
STUDY ON DOUBLE FLUORESCENCE MARKING OF SPINIBARBUS SINENSIS JUVENILE
-
摘要: 为探究盐酸四环素(TCH)和茜素红S(ARS)对中华倒刺鲃(Spinibarbus sinensis Bleeker)的影响及标记效果, 丰富鱼类荧光标记模式, 研究使用TCH(100—500 mg/L)和ARS(100—300 mg/L)对中华倒刺鲃幼鱼进行双重浸染荧光标记, 实验共设置5个处理组和1个对照组, 浸染时间均为24h。结果显示, 经90d的养殖实验后, 标记鱼的耳石(包括矢耳石和星耳石)、倒刺、鳍条和鳍棘均能检测到双重荧光标记环, 且TCH产生的黄色荧光环比ARS产生的红色荧光环更接近骨质结构内部。较高浓度处理组的矢耳石(≥300 mg/L TCH和≥150 mg/L ARS)、星耳石(≥300 mg/L TCH 和 ≥200 mg/L ARS)和倒刺(400—500 mg/L TCH 和150—300 mg/L ARS)中均能检测到明显的标记环(n≥2), 但所有处理组的侧线鳞和非侧线鳞的荧光标记不明显(0≤n≤1)。经200—500 mg/L TCH和150—300 mg/L ARS处理的鳍条, 及经300—500 mg/L TCH和200—300 mg/L ARS处理的鳍棘中可以同时检测到明显的TCH和ARS的标记环(n≥2)。此外, 在整个实验中各处理组标记鱼与对照组相比, 在生长和存活率方面均无显著性差异(P>0.05)。结果表明, 使用TCH和ARS双重标记中华倒刺鲃幼鱼是可行的。双重荧光标记方法在水生生物标记回捕实验及实验性生物学研究方面具有潜在的应用前景。Abstract: In order to explore the effects of tetracycline hydrochloride (TCH) and alizarin red S (ARS) on Spinibarbus sinensis and their marker effects, and to enrich the fluorescent marking patterns of fish, TCH (from 100 to 500 mg/L) and ARS (from 100 to 300 mg/L) were used for double immersion marking juvenile Spinibarbus sinensis with 6 experimental groups for 24h. The results indicated that 24h double immersion produced detectable double marks in otoliths (sagittae and asteriscus), barbs, fin rays, and fin spines after 90 days in the laboratory growth experiment, with the exception of parts of scales. Yellow fluorescent rings produced by TCH were closer to the inside of the bony structures than red fluorescent rings produced by ARS. Sagittae (≥300 mg/L TCH and ≥150 mg/L ARS), asteriscus (≥300 mg/L TCH and ≥200 mg/L ARS), and barbs (400—500 mg/L TCH and 150—300 mg/L ARS) showed acceptable fluorescent marks at higher concentrations (n≥2). The fluorescence marking of lateral scales and non-lateral scales were not obvious in all treatment groups (0≤n≤1). Fin rays treated with 200—500 mg/L TCH and 150—300 mg/L ARS and fin spines treated with 300—500 mg/L TCH and 200—300 mg/L ARS simultaneously had both acceptable TCH and ARS marks (n≥2). In addition, double fluorescence labeling did not impact fish survival or growth (P>0.05) in the present study. The results suggest that double immersion with TCH and ARS is suitable for double mass-marking juvenile S. sinensis, and these double marks are useful in the experimental development of biological research or restocking methodologies.
-
湖泊富营养化及其引发的沉水植被退化仍然是我国众多湖泊目前面临的一个主要问题。由于沉水植物在湖泊生态系统中发挥着重要的生态功能(初级生产者, 维持生物多样性和清水稳态, 稳定底质环境等[1]), 沉水植被的退化将引发一系列水生态和水环境问题, 比如引起湖泊的“草-藻”稳态转换[2]。因此, 开展沉水植被修复是很多富营养化湖泊生态修复中的一项重要工作, 并在很多湖泊中得到实施, 比如武汉东湖[3]、杭州西湖[4]和荆州洪湖[5]等。但是, 在污染严重的水体中, 沉水植物的自然恢复是十分困难的, 一般而言, 湖泊低透明度和水下植物繁殖体的缺乏是影响水生植被恢复的主要限制因素[6, 7]。在这种情况下应尽快进行生态工程促进水生植被生境改善, 并积极开展水生植被重建与恢复工作[8]。就污染水体的低透明度而言, 水生植物必然会受到光照不足的影响, 一般认为环境光强持久或短时间显著低于植物光饱和点, 可以称为弱光逆境[9]。在弱光条件下, 首先通过降低光合作用效率抑制沉水植物的生长, 此外还通过抑制抗氧化酶活性影响沉水植物的生存[10, 11], 而大多数富营养化湖泊普遍表现出弱光或极度弱光的水下环境, 成为影响这些湖泊中沉水植被恢复的一大难题。除了水下光照环境, 底质条件对沉水植物的恢复亦有重要的影响, 底质是沉水植物的固定基质和营养来源, 对沉水植物生根、繁殖和生长产生不同程度的影响[12, 13], 研究表明富营养水体中肥沃底质对沉水植物造成胁迫, 使其生物量下降[14]。
洱海位于云南西北部的大理州境内, 是我国著名的高原淡水湖泊之一, 在洱海水面高程为1973.7 m时, 湖泊面积是249.8 km2。南部湖心平台(简称湖心平台)位于洱海南部湖心区域, 南起洱滨村, 北至才村和文笔村(25.64375—25.71297N, 100.21799—100.24011E), 南北两端是水深10 m以上的深水区, 东西两侧是水深10—13 m的深水带与沿岸带相隔, 中间形成一片水深6 m左右的暗滩区域。湖心平台曾经是洱海沉水植物成片连续分布的最大区域, 在洱海沉水植被演替过程中具有非常重要的地位(表 1)[15-17]; 自1998年洱海发生大面积水华后, 湖心平台区域沉水植被大幅退化[18, 19], 在2009年该区域未发现沉水植物生存[20]。如果能成功恢复湖心平台的沉水植被, 将大幅提升洱海沉水植物群落的分布面积和生态功能。
Table 1. Changes of aquatic vegetation community in the lake center platform of Erhai Lake时间
Time分布面积
Distribution area (km2)在全湖水生植被面积的百分比
Distribution proportion (%)群落
Community优势种
Dominant species1975—1983 金鱼藻群落 金鱼藻、苦草、黑藻 1981—1983 23.0 29.76 微齿眼子菜、金鱼藻群落 金鱼藻、微齿眼子菜 1998 33.4 34.80 微齿眼子菜群落 微齿眼子菜、苦草、黑藻 2015年以来, 随着洱海环境保护力度的加大, 洱海水质呈现出逐年向好的发展趋势, 透明度逐年提高, 湖心平台水下光环境基本达到沉水植物的光补偿点, 理论上有沉水植物恢复的可能性, 但是没有出现自然恢复的现象, 因此我们希望通过人工强化的修复措施实现该区域的沉水植被重建。这里面有两个问题: (1)选用哪种沉水植物; (2)如何通过改善局部环境来帮助沉水植物应对弱光胁迫。苦草(Vallisneria natans)是水鳖科(Hydrocharitaceae)苦草属(Vallisneria)沉水草本植物, 无直立茎, 叶基生, 条形[21], 生长区域广泛, 在淡水湖泊、沟渠、池塘和内河航道的静水或流动水体中均能很好地生长[22]。苦草对弱光环境具有很强的耐受能力, 因此成为洱海沉水植物群落中分布下限最深的物种[23], 并且在1980s中后期以来, 苦草成为洱海水生植被的优势种之一[24], 所以我们选择了对弱光环境耐受能力强的苦草作为湖心平台区域沉水植被恢复的尝试物种。另外, 就湖心平台底泥中富集了大量有机碎屑的情况而言, 改善底质环境理论上对沉水植被的恢复有利, 所以我们选择了黏土作为底质的处理, 以原位淤泥作为对照的方法, 探究底质改善对湖心平台水域沉水植被恢复的影响。对当地水生植被的恢复可以起到很好的指导作用, 在洱海湖泊的生态修复中具有重要的意义。
1. 材料与方法
1.1 实验材料
实验用的苦草来自洱海苗圃基地, 苗圃基地邻于洱海北部, 由水深1.5 m水生植物培养池构成, 取洱海的苦草置于苗圃培养池, 通过培养扩增苦草数量。实验选用的是生长茂盛、健康和形态指标大体一致[株高: (65±5) cm, 鲜重: (10.5±0.5) g]的苦草, 并去除植株上的匍匐茎。
1.2 实验时间与地点
实验时间选择苦草在洱海的生长旺盛期, 从2018年6月7日到2018年8月7日, 共9周(63d)。
如图 1所示,实验地点选择在洱海南部湖心平台水深最浅的区域, 实验地点中心坐标(25.684234 N, 100.226728E)。
1.3 实验设计
实验以黏土底质作为处理组, 以淤泥底质(湖心平台原位底泥)作为对照组。淤泥是用彼德森采泥器采集湖心平台水底的底泥, 黏土采自苗圃基地, 是洱海岸边常见的泥土。黏土含水率20%, N含量0.11%, C含量2.75%, 有机质含量7.76%; 而淤泥含水率83%, N含量1.09%, C含量13.63%, 有机质含量20.52%。比较两种底泥, 淤泥含有更高浓度的C、N和有机质含量, 而黏土含水率低, 土质偏硬。
种植方式采用沉桶种植。种植桶是白色塑料桶(口直径: 35 cm, 底直径: 27 cm, 高: 33 cm), 在桶内装填底质(淤泥或黏土)15 cm, 将苦草按照每桶30株的密度均匀植于桶内, 每个实验处理分配20桶作为平行。种植桶沉入水底前, 用另一端绑着浮标(直径10 cm的白色泡沫球)的尼龙绳系在桶上, 绳长度7.5 m, 略大于实验地点的水深(约6.3 m), 保证种植桶不受浮标漂动影响, 然后缓慢沉入水底。
1.4 测量指标
水环境指标 在实验期间, 光合有效辐射、水温、pH、透明度、实验区水体总氮(TN)、氨氮(NH4-N)、硝态氮(NO3-N)、总磷(TP)和正磷酸盐(PO4-P)每1周测量1次。水温和pH用多参数水质分析仪YSI(Yellow Springs Instruments, Ohio, US)测量, 透明度用黑白透明度盘测量, 水底光合有效辐射使用水下照度计(Li-1400; Li-Cor Company, Lincoln, NE, U.S.A; UWQ-8342)于上午11—12点在苦草冠层深度测量, 水的消光系数用不同水深的光合有效辐射计算得来。总氮、氨氮、硝态氮、总磷和正磷酸盐根据标准方法测定[25]。
苦草生长和生理指标 在实验周期内, 每1周从对照组和实验组各随机取5个实验桶计数植株数量作为苦草存活量, 从5个桶内每桶随机取1株苦草带回实验室, 测量地上部分(叶片)和地下部分(根和茎)的长度, 经80℃烘干至恒重后测量各部分的生物量(干重)、游离氨基酸、可溶性糖、淀粉、碳和氮含量。植物组织碳和氮含量通过元素分析仪(Flash EA 1112, CE Instruments, Italy)测定; 将烘干的植物组织50 mg用5 mL的80%乙醇在80℃的水浴萃取20min, 重复2次, 离心后合并上清液, 并定容到5 mL用于测定总糖和氨基酸, 萃取后的残渣中加入1 mL 30%的高氯酸继续萃取120min, 离心10min后收集上清液用于测定淀粉含量。总糖的测定使用1 g/L的葡萄糖溶液做标准曲线, 用蒽酮试剂法测定[26], 游离氨基酸的测定用3 μg/mL的丙氨酸做标准曲线, 用茚满三酮—抗坏血酸法测定[27]。用5 g/L的淀粉溶液做标准曲线, 用碘试剂法测定淀粉含量[28]。
1.5 分析方法
使用SPSS 22软件进行统计分析, 采用双因素方差分析时间和底质因素对苦草存活数量和不同部位的生物量、长度的作用效应, 采用成组样本t检验实验组和对照组在游离氨基酸、可溶性糖、淀粉、碳、氮含量上是否有差异, 方差分析前所有数据进行正态分布和方差齐性检验, 如有必要时将数据进行转换以满足方差齐性检验。使用Origin 2017软件绘图。
2. 结果
2.1 水环境特征
在实验周期内, 实验区域水深6.1—6.6 m, 透明度1.5—2.0 m, 湖底光照8.3—15.8 μmol/(m2·s), 有研究报道苦草的光补偿点为9.4 μmol/(m2·s), 光饱和点200 μmol/(m2·s)[29](图 2), 湖底光照强度不能满足苦草的需光量, 光照不足是影响生长的主要限制因子, 在此区域苦草必然受到弱光胁迫的影响。另外, 实验区域水体水温22.1—24.6℃; pH 8.5—8.6; TN浓度0.56—0.98 mg/L, NH4-N浓度0.012—0.071 mg/L, NO3-N浓度0.031—0.052 mg/L; TP浓度0.034—0.048 mg/L, PO4-P浓度0.002— 0.015 mg/L, 湖心区域水质为Ⅲ类水, 按照地表水质标准(GB3838-2002), 属于Ⅲ类水。有研究报道苦草能在5—35℃温度条件下良好生长[30], 在氨氮<1.56 mg/L时, 现存量增加百分比略有增加, 而硝氮<25 mg/L时, 促进苦草生长, 在总磷浓度<3.2 mg/L时, 苦草的生产力可保持正增长[1], 在本实验地点水温和营养盐因素对苦草的生长并没有不利的影响。
![]() 图 2 水环境因子变化A. 苦草的光补偿点Figure 2. Changes in water environmental factorsA. Light compensation point of V. natans
图 2 水环境因子变化A. 苦草的光补偿点Figure 2. Changes in water environmental factorsA. Light compensation point of V. natans2.2 苦草存活数量
在实验期间, 苦草存活量显著性下降, 对照组从30株/桶下降到(10±2)株/桶, 实验组从30株/桶下降到(18±4)株/桶(图 3), 这说明弱光环境限制了苦草的生长, 导致了部分苦草死亡。双因素方差分析表明, 黏土组苦草存活株数显著性大于淤泥对照组(表 2, P<0.05), 说明对比于淤泥底质, 黏土更有利于苦草的存活, 然而2种底质苦草存活数仍明显少于初始种植数。
表 2 苦草存活数和不同器官生物量、长度的双因素方差分析Table 2. Two factor ANOVA table of survival number, biomass and length of different organs of V. natans (*P<0.05,** P <0.01, ***P<0.001)指标
Index方差变异百分比 Percentage (%) 底质 时间 底质×时间 误差 存活数
Survival number2.9** 61.7*** 7.1* 28.2 生物量
Biomass叶片 0.0 91.0*** 4.1*** 4.9 根茎 6.3*** 63.3*** 4.9 25.3 根茎/叶片 3.8*** 66.8*** 22.6*** 6.8 长度
Length叶片 0.2 55.1*** 1.5 43.1 根茎 0.0 65.5*** 5 29.6 根茎/叶片 0.9 58.4*** 5.7 35.1 2.3 弱光胁迫下底质对苦草形态指标的影响
在实验初始时, 单株苦草叶片的平均生物量为(0.96±0.04) g, 根茎的平均生物量为(0.27±0.01) g。在弱光环境下, 苦草生长明显受到抑制, 叶片生物量和根茎生物量都有显著性下降(表 2, P<0.05)。在实验期间, 实验组和对照组单株叶片的平均生物量分别下降至(0.10±0.03)和(0.10±0.02) g, 下降幅度分别是90%和90%; 根生物量在第1周下降趋势最迅速, 之后在2个实验组之间表现出差异, 对照组的下降趋势减缓, 实验组的变化是先上升后下降, 第9周, 实验组和对照组的根生物量分别是(0.13±0.04)和(0.06±0.01) g, 相比初始值下降幅度分别是51%和80%; 实验组和对照组单株苦草根茎和叶片的生物量比值显著性上升(表 2, P<0.05), 分别从(0.28±0.01)上升到(1.35±0.09)和(0.59±0.09), 并且叶片生物量的下降幅度远大于根茎(图 4)。双因素方差分析表明, 时间因素对叶片和根茎的生物量及两者的比值有显著性影响, 底质因素对根茎的生物量有显著性影响, 实验组根茎生物量大于对照组(表 2, P<0.05)。
在初始时, 苦草叶片长度平均值(51.7±3.7) cm, 根茎长度平均值(13.4±1.0) cm。在弱光环境下, 苦草叶片和根茎长度显著性下降(表 2, P<0.05), 第1周, 实验组的苦草叶片长度下降至(36.3±9.2) cm, 对照组的苦草叶片长度下降至(40.1±14.4) cm, 比初始值分别下降了30%和22%, 此后变化不显著; 第5周, 实验组苦草的根茎长度为(3.6±1.0) cm, 对照组苦草的根茎长度为(3.7±1.1) cm, 比初始值分别下降了73%和72%, 此后苦草根长的变化不显著; 苦草根茎和叶片长度的比值显著性下降(表 2, P<0.05), 实验组和对照组的根茎和叶片长度的比值分别从(0.26±0.00)下降到(0.12±0.01)和(0.11±0.01), 并且叶片长度的下降幅度远小于根茎(图 4)。双因素方差分析表明, 时间因素对叶片和根茎的长度及两者的比值都有显著性影响, 而底质因素对它们没有显著性效应, 即实验组和对照组之间没有显著性差异。
2.4 在弱光胁迫下两种底质中苦草的生理指标的变化
在初始时, 苦草叶片的C含量是37.2%, 根茎的C含量是36.2%, 在弱光环境下, 实验组和对照组的叶片和根茎的C含量均呈下降趋势, 在实验期间, 实验组和对照组的叶片C含量分别下降至30.6%和32.1%, 相比于初始值其下降幅度分别是17.8%和13.8%; 实验组和对照组的根茎C含量分别下降到28.3%和30.2%, 相比于初始值其下降幅度分别是21.8%和16.5%; 叶片C含量的下降幅度与根茎相差不大, 实验组和对照组根茎和叶片的C含量比值在实验期间的变化也不明显(图 5)。根茎和叶片的C含量在实验组和对照组之间没有显著性差异(表 3)。
![]() 图 5 苦草的碳(C)和氮(N)含量变化Figure 5. Changes of carbon (C) and nitrogen (N) contents in V. natans表 3 样本t 检验苦草生理指标在两种底质间的差异显著性分析Table 3. Significant analysis on the difference of physiological indexes of V. natans between two kinds of sediment by t test
图 5 苦草的碳(C)和氮(N)含量变化Figure 5. Changes of carbon (C) and nitrogen (N) contents in V. natans表 3 样本t 检验苦草生理指标在两种底质间的差异显著性分析Table 3. Significant analysis on the difference of physiological indexes of V. natans between two kinds of sediment by t test部位Position 生理指标
Physiological index底质(P值)
Sediment (P)叶片Leaf C含量 0.943 N含量 0.047 游离氨基酸含量 0.168 可溶性糖含量 0.501 淀粉含量 0.306 根茎Rhizome C含量 0.575 N含量 0.000 游离氨基酸含量 0.002 可溶性糖含量 0.681 淀粉含量 0.016 在初始时, 苦草叶片的N含量是0.94%, 根茎的N含量是0.91%, 在弱光环境下, 叶片和根茎的N含量呈上升趋势, 在实验期间, 实验组和对照组叶片N含量分别下降到2.97%和3.07%, 相比于初始值上升幅度分别是215%和225%; 实验组和对照组根茎的N含量分别下降到1.05%和1.75%, 相比于初始值上升幅度分别是15.1%和92.3%; 叶片N含量的上升幅度远大于根茎, 实验组和对照组根茎和叶片的N含量比值分别从0.97下降到0.35和0.57(图 5)。实验组叶片和根茎的N含量均显著性大于对照组(表 3, P<0.05)。
在初始时, 苦草叶片和根茎的游离氨基酸含量分别是0.27和0.21 mg/g, 在弱光环境下, 叶片和根茎的游离氨基酸含量都呈上升趋势, 在实验期间, 实验组和对照组叶片的氨基酸含量分别上升到1.22和1.30 mg/g, 比初始值上升了347%和374%; 实验组和对照组根茎的氨基酸含量分别上升到0.32和0.62 mg/g, 相比于初始值上升了49%和190%; 叶片的氨基酸含量的上升幅度大于根茎, 根茎和叶片的氨基酸含量比值下降, 实验组和对照组分别从0.78下降到0.26和0.48(图 6)。样本t检验表明, 实验组根茎的氨基酸含量显著低于对照组(表 3, P<0.05)。
![]() 图 6 苦草的游离氨基酸、可溶性糖和淀粉含量变化Figure 6. Changes of free amino acid, soluble carbohydrate and starch content in V. natans
图 6 苦草的游离氨基酸、可溶性糖和淀粉含量变化Figure 6. Changes of free amino acid, soluble carbohydrate and starch content in V. natans在初始时, 苦草叶片的可溶性糖含量是16.2 mg/g, 根茎的可溶性糖含量是14.1 m/g, 在弱光环境下, 叶片的可溶性糖含量在第1周略有上升随后迅速下降, 在实验结束时, 实验组和对照组叶片的可溶性糖含量分别下降至8.2和9.4 mg/g, 相比于初始值下降幅度分别是49%和42%; 根茎的可溶性糖含量呈先上升后下降的趋势, 实验组根茎的可溶性糖含量上升至29.3后下降至9.0 mg/g, 对照组根茎的可溶性糖含量先上升至31.9后下降至5.6 mg/g; 实验组根茎和叶片的可溶性糖含量比值从0.87上升至3.9随后下降至1.1, 对照组从0.87上升至5.4随后下降至0.60(图 6)。叶片和根茎的可溶性糖含量在实验组和对照组之间没有显著性差异(表 3)。
在初始时, 苦草叶片和根茎的淀粉含量分别是239和286 m/g, 在弱光环境下, 苦草叶片和根茎的淀粉含量持续下降, 在实验结束时, 实验组和对照组叶片的淀粉含量分别下降到19和9 mg/g, 相比初始值下降幅度分别是92%和96%; 实验组和对照组根茎的淀粉含量分别下降到212和133 mg/g, 相比根茎初始值下降幅度分别是26%和53%; 叶片淀粉含量的下降幅度大于根茎, 根茎和叶片的淀粉含量比值呈上升趋势, 实验组从1.2上升至11.4, 对照组从1.2上升至15(图 6)。叶片淀粉含量在实验组和对照组之间没有显著性差异, 而实验组根茎的淀粉含量显著大于对照组(表 3, P<0.05)。
3. 讨论
3.1 弱光环境对苦草影响
植物通过光和色素吸收光能, 将无机碳和水转化为有机物, 用于自身合成和代谢, 湖泊底部良好的光照是沉水植物生长的基本前提。在洱海湖心平台所测的湖底光合有效辐射强度平均值是10.5 μmol/(m2·s), 略高于他人研究报道的苦草光补偿点9.4 μmol/(m2·s) [29], 所以弱光是苦草植被恢复的主要限制因子。
在实验结束时, 实验桶内的苦草存活数量已稳定下来, 存活数量显著小于初始种植数量。相比于初始状态, 单株苦草的叶生物量、根生物量及根长和叶长均显著性下降。有研究表明, 随水深梯度的增加苦草生物量和和无性系分株数显著性下降, 而株高呈先增加后降低的趋势[31], 本研究结果与其一致。一方面苦草株高和生物量下降, 使其长期处于弱光胁迫下;另一方面低光照会减弱沉水植物的胁迫耐受能力[32], 两者共同解释了前面苦草存活株数小于初始种植株数的实验结果。苦草叶片的游离氨基酸含量上升, 而C含量、淀粉含量和可溶性糖含量下降, N含量上升; 根茎的游离氨基酸含量上升, 可溶性糖的含量先上升后下降, C含量下降, N含量上升。有文献报道水下光强的大幅降低能导致沉水植物出现生理胁迫, 包括植物组织N含量增加, 碳源性次生化学产物的含量下降, 比如可溶性糖和酚类[33, 34]。苦草的生理响应可见苦草出现一定程度的胁迫反应。
在苦草形态响应弱光环境的过程中, 苦草对不同器官的资源重新分配有一定的差异。相比于初始状态, 苦草根茎和叶片的生物量比值上升, 而根茎与叶片的长度比值下降, 实验结果与张素娟的研究结果一致[35]。两种器官相对比, 叶片的生物量损失相对更大, 而长度降低幅度相对较小, 这也反映了苦草为适应弱光环境发生了一些形态上的改变, 因为苦草植株的高度很大程度取决于叶片的长度, 并且苦草需要根的定植才能存活。弱光胁迫可以引起细胞活性氧的产生和积累, 诱导相关酶系统活性增强[11, 36], 氨基酸是合成酶的物质基础, 有许多文献表明游离氨基酸含量是指示环境胁迫的重要生理指标[37, 38]。与根茎相比, 苦草叶片的游离氨基酸含量上升幅度更大, 并且根茎和叶片的氨基酸含量比值大幅降低, 这说明苦草氨基酸的合成主要发生在叶片细胞中, 叶片细胞的胁迫响应更显著。可溶性糖是植物能直接利用的能源物质[39], 在弱光胁迫下, 光合作用合成糖类和ATP的量都很低, 但用于合成氨基酸的能量和糖类的需求增多, 植株需要消耗淀粉分解成糖以提供能量基础, 所以植株储存的碳水化合物的含量与植物对不良环境的耐受能力息息相关[40]。苦草叶片的可溶性糖含量大幅降低, 而根茎的可溶性糖含量先升高后下降, 根茎和叶片的可溶性糖含量比值也先升高后下降, 这也表明了根茎细胞对逆境的抵抗能力相对更强; 根茎与叶片的淀粉含量比值大幅升高, 说明叶细胞受到弱光胁迫造成的能量损耗远大于根茎的细胞, 当能量不能满足代谢损耗时, 逆境抵抗能力也因此下降, 造成细胞死亡。
3.2 在弱光环境下不同底质对苦草影响
在实验期间, 淤泥和黏土2种底质下苦草存活数量表现出显著性差异, 在实验结束时, 实验组(黏土底质)和对照组(淤泥底质)存活数量平均值分别是18和10株/桶, 实验组大于对照组。并且底质因素对苦草根茎生物量也显著性影响, 表现为实验组大于对照组。虽然有研究报道在光照合适的条件下, 相比于黏土底质, 苦草更适合在高营养的湖泥中生长[41, 42], 但是在弱光环境下, 底质对沉水植物生长影响的研究却比较少, 并且, 李文朝[43]的研究认为, 底质条件是否适合沉水植物生长, 不仅取决于底质本身, 很大程度受制于湖水温度和水深, 30℃且水深远大于l m时, 底泥和底层湖水中的厌氧环境会引起沉水植物烂根, 另外, 底泥的物理结构与沉水植物的扎根能力和扎根深度密切相关[44], 密度大、孔隙小的黏土在根系的附着上占优势。
在生理特征上, 叶片的N含量及根茎的N含量和氨基酸含量在不同底质处理下有显著性差异, 均表现为实验组小于对照组, 这说明实验组苦草受到程度更小的生理胁迫, 并且不同底质条件苦草根茎的淀粉含量比较, 实验组组显著性大于对照组, 这也说明了实验组苦草受到的胁迫损伤相对更小。以上2点均为2种底质条件下苦草存活率和形态特征的差异提供了一定的解释。沉水植物可以直接从底质中吸收氮等营养盐, 并且, 由于氨态氮浓度在底泥中往往比湖水高, 沉水植物吸收的氮主要来源于底泥[12]。相对于黏土底质, 淤泥底质的N含量约为黏土的9.9倍, 苦草叶片、根茎的N含量和氨基酸含量在不同底质之间的差异与两种底质N含量的极大差异有关, 另外底泥中有机物含量和底泥营养含量密切相关, 过多的有机物分解可造成底泥缺氧和还原性的状态, 对植物有毒害作用[45, 46], 这也为本次实验结果提供了一个较合理的解释。
综合来看, 在弱光环境下对比于淤泥对照组, 黏土实验组的苦草在生理上表现出幅度更小的胁迫响应, 同时受到相对更小的胁迫损伤, 使其根茎的生物量显著大于对照组, 死亡植株数下降, 存活数量显著大于对照组。所以使用黏土代替本底淤泥的措施对苦草的恢复有利。
3.3 苦草恢复的可行性分析和恢复策略
因为对照组底质是在湖心平台退化区直接采取的底泥, 所以从对照组的苦草存活数量和形态特征来看, 虽然在实验结束时, 苦草留有一定的存活数量, 但是苦草形态特征并没有明显的上升现象, 并且实验并未发现有大规模的分株现象, 所以此时的退化区环境并不适合直接进行沉水植物恢复工程, 应该优先考虑改善环境特征。因为光照是限制苦草生长的主要限制性因子, 应该采取降低水位和水体营养盐浓度以增加透明度等提高水下光合有效辐射的措施, 或者寻找或驯化出对弱光环境适应性更强的沉水植物品种等。另外, 虽然底质因素对苦草存活数的方差变异解释度只有2.9%, 对根茎的生物量的方差变异解释度只有6.3%, 但是都有显著性影响, 所以, 底质因素也是影响苦草恢复的重要因子, 从淤泥对照组和黏土实验组的对比来看, 黏土实验组存活数量大于淤泥对照组, 黏土底质条件下苦草拥有更高的存活率, 并且黏土底质条件下苦草受到的胁迫损伤相对更小, 所以使用黏土实施底质改善措施对苦草的恢复有积极的作用。
4. 结论
(1)在平均光强略高于苦草光补偿点的极度弱光[8.3—15.8 μmol/(cm2·s)]和水深6.1—6.6 m的洱海湖底, 桶栽的苦草部分死亡, 存活数量下降, 并在形态特征和生理特征上均表现出胁迫响应, 其单株生物量和株高下降, N含量和游离氨基酸含量上升, C含量和淀粉含量下降。(2)苦草不同器官对弱光环境的响应有所差异, 叶片(地上部分)受到的胁迫影响大于根茎(地下部分)。(3)苦草对弱光环境的响应在黏土和湖底淤泥2种不同底质条件下有显著性差异, 苦草在黏土底质上表现出更小的胁迫反应和更高的存活数量, 2种底质相比较, 黏土更适合作为苦草恢复的底质条件。研究表明, 在洱海当前的水质环境下有希望结合局部的底质改善来实现在湖心平台的沉水植物恢复。
-
![]()
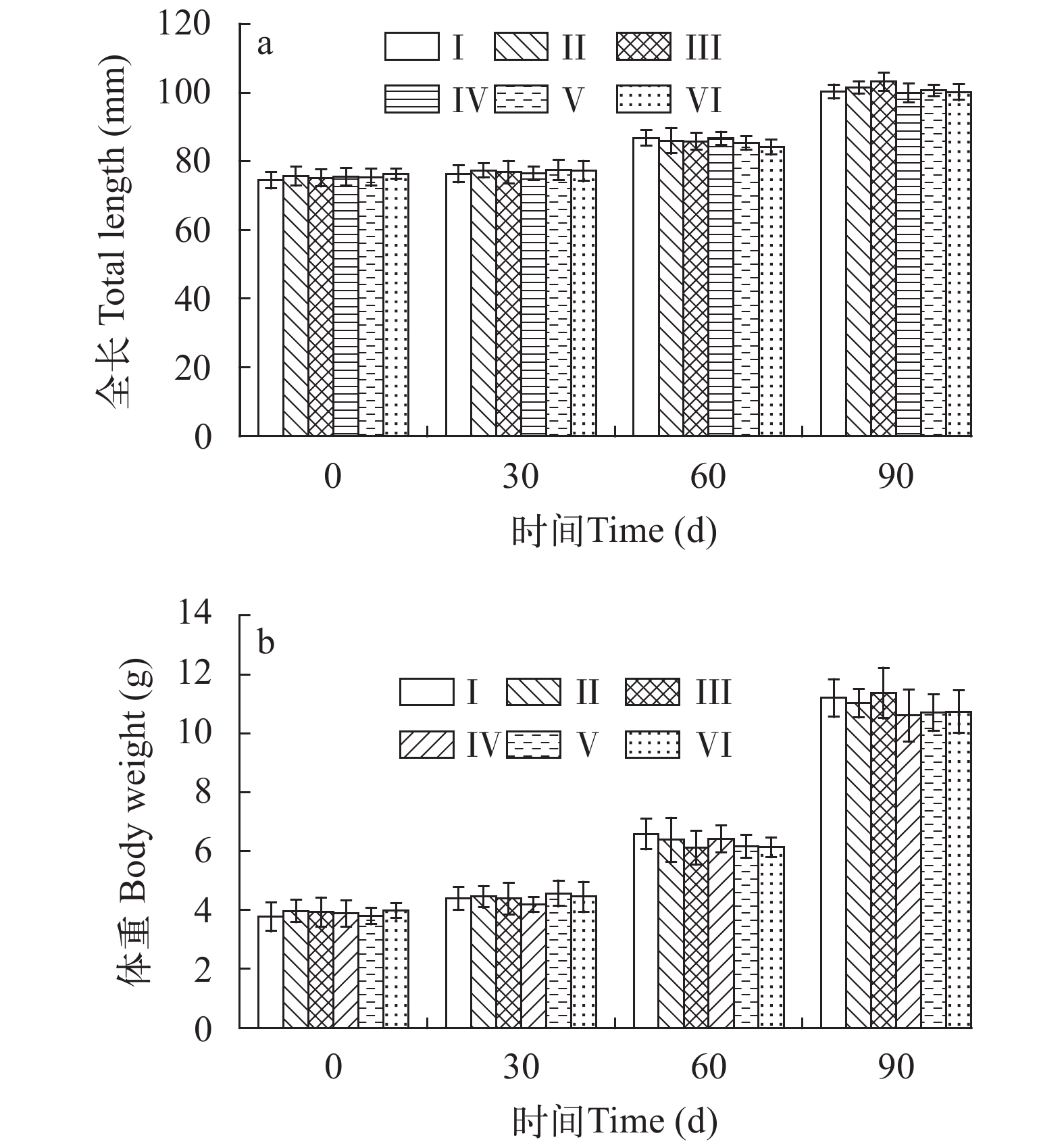
图 1 双重标记后6个实验组中华倒刺鲃平均全长(a)和体重(b)
“0—30d”表示生长实验的第一阶段;“30—90d”表示生长实验的第二阶段; 误差线代表标准误, P>0.05
Figure 1. Mean total length (a) and body weight (b) of S. sinensis from six experimental groups after double fluorescent marking
“0—30d”represent the first stage of growth experiment; “30—90d”represent the second stage of growth experiment; Vertical bars represent standard error, P>0.05
![]()
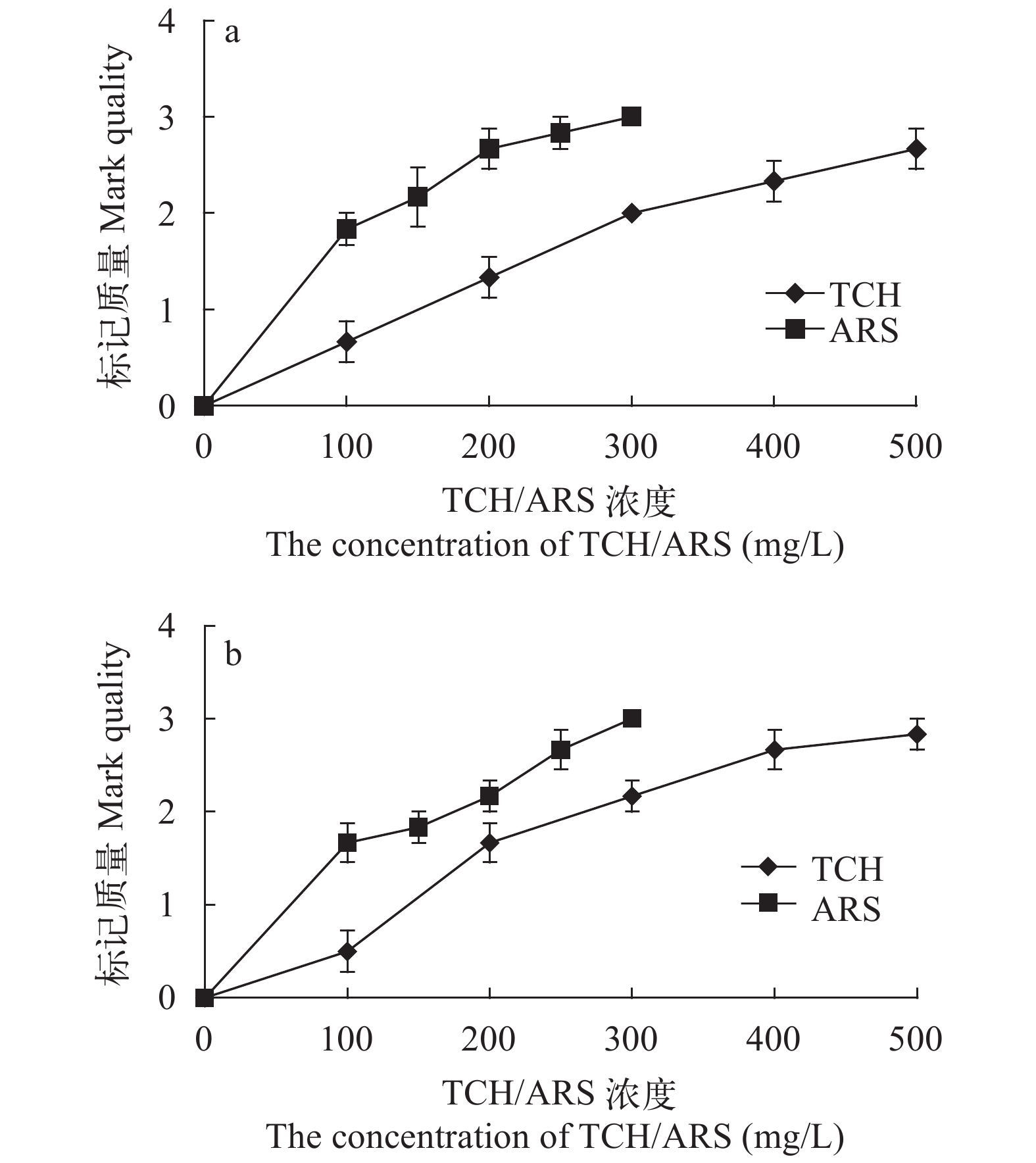
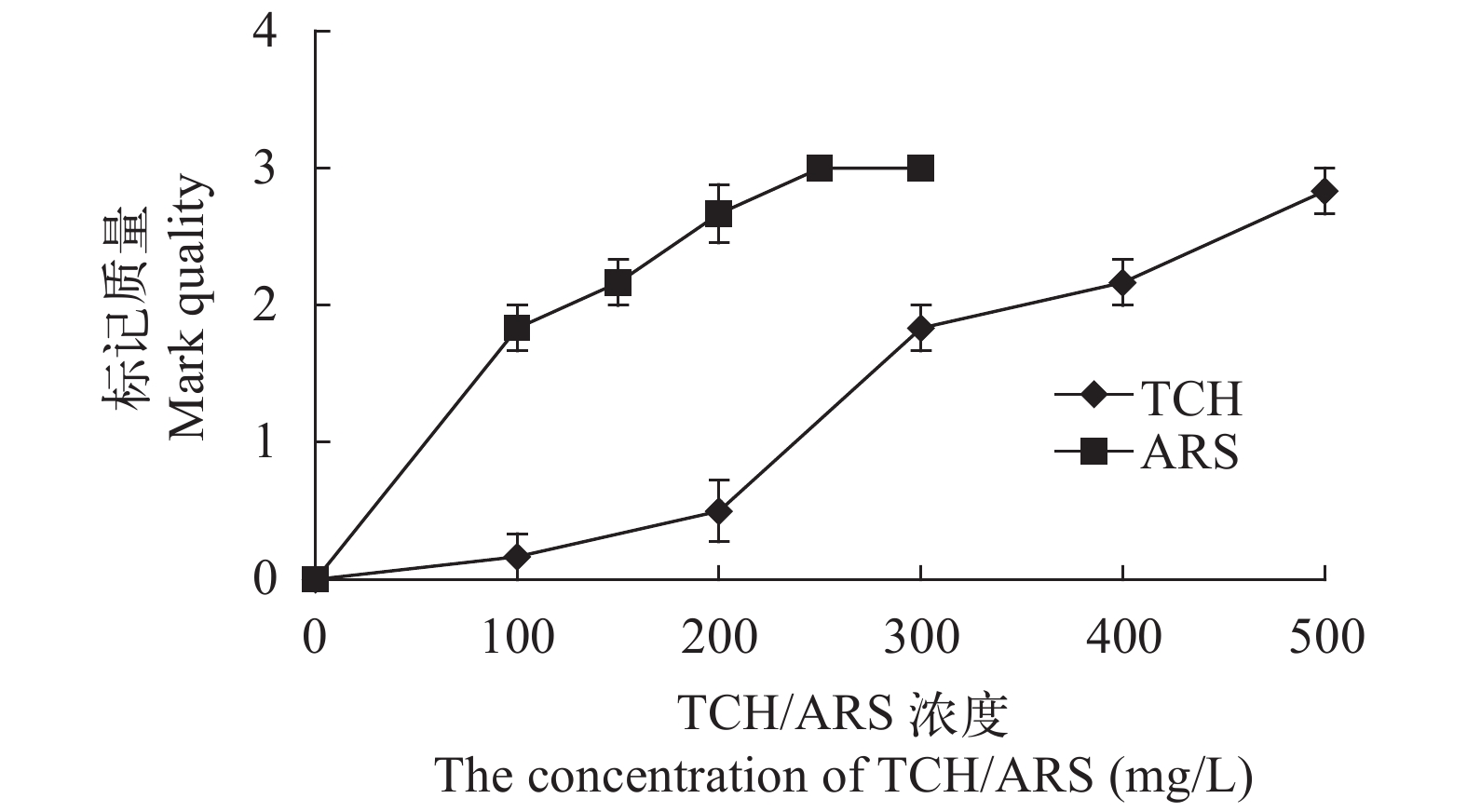
图 2 TCH和ARS荧光染料对中华倒刺鲃幼鱼矢耳石(a)和星耳石(b)的标记质量
Figure 2. The mark quality of TCH and ARS fluorochrome dyes on the sagittae (a) and asteriscus (b) of S. sinensis
![]()
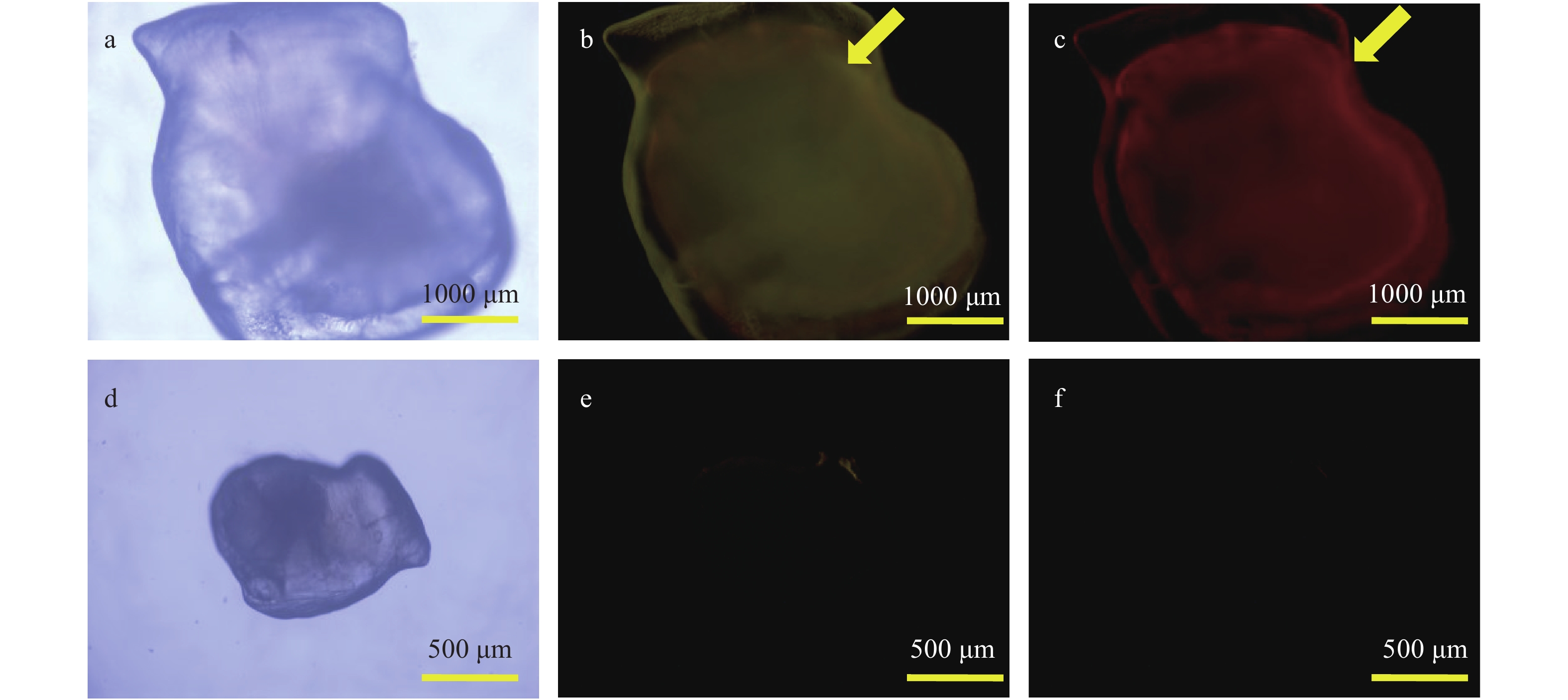
图 3 不同光源下中华倒刺鲃幼鱼矢耳石的双重荧光标记效果
a, b, c为处理组; d, e, f为对照组. 透射光(a, d); 蓝光(b, e); 绿光(c, f); 下同
Figure 3. The effect of double fluorescent marking in sagittae of S. sinensis larval observed with different light sources
a, b, c are treatment groups; d, e, f are control group. Transmitted light (a, d); blue light (b, e); green light (c, f); the same applies below
![]()
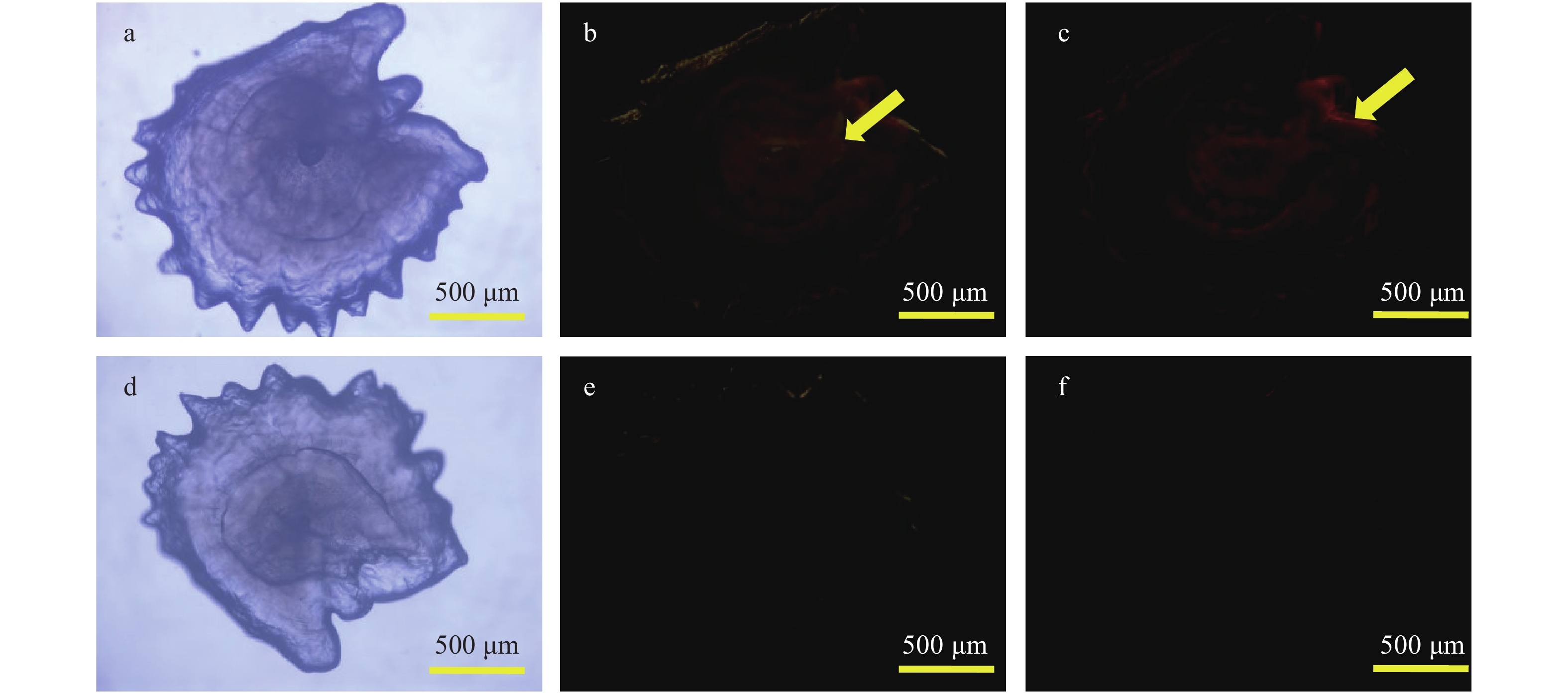
图 4 不同光源下中华倒刺鲃幼鱼星耳石的双重荧光标记效果
Figure 4. The effect of double fluorescent marking in asteriscus of S. sinensis larval observed with different light sources
![]()
图 5 不同光源下中华倒刺鲃幼鱼倒刺的双重荧光标记效果
Figure 5. The effect of double fluorescent marking in barb of S. sinensis larval observed with different light sources
![]()
图 6 不同光源下中华倒刺鲃幼鱼背鳍的双重荧光标记效果
Figure 6. The effect of double fluorescent marking in dorsal fin ray of S. sinensis larval observed with different light sources
![]()
图 7 不同光源下中华倒刺鲃幼鱼背棘的双重荧光标记效果
Figure 7. The effect of double fluorescent marking in dorsal fin spine of S. sinensis larval observed with different light sources
![]()
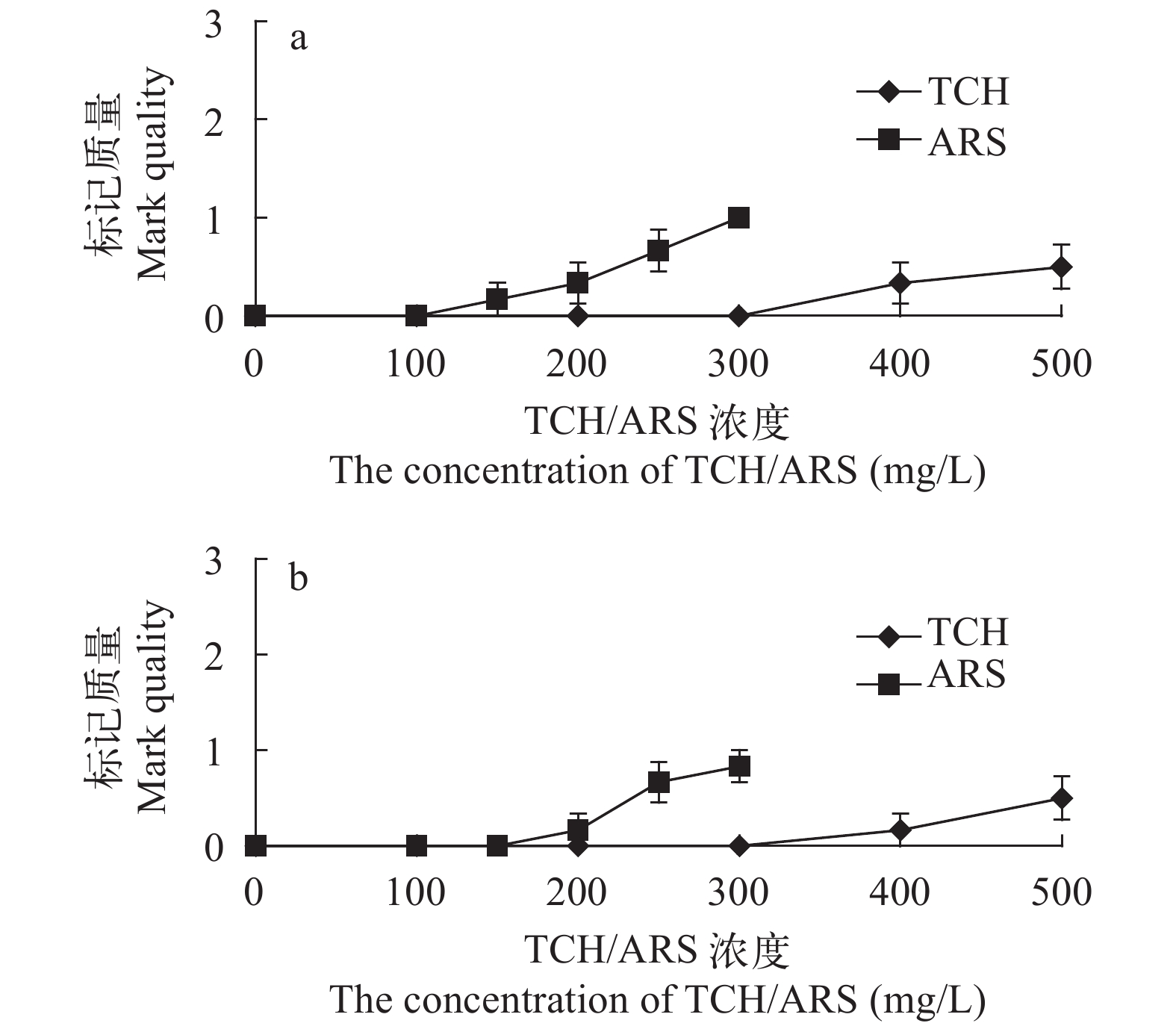
图 8 TCH和ARS荧光染料对中华倒刺鲃幼鱼侧线鳞(a)和非测线鳞(b)的标记质量
Figure 8. The mark quality of TCH and ARS fluorochrome dyes on the lateral line (a) and non-lateral line scales (b) of S. sinensis
![]()
图 9 TCH和ARS荧光染料对中华倒刺鲃幼鱼倒刺的标记质量
Figure 9. The mark quality of TCH and ARS fluorochrome dyes on the barbs of S. sinensis
![]()
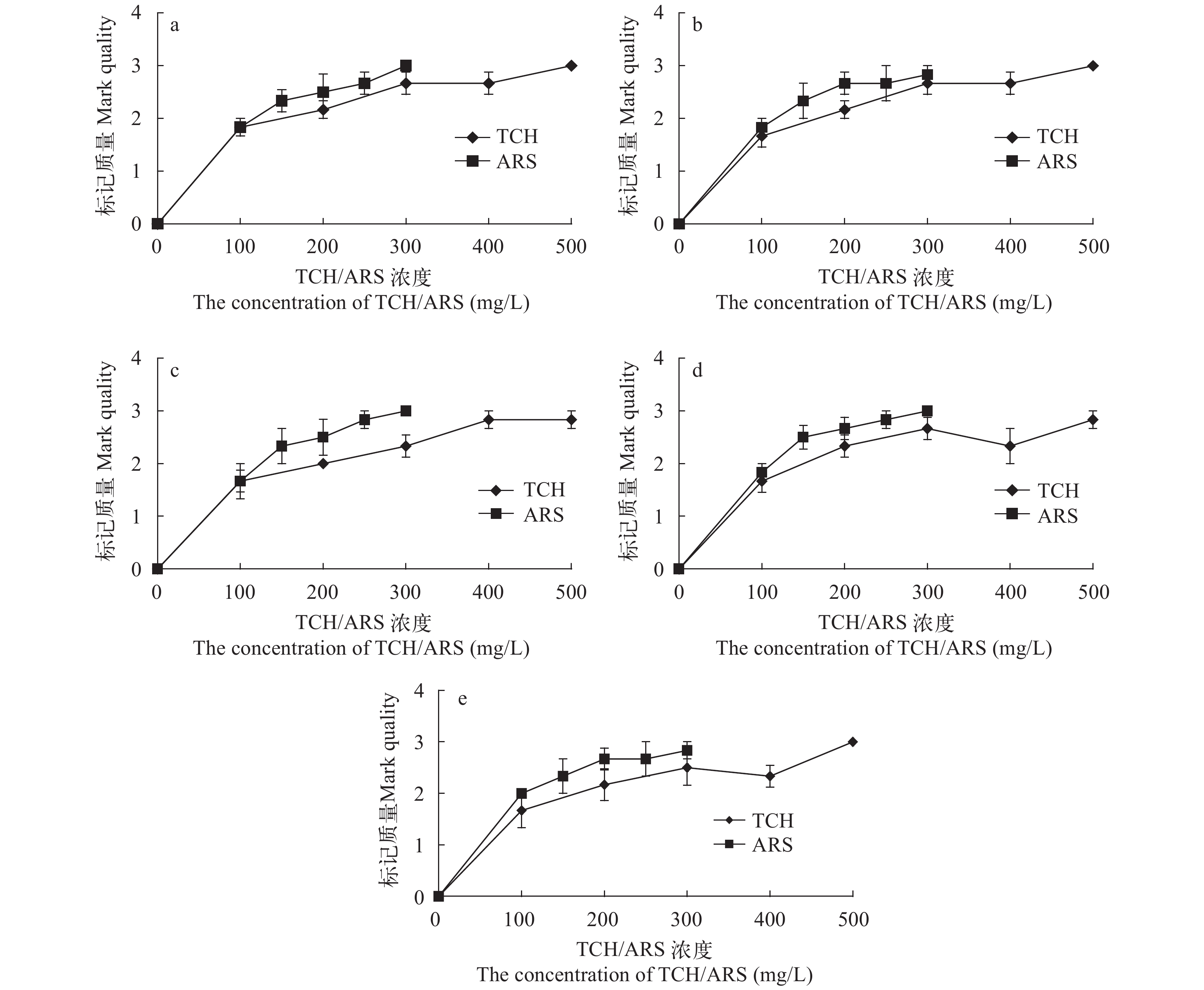
图 10 TCH和ARS荧光染料对中华倒刺鲃幼鱼背鳍(a)、胸鳍(b)、腹鳍(c)、臀鳍(d)和尾鳍(e)的标记质量
Figure 10. The mark quality of TCH and ARS fluorochrome dyes on the dorsal (a), pectoral (b), ventral (c), anal (d) and caudal (e) fin rays of S. sinensis
![]()
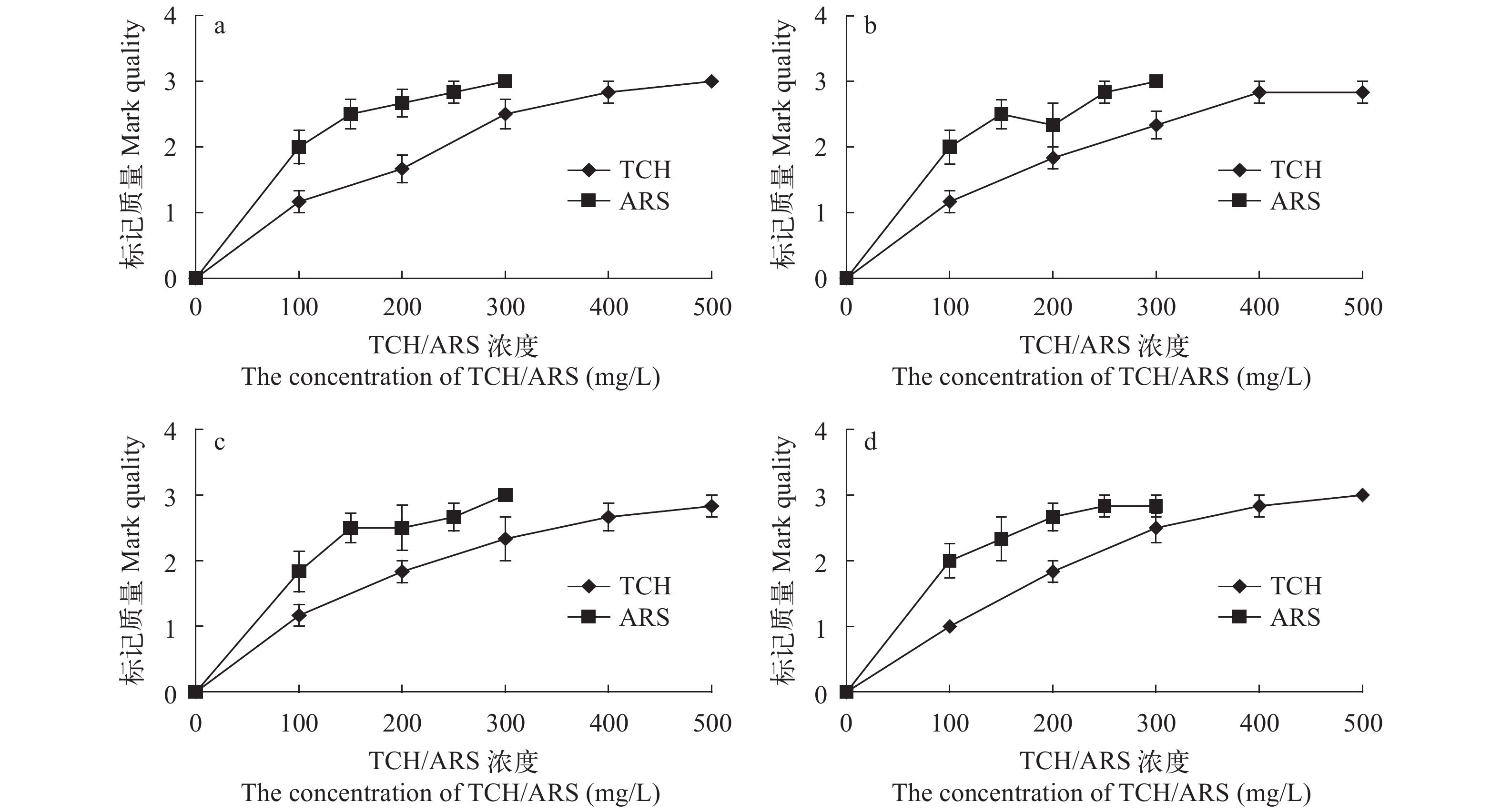
图 11 TCH和ARS荧光染料对中华倒刺鲃幼鱼背棘(a)、胸棘(b)、腹棘(c)和臀棘(d)的标记质量
Figure 11. The mark quality of TCH and ARS fluorochrome dyes on the dorsal (a), pectoral (b), ventral (c) and anal (d) fin spines fin rays of S. sinensis
表 1 使用盐酸四环素和茜素红S双重浸泡标记的顺序和浓度
Table 1 The order of double immersion marking and concentration sets for TCH and ARS of experimental group
浸泡标记顺序
Marking sequence荧光染料
Fluorochrome dye实验组
Experimental groupⅠ Ⅱ Ⅲ Ⅳ Ⅴ Ⅵ 第一次The first TCH (mg/L) 0 100 200 300 400 500 第二次The second ARS (mg/L) 0 100 150 200 250 300  下载: 导出CSV
下载: 导出CSV
表 2 用于检测盐酸四环素和茜素红S标记的双色镜和滤光片组合
Table 2 Dichromatic mirror and filter combination wavelength for visualizing TCH and ARS marks
光源
Light source波长Wavelength (nm) 激发滤光片
Excitation
filter双色镜
Dichromatic
mirror抑制滤光片
Suppression
filter蓝光Blue light 450—490 510 515 绿光Green light 515—560 580 590 注: 蓝光用于检测盐酸四环素(TCH)标记, 绿光用于检测茜素红S(ARS)标记Note: Blue wavelength was used to detect TCH marks and green wavelength was used to detect ARS marks
下载: 导出CSV
-
[1] 薛治泉. 浅析长江上游名特土著鱼的开发与保护 [J]. 中国水产, 2007(3): 76-77. doi: 10.3969/j.issn.1002-6681.2007.03.052 Xue Z Q. Analysis on the development and protection of the famous indigenous fish in the upper reaches of the Yangtze River [J]. China Fisheries, 2007(3): 76-77. [ doi: 10.3969/j.issn.1002-6681.2007.03.052
[2] 蔡焰值, 何长仁, 蔡烨强, 等. 中华倒刺鲃生物学初步研究 [J]. 淡水渔业, 2003(3): 16-18. doi: 10.3969/j.issn.1000-6907.2003.03.005 Cai Y Z, He C R, Cai Y Q, et al. The study on the biology of Spinibarbus sinensis [J]. Freshwater Fisheries, 2003(3): 16-18. [ doi: 10.3969/j.issn.1000-6907.2003.03.005
[3] 陈大庆. 长江渔业资源现状与增殖保护对策 [J]. 中国水产, 2003(3): 17-19. doi: 10.3969/j.issn.1002-6681.2003.03.013 Chen D Q. Current situation of fishery resources in the Yangtze River and counter measures for proliferation protection [J]. China Fisheries, 2003(3): 17-19. [ doi: 10.3969/j.issn.1002-6681.2003.03.013
[4] 高少波, 唐会元, 乔晔, 等. 金沙江下游干流鱼类资源现状研究 [J]. 水生态学杂志, 2013, 34(1): 44-49. Gao S B, Tang H Y, Qiao Y, et al. Status of fishery resources in the mainstream of the lower reaches of Jinsha River [J]. Journal of Hydroecology, 2013, 34(1): 44-49. [
[5] 朱忠胜, 宋娇文. 遵义市水产种质资源保护区建设与管理的思考 [J]. 中国水产, 2018(2): 62-64. Zhu Z S, Song J W. Thoughts on the construction and management of aquatic germplasm resources conservation area in Zunyi City [J]. China Fisheries, 2018(2): 62-64. [
[6] 赖鸿志. 索风营鱼类增殖放流站鱼类循环水养殖系统的研究与应用 [J]. 贵州水力发电, 2011, 25(1): 27-29. doi: 10.3969/j.issn.1007-0133.2011.01.008 Lai H Z. Research and application of fish recirculating aquaculture system in Suofengying fish breeding and discharging station [J]. Guizhou Water Power, 2011, 25(1): 27-29. [ doi: 10.3969/j.issn.1007-0133.2011.01.008
[7] 郭艳娜, 陈栋为, 赵再兴, 等. 乌江思林水电站库区鱼类增殖放流效果初步研究 [J]. 水生态学杂志, 2018, 39(1): 50-55. Guo Y N, Chen D W, Zhao Z X, et al. Evaluation of fishery resource enhancement by artificial propagation and release in the Silin Hydropower Station Reservoir [J]. Journal of Hydroecology, 2018, 39(1): 50-55. [
[8] 吴金明, 霍来江, 杜浩, 等. 茜素红S对中华倒刺鲃幼鱼不同组织抗氧化酶活性和脂质过氧化的影响 [J]. 水生生物学报, 2016, 40(6): 1172-1177. doi: 10.7541/2016.152 Wu J M, Huo L J, Du H, et al. Preliminary study of the alizarin red S effect on antioxidant enzymes activity and lipid peroxidation of Spinibarbus sinensis juveniles [J]. Acta Hydrobiologica Sinica, 2016, 40(6): 1172-1177. [ doi: 10.7541/2016.152
[9] 孙志禹, 张敏, 陈永柏. 水电开发背景下长江上游保护区珍稀特有鱼类保护实践 [J]. 淡水渔业, 2014, 44(6): 3-8. doi: 10.3969/j.issn.1000-6907.2014.06.001 Sun Z Y, Zhang M, ChenY B. Protection of the rare and endemic fish in the conservation area located in the upstream of the Yangtze River [J]. Freshwater Fisheries, 2014, 44(6): 3-8. [ doi: 10.3969/j.issn.1000-6907.2014.06.001
[10] 梅会清, 王波, 翟旭亮, 等. 重庆水生生物增殖放流工作思考 [J]. 中国水产, 2018(6): 56-59. Mei H Q, Wang B, Zhai X L, et al. Thoughts on the proliferation and release of aquatic organisms in Chongqing [J]. China Fisheries, 2018(6): 56-59. [
[11] 聂丽娜. 沱江四桥建设对长江上游国家级自然保护区鱼类资源的影响及保护措施 [J]. 环境影响评价, 2017, 39(5): 48-52. Nie L N. Impact of construction of Tuojiang Bridges on fish resources of National Nature Reserve in upper reaches of Yangtze River [J]. Environmental Impact Assessment, 2017, 39(5): 48-52. [
[12] Beckman D W, Schulz R G. A simple method for marking fish otoliths with alizarin compounds [J]. Transactions of the American Fisheries Society, 1996, 125(1): 146-149. doi: 10.1577/1548-8659(1996)125<0146:ASMFMF>2.3.CO;2
[13] Gelsleichter J, Cortes E, Manire C A, et al. Use of calcein as a fluorescent marker for elasmobranch vertebral cartilage [J]. Transactions of the American Fisheries Society, 1997, 126(5): 862-865.
[14] 洪波, 孙振中. 标志放流技术在渔业中的应用现状及发展前景 [J]. 水产科技情报, 2006, 33(2): 73-76. doi: 10.3969/j.issn.1001-1994.2006.02.003 Hong B, Sun Z Z. Application status and development prospect of marker releasing technology in fishery [J]. Fisheries Science & Technology Information, 2006, 33(2): 73-76. [ doi: 10.3969/j.issn.1001-1994.2006.02.003
[15] Barker J M, McKaye K R. Immersion marking of juvenile Midas cichlids with oxytetracycline [J]. North American Journal of Fisheries Management, 2004, 24(1): 262-269. doi: 10.1577/M02-144
[16] 霍来江. 茜素红S对中华倒刺鲃幼鱼的染色效果以及对其氧化应激响应 [D]. 武汉: 华中农业大学, 2015: 20-25. Huo L J. The dyeing effect of ARS on juvenile Spinibarbus sinenisis oxidative stressstress response to it [D]. Wuhan: Huazhong Agricultural University, 2015: 20-25.
[17] 徐永江, 柳学周, 史宝, 等. 茜素络合物对半滑舌鳎(Cynoglossus semilaevis)苗种耳石的染色标记效果 [J]. 渔业科学进展, 2016, 37(6): 11-18. doi: 10.11758/yykxjz.20151105002 Xu Y J, Liu X Z, Shi B, et al. Alizarin complexone marking of otoliths of juvenile Cynoglossus semilaevis [J]. Progress in Fishery Sciences, 2016, 37(6): 11-18. [ doi: 10.11758/yykxjz.20151105002
[18] 许莉佳. 荧光标记斑马鱼稚鱼耳石的效果及其对抗氧化酶活力的影响 [D]. 西南大学, 2012: 11-23. Xu L J. Marking juvenile Zebrafish with fluorescent substances and its effects on antioxidase [D]. Chongqing: Southwest University, 2012: 11-23.
[19] 吕红健. 许氏平鲉和褐牙鲆标志技术与标志放流追踪评价 [D]. 青岛: 中国海洋大学, 2013: 29-63. Lü H J. Experimental studies of marking techniques and mark-recapture for juvenile black rockfish (Sebastes schlegelii) and Japanese flounder (Paralichthys olivaceus) [D]. Qingdao: Ocean University of China, 2013: 29-63.
[20] Lü H J, Fu M, Xi D, et al. Experimental evaluation of using calcein and alizarin red S for immersion marking of bighead carp Aristichthys nobilis (Richardson, 1845) to assess growth and identification of marks in otoliths, scales and fin rays [J]. Journal of Applied Ichthyology, 2015, 31(4): 665-674. doi: 10.1111/jai.12778
[21] Crook D A, O’Mahony D, Gillanders B M, et al. Production of external fluorescent marks on Golden perch fingerlings through osmotic induction marking with alizarin red S [J]. North American Journal of Fisheries Management, 2007, 27(2): 670-675. doi: 10.1577/M06-053.1
[22] Honeyfield D C, Ostrowski C S, Fletcher J W, et al. Dietary calcein marking of brook trout, Atlantic salmon, yellow perch, and coho salmon scales [J]. North American Journal of Fisheries Management, 2006, 26(2): 431-437. doi: 10.1577/M05-107.1
[23] Monaghan J P. Comparison of calcein and tetracycline as chemical markers in summer flounder [J]. Transactions of the American Fisheries Society, 1993, 122(2): 298-301. doi: 10.1577/1548-8659(1993)122<0298:NCOCAT>2.3.CO;2
[24] Liu Q, Zhang X M, Zhang P D, et al. The use of alizarin red S and alizarin complexone for immersion marking Japanese flounder Paralichthys olivaceus (T.) [J]. Fisheries Research, 2009, 98(1): 67-74.
[25] 陈雷, 刘海映, 王秀利, 等. 荧光标记红鳍东方鲀可行性研究 [J]. 生物技术通报, 2014(2): 112-115. Chen L, Liu H Y, Wang X L, et al. Feasibility of fluorescent tag on Takifugu rubripe [J]. Biotechnology Bulletin, 2014(2): 112-115. [
[26] Tsukamoto K. Mass-marking of ayu eggs and larvae by tetracycline-tagging of otoliths [J]. Bulletin of the Japanese Society of Scientific Fisheries, 1985, 51(6): 903-911. doi: 10.2331/suisan.51.903
[27] Tsukamoto K. Otolith tagging of ayu embryo with fluorescent substances [J]. Nippon Suisan Gakkaishi, 1988, 54(8): 1289-1295. doi: 10.2331/suisan.54.1289
[28] 刘奇. 褐牙鲆标志技术与增殖放流试验研究 [D]. 青岛: 中国海洋大学, 2009: 20-26. Liu Q. Japanese flounder marking techniques and juveniles released for stock enhancemnt [D]. Qingdao: Ocean University of China, 2009: 20-26.
[29] 付自东, 李静, 岳碧松, 等. 用荧光物质浸泡标记胭脂鱼仔、稚鱼耳石 [J]. 动物学杂志, 2005(4): 60-65. doi: 10.3969/j.issn.0250-3263.2005.04.012 Fu Z D, Li J, Yue B S, et al. Otolith marking of larval and juvenile Chinese sucker with fluorescent substances [J]. Chinese Journal of Zoology, 2005(4): 60-65. [ doi: 10.3969/j.issn.0250-3263.2005.04.012
[30] 何春林, 付自东, 严太明, 等. 用荧光物质浸泡标记重口裂腹鱼仔鱼耳石 [J]. 四川动物, 2008(3): 331-334. He C L, Fu Z D, Yan T M, et al. Otolith marking of larval Schizothorax davidi with fluorescent substances [J]. Sichuan Journal of Zoology, 2008(3): 331-334. [
[31] Lü H J, Zhang X M, XI D, et al. Use of calcein and alizarin red S for immersion marking of black rockfish Sebastes schlegelii juveniles [J]. Chinese Journal of Oceanology and Limnology, 2014, 32(1): 88-98. doi: 10.1007/s00343-014-3022-9
[32] 刘岩, 孙典荣, 耿倩, 等. 茜素红与钙黄绿素标记不同规格黑鲷幼鱼的比较研究 [J]. 南方水产科学, 2016, 12(6): 17-24. doi: 10.3969/j.issn.2095-0780.2016.06.003 Liu Y, Sun D R, Geng Q, et al. Comparative study on immersion marking with alizarin red S and calcein for black porgy Acanthopagrus schlegelii of different sizes [J]. South China Fisheries Science, 2016, 12(6): 17-24. [ doi: 10.3969/j.issn.2095-0780.2016.06.003
[33] 许晓军, 杜建明, 张海琪, 等. 中华鳖稚鳖荧光标记技术研究 [J]. 宁波大学学报(理工版), 2012, 25(2): 13-16. Xu X J, Du J M, Zhang H Q, et al. Study on fluorescent tagging of juvenile soft-shelled turtle (Pelodiscus sinensis) [J]. Journal of Ningbo University (
Natural Science & Engineering Edition ) , 2012, 25(2): 13-16. [ [34] Bashey F. A comparison of the suitability of alizarin red S and calcein for inducing a nonlethally detectable mark in juvenile guppies [J]. Transactions of the American Fisheries Society, 2004, 133(6): 1516-1523. doi: 10.1577/T03-073.1
[35] Reinert T R, Wallin J, Griffin M C, et al. Long-term retention and detection of oxytetracycline marks applied to hatchery-reared larval striped bass, Morone saxatilis [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1998, 55(3): 539-543. doi: 10.1139/f97-280
[36] Bumguardner B W, King T L. Toxicity of oxytetracycline and calcein to juvenile striped bass [J]. Transactions of the American Fisheries Society, 1996, 125(1): 143-145. doi: 10.1577/1548-8659(1996)125<0143:TOOACT>2.3.CO;2
[37] Meyer S, Sorensen S R, Peck M A, et al. Sublethal effects of alizarin complexone marking on Baltic cod (Gadus morhua) eggs and larvae [J]. Aquaculture, 2012(324-325): 158-164.
-
期刊类型引用(3)
1. 李毅,杨婉玲,王超,赖子尼,李海燕,刘乾甫,高原,麦永湛,曾艳艺. 珠江河网密刺苦草种群衰退的表观性状特征指示. 水生态学杂志. 2024(04): 155-164 .  百度学术
百度学术
2. 张家硕,赵一帆,许盛,何培民,邵留. 芸苔素内酯浸泡处理对苦草生长的影响. 环境科学与技术. 2024(07): 1-7 . 百度学术
3. 涂茜,冯志,朱海涛,章北平,谢承培. 种植密度对4种沉水植物净化富营养化水体效果的影响. 环境保护科学. 2022(06): 102-109 . 百度学术
其他类型引用(6)









计量
- 文章访问数: 3646
- HTML全文浏览量: 1081
- PDF下载量: 94
- 被引次数: 9