp); B. Cystocarp distribution on axes(c, cystocarp; ch, cystocarp hole)
![]()
图 3 缢基蜈蚣藻内部结构
A、B. 藻体的横切面观(mf, 髓丝); C. 雌配子体的果胞枝生殖枝丛横切面观(1—6. 果胞枝生殖枝丛主枝; ca. 果胞); D. 雄配子体的精子囊; E、F. 雌配子体的辅助细胞生殖枝丛横切面观(1—5, 辅助细胞生殖枝丛主枝; ac, 辅助细胞); G、H. 囊果连续的发育过程; G. 幼小囊果; H. 成熟囊果; I. 果孢子放散; 切片标本号: LNU17060506、LNU17060516、LNU17060536、LNU17060565
Figure 3. The internal structure of G. constricata Li et Ding
A, B. Cross-section of the thallus (mf, medullary filament); C. Transection of carpogonial branch ampullae from female gametophyte (1—6, leader branch carpogonial branch ampullae; ca, carpogonium); D. Spermatangium of male gametophyte; E, F. Transection of auxiliary cell ampullae from female gametophyte (1—5, leader branch of auxiliary cell ampullae; ac, auxiliary cell); G, H. Cystocarp development; G. Little cystocarp; H. Developing cystocarp; I. Release of mature carpospores. Section number: LNU17060506, LNU17060516, LNU17060536, LNU17060565
![]()
图 4 缢基蜈蚣藻四分孢子体内部结构
A. 四分孢子囊表面观; B. 四分孢子囊母细胞及二分体 (tm, 四分孢子囊母细胞; dy, 二分体); C. 成熟的四分孢子囊。切片标本号: LNU17060512
Figure 4. The internal structure of tetrasporophyte of G. constricata Li et Ding
A. Tetrasporophyte surface view; B. Tetrasporoblas and formed dyad (tm, tetrasporoblas; dy, formed dyad); C. Etrasporangia cross division, Section number: LNU17060512
![]()
图 5 缢基蜈蚣藻果孢子的萌发过程
A. 藻体表面的囊果(ch, 囊果孔); B—E为荧光显微镜拍摄结果; B. 果孢子; C. 果孢子内原生质体迁移; D—E. 萌发管内细胞分裂过程; F. 盘状体时期; G. 盘状体融合的过程; H. 直立芽体形成; I. 直立枝
Figure 5. The development of carpospores in G. constricata
A. Carposporophytes on the surface of thalli (ch. cystocarp hole); B—E were taken with a fluorescence microscope; B. Carpospores; C. Transformation of protoplasts in the germ tube; D—E. Cell division in the germ tube; F. Discoid crust; G. Fusion of discoid crust; H. Upright branches; I. Growth of sporeling
![]()
图 6 缢基蜈蚣藻基于rbcL基因的ML系统树
分支上的数字展示基于1000次重复的支持率, 只显示支持率大于50%的数值, Halymenia floresia Malaysia和Polyopes constrictus Australia作为外群, 粗体字为本文研究种; 下同
Figure 6. Maximum likelihood tree based on rbcL gene of G. constricata
Numerals at internal nodes are bootstrap values (1000 replicates), only values above 50% bootstrap support are shown, Halymenia floresia Malaysia and Polyopes constrictus Australia were used as outgroups. Boldfaced words are the species studied in this study. The same applies below
![]()
图 7 缢基蜈蚣藻基于COⅠ基因的ML系统树
Figure 7. Maximum likelihood tree based on COI gene of G. constricata
![]()
图 8 亚洲蜈蚣藻的外部形态(A)和《中国海藻志》中缢基蜈蚣藻的模式标本(B)
Figure 8. External morphology of G. asiatica Kawaguchi et Wang (A) and typical specimen of G. constricata Li et Ding in Flora Algarum Marinarum Sinicarum (B)
表 1 缢基蜈蚣藻的标本号、采集地点和基因登录号
Table 1 Specimen No., collection information and GenBank accession number
标本编号Specimen No. 采集地点Collection information 基因登录号GenBank accession number rbcL COⅠ LNU2017060501 The First Swimming Beach, Qingdao, China MN412709 MN412717 LNU2017060502 The First Swimming Beach, Qingdao, China MN412710 MN412718 LNU2017060503 The First Swimming Beach, Qingdao, China MN412711 MN412719 LNU2017060504 Luxun Park, Qingdao, China MN412712 MN412720 LNU2017060505 Luxun Park, Qingdao, China MN412713 MN412721 LNU2017060506 Luxun Park, Qingdao, China MN412714 MN412722 LNU2017060507 Maidao, Qingdao, China MN412715 MN412723 LNU2017060508 Maidao, Qingdao, China MN412716 MN412724  下载: 导出CSV
下载: 导出CSV
表 2 缢基蜈蚣藻和亚洲蜈蚣藻的形态比较
Table 2 Comparison of morphological features between G. constricata and G. asiatic
形态学特征Morphological characteristics 缢基蜈蚣藻G. comstricata 亚洲蜈蚣藻G. asiatica 外部形态External morphology 藻体直立, 红褐色或紫红色, 高10—30 cm, 主枝扁平; 1—3回羽状分枝, 小枝对生、互生或偏生, 基部缢缩 藻体直立, 紫红色, 高10—30 cm,主枝扁压, 基部缢缩, 呈圆柱状, 宽2—5 mm;主枝二叉分枝, 小枝不规则羽状分枝 质地Texture 软骨质 软骨质 皮层数Cortex (C) 7—10层 7—10层 四分孢子囊形态和大小Tetrasporangia morphology and size 长椭圆形: 36—52 μm×14—20 μm 长椭圆形: 40—50 μm×15—20 μm 髓部Medullary structure 非中空 非中空 配子体Gametophyte 雌雄异体 雌雄异体 分布Native regions 中国青岛 中国大连、青岛 辅助细胞生殖枝丛类型Auxiliary cell ampullae Grateloupia型(6cpb-5auxb) Grateloupia型(6cpb-5auxb) 文献Reference 本研究; 夏邦美, 2004 Kawaguchi et al., 2001
下载: 导出CSV
-
[1] Wang H W, Kawaguchi S, Horiguchi T, et al. Reinstatement of Grateloupia catenata (Rhodophyta, Halymeniaceae) on the basis of morphology and rbcL sequences [J]. Phycologia, 2000, 39(3): 228-237. doi: 10.2216/i0031-8884-39-3-228.1
[2] Kawaguchi S, Wang H W, Horiguchi T, et al. A comparative study of the red alga Grateloupia filicina (Halymeniaceae) from the Northwestern Pacific and Mediterranean with the description of Grateloupia asiatica sp. nov [J]. Journal of Phycology, 2001, 37(3): 433-442. doi: 10.1046/j.1529-8817.2001.037003433.x
[3] 李芳, 姜朋, 赵树雨, 等. 帚状蜈蚣藻的修订研究—基于形态观察和rbcL序列分析 [J]. 水生生物学报, 2016, 40(6): 1249-1256. doi: 10.7541/2016.163 Li F, Jiang P, Zhao S Y, et al. Research on the revision of Grateloupia fastigiata Li et Ding (Rhodophyta, Halymeniaceae) based on morphological observations and rbcL sequence analyses [J]. Acta Hydrobiologica Sinica, 2016, 40(6): 1249-1256. doi: 10.7541/2016.163
[4] 刘芳, 田依琳, 王宏伟, 等. 对枝蜈蚣藻的修订研究—基于形态观察和rbcL序列分析 [J]. 水生生物学报, 2017, 41(6): 1276-1284. Liu F, Tian Y L, Wang H W, et al. Research on the revision of Grateloupia didymecladia Li et Ding (Rhodophyta, Halymeniaceae) based on morphological observations and rbcL sequence analyses [J]. Acta Hydrobiologica Sinica, 2017, 41(6): 1276-1284.
[5] 夏邦美. 中国海藻志 [A]. 第二卷 红藻门, 第三册 隐丝藻目 [C]. 北京: 科学出版社, 2004: 57-59. Xia B M. Flora Algarum Marinarum Sinicarum [A]. Tomus II Rhodophyta. Part III. Cryptonemiales [C]. Beijing: Science Press, 2004: 57-59.
[6] De Clerck, Gavio O B, Fredericq, et al. Systematics of Grateloupia filicina (Halymeniaceae, Rhodophyta), based on rbcL sequence analysis and morphological evidence, including the reinstatement of G. minima and the description of G. capensis sp. nov. [J]. Journal of Phycology, 2005, 41(2): 391-410. doi: 10.1111/j.1529-8817.2005.04189.x
[7] Nelson W A, Kim S Y, Boo S M, et al. The first record of Grateloupia subpectinata from the New Zealand region and comparison with G. prolifera, a species endemic to the Chatham Islands [J]. Botanica Marina, 2013, 56(5-6): 507-513.
[8] Sherwood A R, Presting G G. Universal primers amplify a 23S rDNA plastid marker in eukaryotic algae and cyanobacteria [J]. Journal of Phycology, 2007, 43(3): 605-608. doi: 10.1111/j.1529-8817.2007.00341.x
[9] Wang H W, Kawaguchi S, Horiguchi T, et al. A morphological and molecular assessment of the genus Prionitis J. Agardh (Halymeniaceae, Rhodophyta) [J]. Phycological Research, 2001, 49(3): 251-261. doi: 10.1111/j.1440-1835.2001.tb00255.x
[10] Miller K A, Hughey J R, Gabrielson P W, et al. Research note: First report of the Japanese species Grateloupia lanceolata (Halymeniaceae, Rhodophyta) from California, USA [J]. Phycological Research, 2009, 57(3): 238-241. doi: 10.1111/j.1440-1835.2009.00542.x
[11] Lee J I, Kim H G, Geraldino P J L, et al. Molecular classification of the genus Grateloupia (Halymeniaceae, Rhodophyta) in Korea [J]. Algae, 2009, 24(4): 231-238. doi: 10.4490/ALGAE.2009.24.4.231
[12] Yang M Y, Kim M S. Taxonomy of Grateloupia (Halymeniales, Rhodophyta) by DNA barcode marker analysis and a description of Pachymeniopsis volvita sp. nov. [J]. Journal of Applied Phycology, 2015, 27(3): 1373-1384. doi: 10.1007/s10811-014-0432-1
[13] Sherwood A R, Sauvage T, Kurihara A, et al. The Hawaiian Rhodophyta Biodiversity Survey (2006-2010): a summary of principal findings [J]. BMC Plant Biology, 2010, 10(1): 258. doi: 10.1186/1471-2229-10-258
[14] Yu L, Wang H W, Luan R X. Grateloupia tenuis Wang et Luan sp. nov. (Halymeniaceae, Rhodophyta): a new species from South China Sea based on morphological observation and rbcL gene sequences analysis [J]. Biomed Research International, 2013(3): 560163.
[15] Wang H W, Guo S R, Zhang X M, et al. Re-identifying of the Grateloupia yangjiangensis (Rhodophyta, Halymeniaceae) on the basis of morphological observations, life history and rbcL sequence analyses [J]. Acta Oceanologica Sinica, 2014, 33(4): 77-84. doi: 10.1007/s13131-014-0450-5
[16] Koichiro T, Daniel P, Nicholas P, et al. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods [J]. Molecular Biology and Evolution, 2011, 28(10): 2731-2739. doi: 10.1093/molbev/msr121
[17] Chiang Y M. Morphological studies of the red algae of the family Cryptonemiaceae [J]. University of California Publications in Botany, 1970(58): 1-95.
[18] Gargiulo G M, Morabito M, Manghisi A. A re-assessment of reproductive anatomy and postfertilization development in the systematics of Grateloupia (Halymeniales, Rhodophyta) [J]. Cryptogamie, Algologie, 2013, 34(1): 3-35. doi: 10.7872/crya.v34.iss1.2013.3
[19] 王宏伟, 李雅卓, 曹翠翠, 等. 亚洲蜈蚣藻(Grateloupia asiatica Kawaguchi et Wang)孢子发育及生活史的研究 [J]. 辽宁师范大学学报(自然科学版), 2014(2): 246-251. Wang H W, Li Y Z, Cao C C, et al. A study on the spore development and life cycle of Grateloupia asiatica Kawaguchi et Wang (Halymeniaceae, Rhodophyta) [J]. Journal of Liaoning Normal University (Natural Science Edition), 2014(2): 246-251.

计量
- 文章访问数: 1790
- HTML全文浏览量: 416
- PDF下载量: 58