
GROWTH, MORTAILY AND OPTIMUM CATCHABLE SIZE OF COILIA MYSTUS IN OUJIANG RIVER ESTUARY
-
摘要: 根据2015、2018—2020年瓯江口水域渔业资源监测调查资料, 对1359尾凤鲚的体长、全长和体重等生物学信息进行测定, 利用体长频率分布估算凤鲚种群生长、死亡参数等。结果表明, 瓯江口凤鲚体长为5.2—21.9 cm; 体长和体重的关系式W=0.0035×L3.0783。用ELEFAN技术拟合的von Bertalanffy生长方程的各参数为L∞=26.60 cm, k=0.47及t0=–0.55 a; Z=2.30, M=1.00, F=1.30, E=0.567, 表明资源处于过度开发状态; 凤鲚的首次捕捞体长Lc=15.35 cm, 对应首次捕捞平均年龄为1.28 a, 小于临界年龄(1.36 a)和体重生长的拐点年龄(1.85 a), 表明当前渔业主要捕捞对象为幼鱼和补充群体, 无法保证资源的有效补充。在瓯江口凤鲚目前的资源状态下, 应努力降低捕捞死亡率, 并加强对凤鲚栖息环境的保护, 而对目前以小型化和低龄化为主的凤鲚群体, 以控制开捕体长为主, 根据Beverton-Holt动态模型, 建议开捕体长为15.50 cm, 对应开捕年龄为1.31 a。Abstract: Coilia mystus is one of the main fishing species in Oujiang River Estuary and it has high economic value. Based on the fishery resources monitoring data collected in Oujiang River Estuary in Wenzhou from 2015 and 2018 to 2020, the present study analyzed the biological characteristics (n=1359) of C. mystus, including body length, total length and body weight. The population growth and death parameters were estimated by body length frequency distribution. The results indicated that the average body length of C. mystus in Oujiang River Estuary was (15.1±3.0) cm, with the group of predominant body length 14.0—18.0 cm, accounting for 54.3%; the average body weight was (16.8±10.1) g, with the group of predominant body weight 5.0—25.0 g, accounting for 72.0%. The distribution of body length and body weight differed significantly between males and females (P<0.05). The average body length and body weight of females were greater than those of males, but the body length-weight relationship was not significantly different (P>0.05). The relationship between body length and weight was expressed as: W=0.0035×L3.0783 (R2=0.9509). The power exponent b was 3.0783, which is consistent with uniform growth characteristics. The growth parameters of von Bertalanffy formula estimated by ELEFAN were L∞=26.60 cm, k=0.47, and t0=−0.55 a. The total mortality coefficient (Z) was calculated using a length-converted catch curve as 2.30. The natural mortality coefficient (M) estimated referring to the Pauly empirical formula, was 1.00. Therefore, fishing mortality coefficient (F) was 1.30, and the exploitation rate (E) equaled 0.567, demonstrating that the stock had been over-exploited. The catch curve also determined that the current first capture L50 was 14.50 cm, corresponding to an age of 1.13, which was lower than both the critical age (1.36 a) and the turning point age (1.85 a). This indicated that the main fishing objectives were juveniles and included the recruitment group of C. mystus. Under the current resource status in Oujiang River Estuary, great amount of effort should be made to reduce fishing mortality and protect C. mystus habitat. It was suggested that the control of catchable length should be the main management strategy for the trend of miniaturization and low age of C. mystus resources. According to the Beverton-Holt dynamic model, the minimum capture size for C. mystus should be 15.50 cm (age 1.31 years).
-
Keywords:
- Biological parameter /
- Optimum catchable size /
- Oujiang River Estuary /
- Coilia mystus
-
凤鲚(Coilia mystus), 俗称凤尾鱼、烤子鱼、籽鲚等, 隶属于鲱形目(Clupeiformes), 鳀科(Engraulidae), 鲚属(Coilia), 主要分布于我国黄、渤海和东海近海海域及长江、珠江、闽江和瓯江等江河口[1, 2]。袁传宓等[3]从形态特征角度分析将我国凤鲚分为长江型、闽江型和珠江型3个生态类群, 阎雪岚等[4]从遗传结构进行分析, 认为至少存在长江型和闽江型2个亚种。瓯江位于浙江省南部, 是浙江境内第二大河流, 因其径流量较大, 为河口区带来较丰富的营养物质。瓯江口海域作为东海近海第二大渔场——温台渔场的重要组成部分, 渔业资源丰富, 为许多鱼类索饵、产卵和繁育提供了场所[5-7]。凤鲚具有较高的经济价值, 是瓯江口主要捕捞对象之一, 20世纪50年代年产量达到500—600 t[5]。仲伟等[8]认为瓯江凤鲚大多为1龄, 产卵场可能位于江心屿一带, 汛期个体比长江口、舟山近海凤鲚大。近年来, 由于过度捕捞、水域环境污染及河道采砂作业等人类活动破坏了凤鲚产卵、索饵和洄游通道, 使其出现个体小型化、资源量下降严重等问题, 导致当前的凤鲚种群不稳定[1, 9, 10], 急需加强瓯江渔业保护, 制定有效的管理措施以维持凤鲚资源的可持续利用。
生长和死亡作为鱼类的重要生物学属性, 是影响鱼类资源补充潜力与稳定性的重要因素[11]。鱼类年龄和生长的研究可作为预测资源量变动的主要根据, 也可以为渔业资源可持续利用提供重要参考[12, 13]。死亡参数为渔业资源评估和探索渔业资源变动规律提供基础资料[12]。目前, 我国学者关于凤鲚的研究主要集中于基础生物学[10, 14-16]和遗传多样性[17], 且主要研究区域为长江口及邻近海域, 有关瓯江口凤鲚的相关报道较少[3, 16]; 同时, 有关瓯江口凤鲚的生长、死亡和资源状况研究仍缺乏。因此本研究以瓯江口凤鲚为研究对象, 测定其体长、体重和性腺成熟度等生物学指标, 运用FiSAT Ⅱ 软件中的体长频率变换法估算生长参数和死亡参数, 运用Beverton-Holt动态综合模型评价种群资源的利用现状, 并提出最适开捕规格, 旨在了解瓯江口凤鲚的生长特性并丰富其基础研究资料, 为凤鲚资源的科学管理和合理利用提供参考。
1. 材料与方法
1.1 数据来源
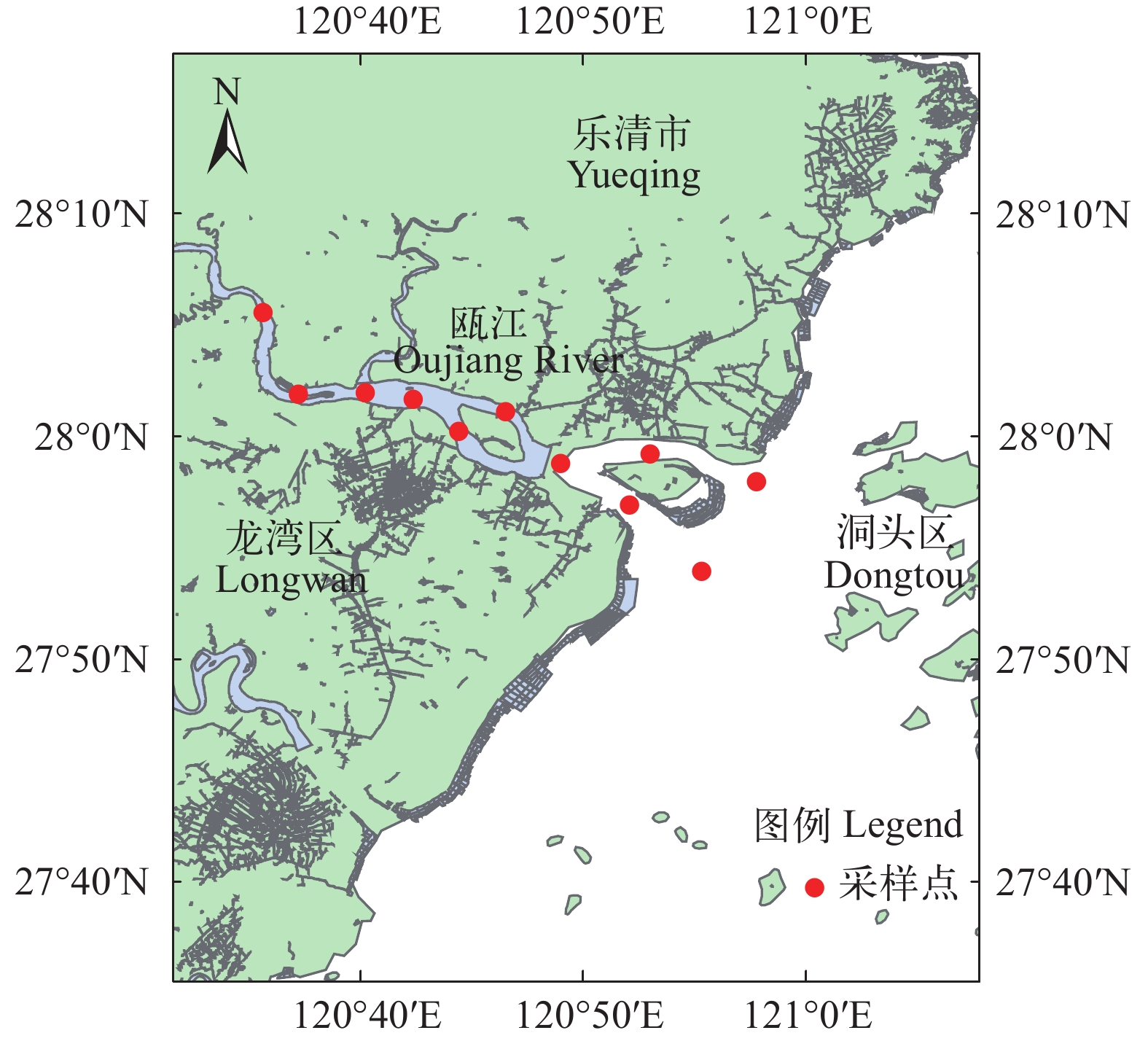
2015、2018—2020年4—8月于瓯江上游至瓯江口水域(27°53′ N—28°5′ N, 120°35′ E—120°55′ E)流刺网(网长60 m, 网高4—8 m, 网目尺寸为5.0 cm)渔获物中取样, 共设置11个采样地点(图 1)。将采集到的凤鲚样品带回实验室进行生物学测定, 测定内容包括体长、全长、体重和性腺成熟度等, 其中体长精确到0.1 cm, 体重精确到0.1 g。各取样地点凤鲚尾数不足50尾时全部测定, 否则随机测定50尾, 研究共测定凤鲚1359尾, 体长数据按月份、组距1.0 cm进行整理, 形成体长频率时间序列。
1.2 分析方法
体长与体重关系 将尾数占样品总尾数10%以上的体长组和体重组定义为优势体长组和优势体重组[18]。体长和体重关系采用幂函数关系进行拟合[19], 表达式为:
$ W = a × L^{b} $
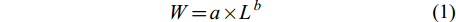
(1) 式中, W表示体重(g); L表示体长(cm); a为生长的条件因子; b为生长指数, b=3为等速生长, b≠3则为异速生长[20]。
生长、死亡参数估算 将按年、月整理的凤鲚体长频率时间序列数据导入FiSAT Ⅱ软件, 用ELEFAN I(Electronic length frequency analysis I)技术估算渐进体长和生长速率等生长参数, 取拟合优度(Score)最大且从生物学角度考虑能被接受的数值(L∞和K)作为生长参数的估计值[21, 22]。采用von Bertalanffy生长方程[23]拟合凤鲚的生长:
$ L_{t} = L_{∞} × [1 - {\rm{e}}^{-k×(t-t_0)}] $
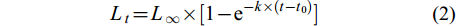
(2) 采用Pauly经验公式[24]计算理论生长初始年龄t0:
$ {\rm{ln}}(-t_{0}) = -0.3922 - 0.2752{\rm{ln}}TL_{∞} - 1.038{\rm{ln}}k$
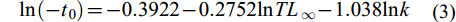
(3) 总死亡系数(Z)采用体长变换渔获曲线法估算, 自然死亡系数(M)采用Pauly经验公式[24]估算:
$ \begin{split} {\rm{ln}}M =& -0.0066 - 0.279 × {\rm{ln}}L_{∞} + 0.6543×{\rm{ln}}k +\\& 0.4634 × {\rm{ln}}T \end{split} $
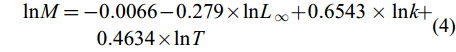
(4) 捕捞死亡系数(F)和开发率(E)由总死亡系数(Z)、自然死亡系数(M)根据公式(5)、(6)计算得出:
$ F = Z - M $
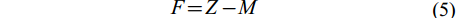
(5) $ E = F/Z\;\; $
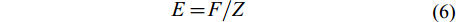
(6) 式中, Lt和L∞分别表示凤鲚t龄时的体长和渐近体长; k表示生长速率; t0表示理论生长初始年龄。TL∞为渐近全长(cm), 由渐近体长L∞根据体长-全长关系转换获得。故选取200尾凤鲚的体长(L)和全长(TL)数据拟合体长-全长关系式: TL=1.0644×L+9.2291(R2=0.9646), 将渐进体长带入关系式求出凤鲚的渐近全长。T为凤鲚栖息环境的平均温度(℃), 根据调查站位测得凤鲚在河口产卵场栖息的平均水温为25.90℃。
临界年龄和拐点年龄 在没有捕捞的状态下, 一个世代的资源量达到最大值时对应的年龄为临界年龄Tc, 计算公式[25]为:
$ T_{{\rm{c}}} = [kt_{0} - {\rm{ln}}M + {\rm{ln}}(bk + M)]/k $
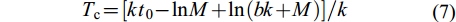
(7) 体重达到最大或体重生长加速度为0时对应的年龄为拐点年龄tp, 其计算公式[12]为:
$ {\rm{t}}_{{\rm{p}}} = {\rm{ln}}b/k + t_{0} $
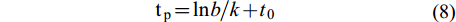
(8) 式中, b为体长-体重关系中的生长指数。上述计算过程在Excel中完成。
资源动态 根据体长变换曲线所拟合的直线方程, 推算线性回归中未被使用的各点相对应的期望值ln(N/dt), 计算各点的期望值和观测值之比, 以及这些点的比率的累积率, 当累积率达50%的点所对应的体长作为平均选择体长的估计值(L50), 即为开捕体长[26]。
在FiSAT Ⅱ软件中, 采用Beverton-Holt模型中的刀刃式选择假设模型建立相对单位补充量渔获量(Y'/R)和相对单位补充资源量(B'/R)模型[27], 根据Y'/R和B'/R二维曲线评价瓯江口凤鲚资源的利用状况。其数学表达式为:
$ \begin{split} Y'/R = & EU^{M/k} × [1 - 3U/(1 + m) +\\&3U^{2}/(1 + 2m)-U^{3}/(1 + 3m)] \end{split}$
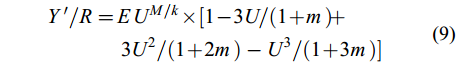
(9) $ B'/R = (Y'/R)/F \qquad \qquad \qquad\qquad \; $

(10) $ U = 1 - (L_{c}/L_{∞})\qquad \qquad \qquad\qquad \quad $

(11) $ m = (1 - E)/(M/k) = k/Z \qquad \qquad \quad $
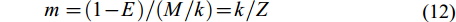
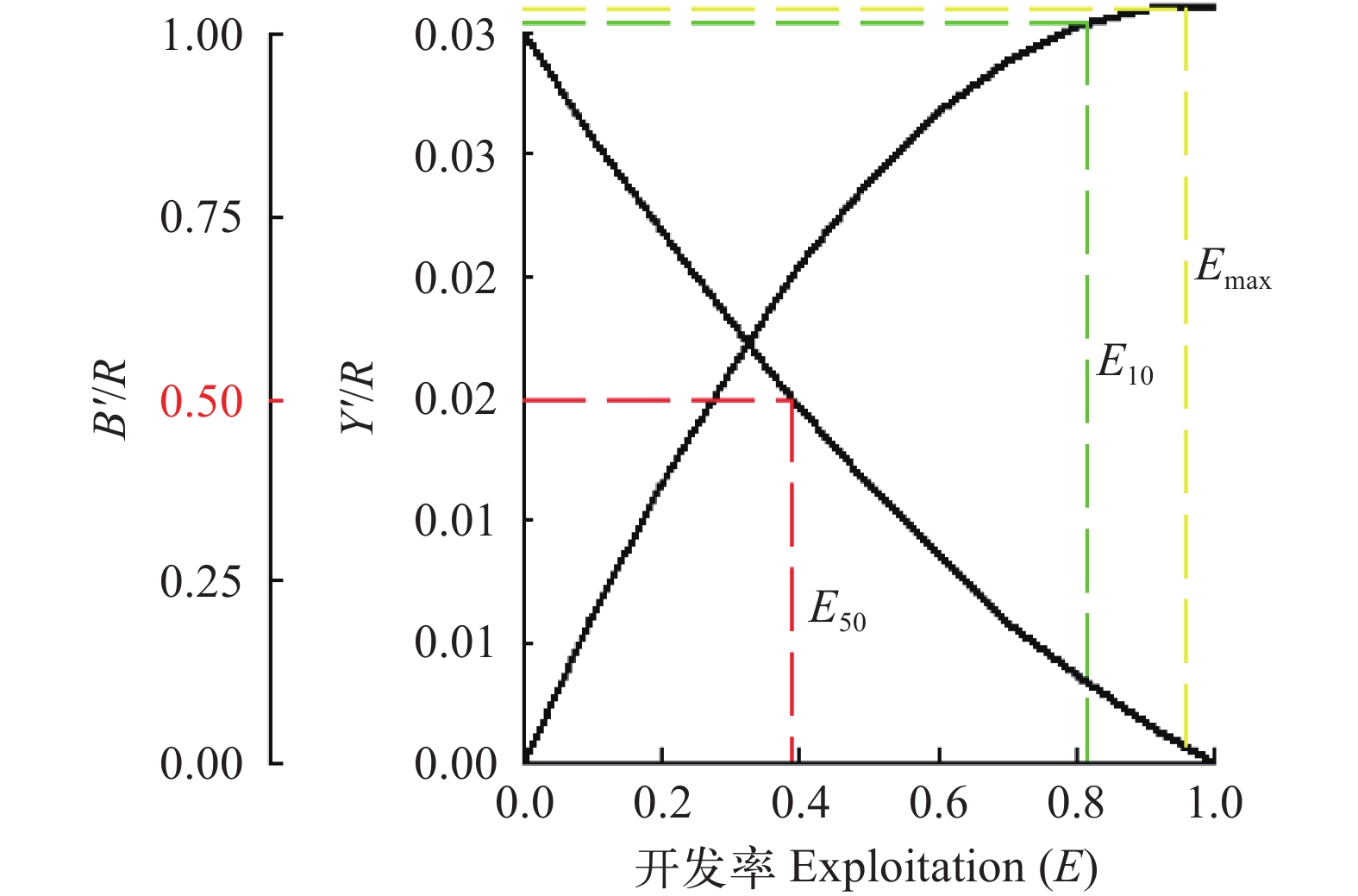
(12) 式中, E为资源开发率, 包括3种水平: Emax为产量最高时的开发率, E10为Y'/R边际增长减少10%的开发率, E50为资源量减少为原始水平50%时的开发率[28]。
2. 结果
2.1 体长与体重的关系
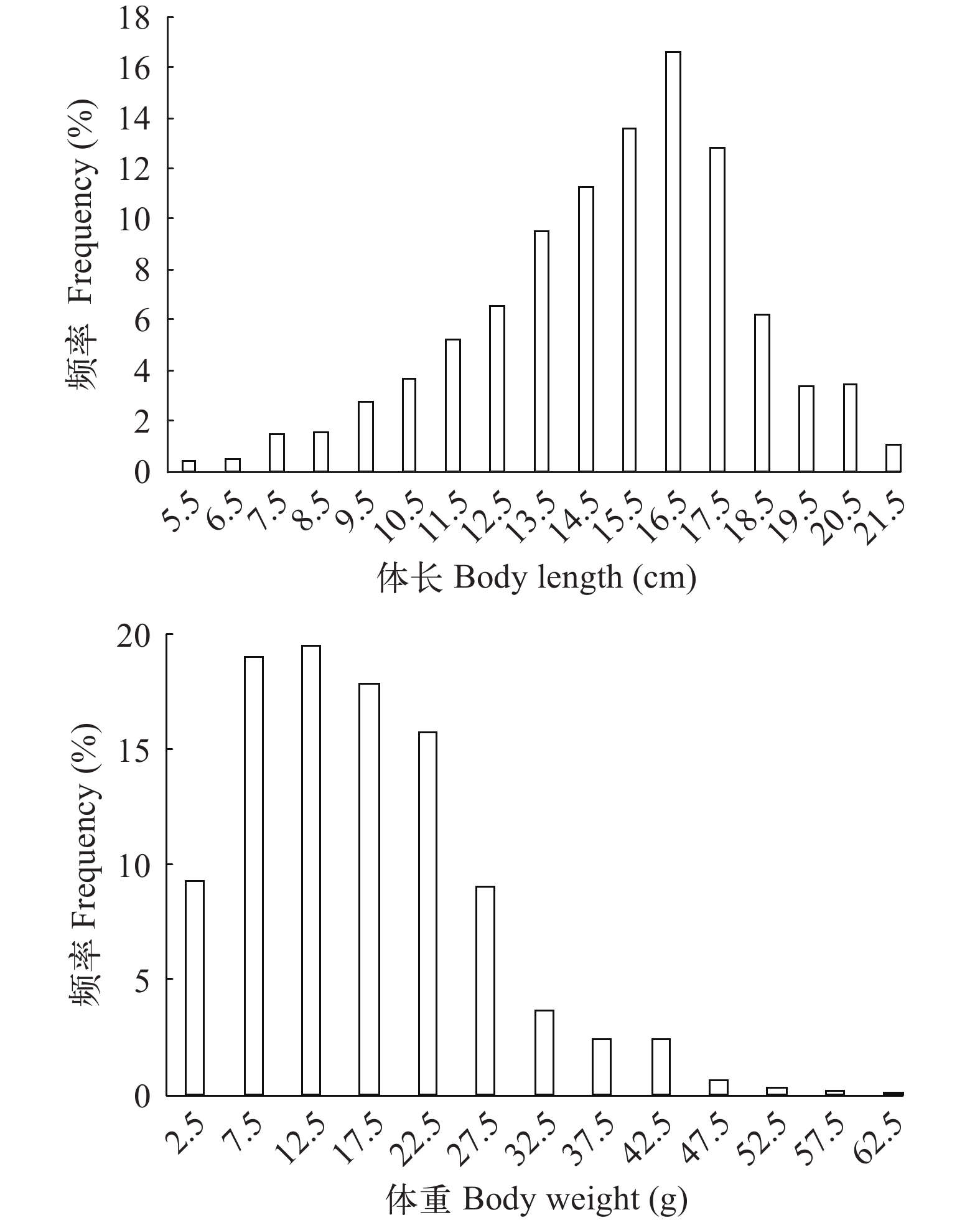
体长、体重分布 共测定样品1359尾, 凤鲚体长为5.2—21.9 cm, 平均值为(15.1±3.0) cm, 优势体长组为14.0—18.0 cm(54.3%, 图 2); 体重为0.3—63.0 g, 平均值为(16.8±10.1) g, 优势体重组为5.0—25.0 g(72.0%, 图 2)。各年份的体长和体重情况见表 1。
![]() 图 2 瓯江口凤鲚体长和体重分布Figure 2. Body length and body weight distribution of Coilia mystus in Oujiang River Estuary表 1 不同年份凤鲚的体长、体重分布Table 1. Body length and body weight distribution in different year groups of Coilia mystus
图 2 瓯江口凤鲚体长和体重分布Figure 2. Body length and body weight distribution of Coilia mystus in Oujiang River Estuary表 1 不同年份凤鲚的体长、体重分布Table 1. Body length and body weight distribution in different year groups of Coilia mystus年份
Year样本数
No. of
samples体长Body length (cm) 体重Body weight (g) 均值±
标准差
Mean±SD范围
Range均值±
标准差
Mean±SD范围
Range2015 429 16.1±2.1 8.2—21.9 19.5±9.2 2.2—53.6 2018 571 13.4±3.2 5.2—21.7 11.9±8.8 0.3—56.7 2019 121 15.7±2.8 6.8—21.9 17.6±10.0 0.9—44.0 2020 238 17.0±1.7 12.7—21.8 23.7±8.5 8.3—63.0 共测定雌性个体362尾, 体长为12.6—21.9 cm, 平均值为(17.4±1.9) cm, 优势体长组为15.0—19.0 cm(66.9%); 体重为6.6—63.0 g, 平均值为(26.3±10.1) g, 优势体重组为15.0—30.0 g(60.8%)。雄性个体584尾, 体长为9.8—21.3 cm, 平均值为(15.6±1.8) cm, 优势体长组为13.0—18.0 cm(83.0%), 体重为4.3—42.5 g, 平均值为(16.51±6.3) g, 优势体重组为5.0—25.0 g(90.2%)。雌雄个体间体长、体重分布差异显著(Mann-Whitney U, P<0.05), 雌性个体平均体长和体重均大于雄性。
体长-体重关系 凤鲚体长与体重呈幂函数增长关系, 可用Wt=a×Lb来表示。不同年份凤鲚的体长-体重关系式为:
$ \begin{split} 2015年 : & W_{t} = 0.0020 × L_{t}^{3.2823}\\&(R^{2} = 0.8897,n = 429) \end{split} $
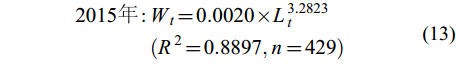
(13) $ \begin{split} 2018年 : &W_{t} = 0.0051 × L_{t}^{2.9243}\\&(R^{2} = 0.9572,n = 571) \end{split} $
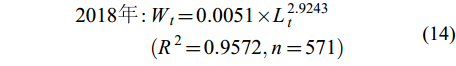
(14) $ \begin{split} 2019年 : & W_{t} = 0.0020 × L_{t}^{3.2460}\\&(R^{2} = 0.9640,n = 121) \end{split} $
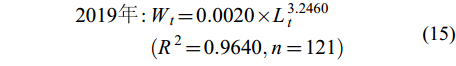
(15) $ \begin{split} 2020年 : & W_{t} = 0.0012 × L_{t}^{3.4738}\\&(R^{2} = 0.8708,n = 238) \end{split} $
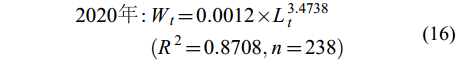
(16) $ \begin{split} 雌\; 性 : &W_{t} = 0.0016 × L_{t}^{3.3711}\\&(R^{2} = 0.9030,n = 362) \end{split} $
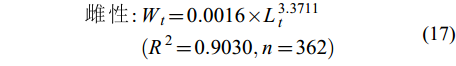
(17) $ \begin{split} 雄\; 性 : & W_{t} = 0.0030 × L_{t}^{3.1084}\\&(R^{2} = 0.8893,n = 584) \end{split}$
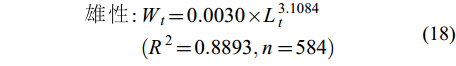
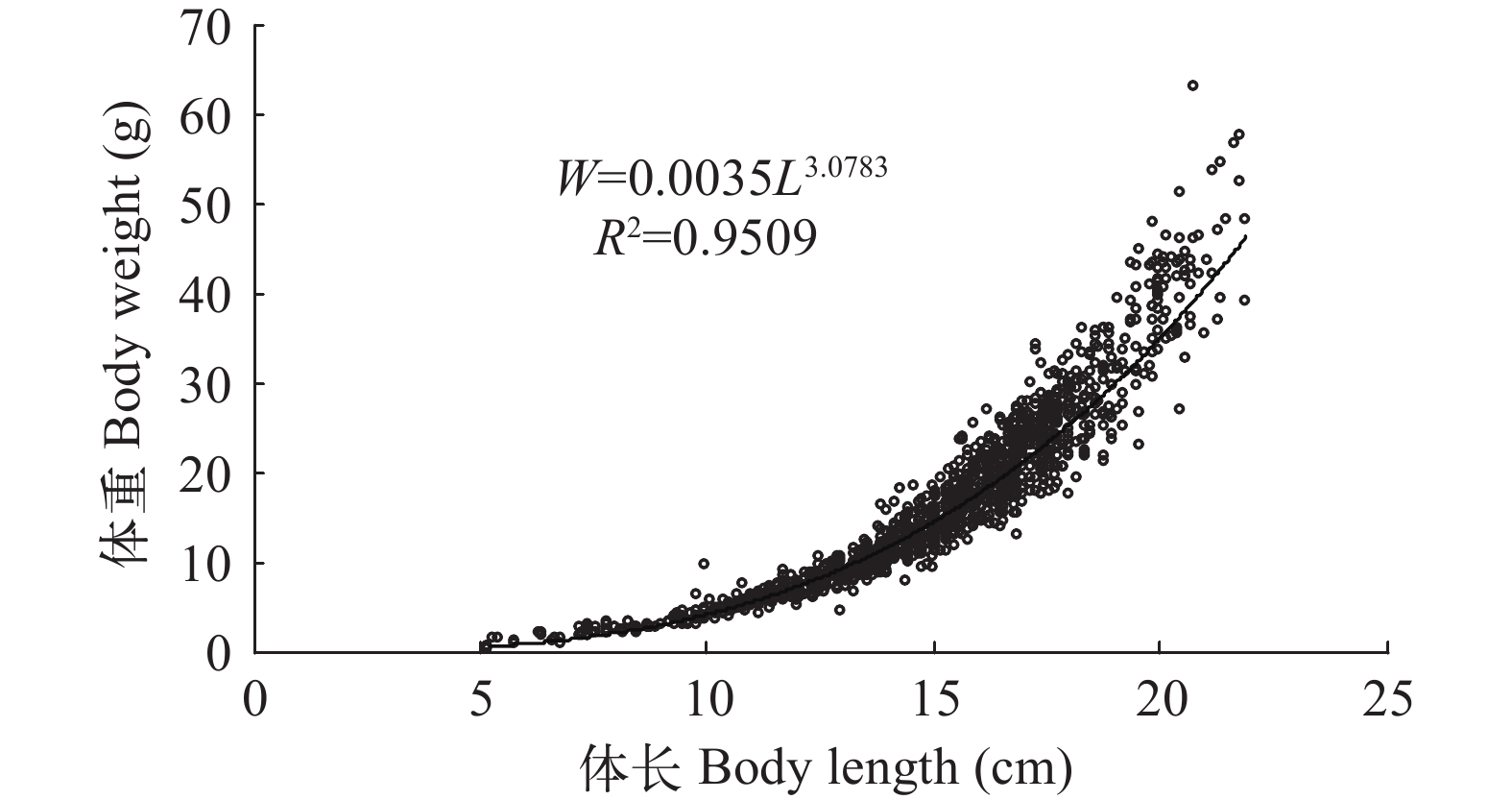
(18) 利用SPSS软件中的单因素方差分析(One-Way ANOVA)检验方程(13)、(14)、(15)、(16)之间的差异显著性, 其中选取5.0 cm作为体长起点, 最大体长为22.0 cm, 组间间隔1.0 cm, 共18组。体重是依据对应体长-体重关系式分别计算的理论值。结果表明, 不同年份凤鲚的体长和体重关系无显著性差异(P>0.05)。用同样的方法检验凤鲚雌、雄性个体的体长-体重关系, 得P>0.05, 即雌、雄性个体的生长没有显著性差异。为此, 本文将瓯江口不同年份、不同性别的凤鲚测定数据合一开展研究, 凤鲚体长-体重的关系式为W=0.0035×L3.0783(R2=0.9509; 图 3)。经t检验, 本研究中的b与3差异不显著(P>0.05), 因此, 凤鲚呈匀速生长。
![]() 图 3 瓯江口凤鲚体长和体重的关系Figure 3. The body length-weight relationship of Coilia mystus in Oujiang River Estuary
图 3 瓯江口凤鲚体长和体重的关系Figure 3. The body length-weight relationship of Coilia mystus in Oujiang River Estuary2.2 年龄和生长
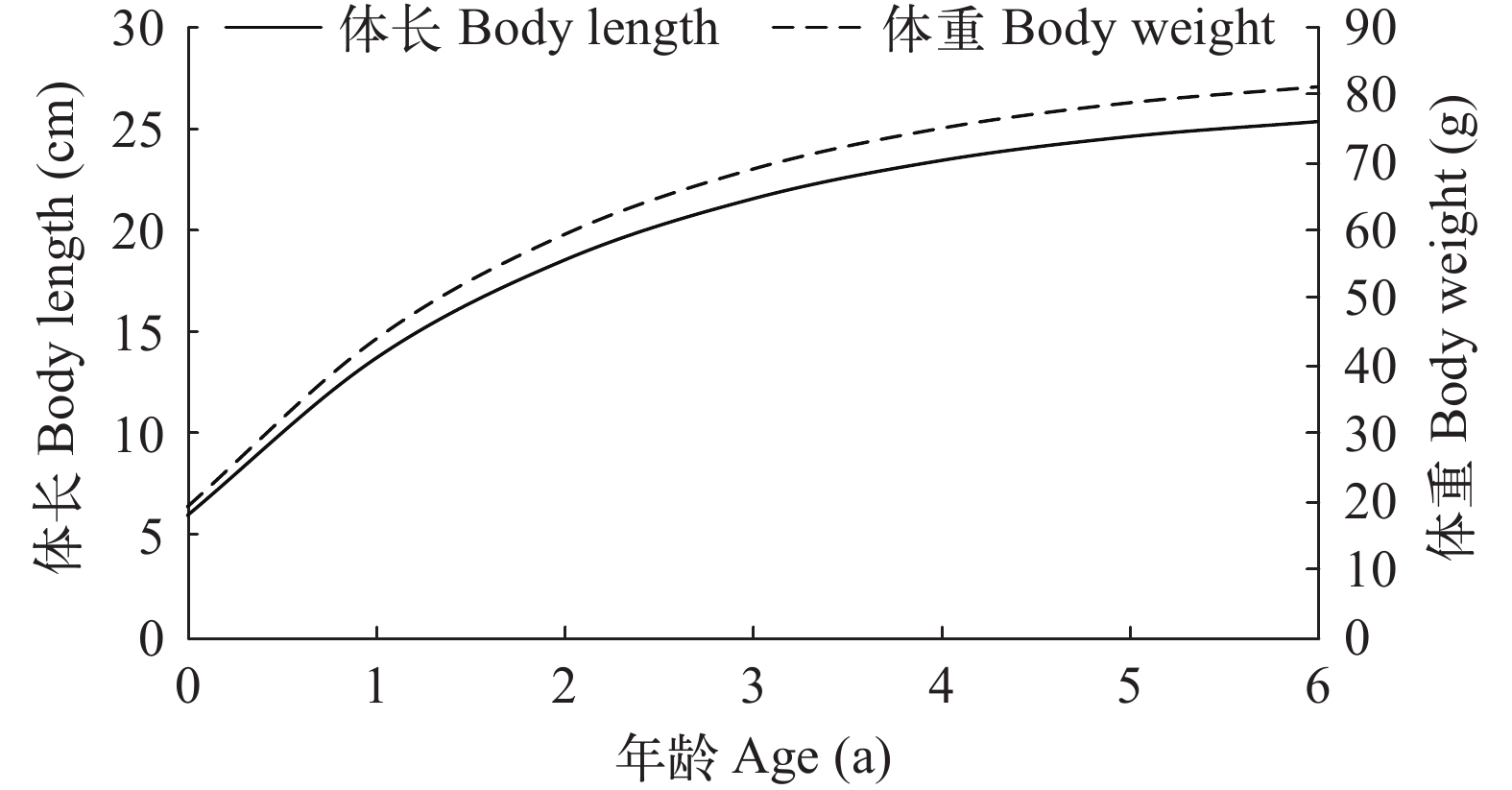
生长参数和生长方程 经ELEFAN Ⅰ方法估算求得渐近体长L∞为26.60 cm, 生长速率k=0.47, 理论生长初始年龄t0=–0.55 a, 拟合的体长生长方程为Lt=26.60×[1–e–0.47(t+0.55)]; 根据体长-体重关系式求得渐进体重W∞为85.17 g, 体重生长方程为Wt=85.17×[1–e–0.47(t+0.55)]3.0783。根据生长方程绘制体长和体重生长曲线(图 4)。
![]() 图 4 瓯江口凤鲚的体长和体重生长曲线Figure 4. The growth curve in body length and body weight of Coilia mystus in Oujiang River Estuary
图 4 瓯江口凤鲚的体长和体重生长曲线Figure 4. The growth curve in body length and body weight of Coilia mystus in Oujiang River Estuary年龄 根据拟合的体长生长方程Lt=26.60×[1–e–0.47(t+0.55)]计算年龄t, t≤0.5记为0龄, 0.5<t≤1.5记为1龄, 1.5<t≤2.5记为2龄, 依次类推。结果表明, 1359尾凤鲚样本包括0—3龄4个年龄组。其中, 0龄组106尾, 占全部样本的7.8%; 1龄组755尾, 占全部样本的55.6%; 2龄组458尾, 占全部样本的33.7%; 3龄组40尾, 占全部样本的2.9%。1龄组和2龄组是优势群体, 共占89.3%。表 2为各年龄组的体长和体重情况。
表 2 不同年龄组凤鲚的体长、体重分布Table 2. Body length and body weight distribution in different age groups of Coilia mystus年龄组
Age样本数
No. of
samples体长Body length (cm) 体重Body weight (g) 均值±
标准差
Mean±SD范围
Range均值±
标准差
Mean±SD范围
Range0 106 8.6±1.3 5.2—10.3 3.0±1.2 5.2—9.6 1 755 14.1±1.6 10.4—16.4 12.1±4.8 3.9—27.0 2 458 17.8±1.0 16.5—20.2 25.5±6.4 13.0—47.8 3 40 20.9±0.5 20.3—21.9 43.3±7.4 27.0—63.0 生长速度和生长加速度 凤鲚生长过程的总和可由生长曲线反映, 而生长过程中具体的特征变化则由生长速度和生长加速度来描述。将体长、体重生长方程对年龄t求导, 得到体长、体重的生长速度和生长加速度方程分别为:
dL/dt=12.5×[1–e–0.47(t+0.55)]
dW/dt=83.2e–0.47(t+0.55)[1–e–0.47(t+0.55)]2.0783
d2W/d2t=39.1e–0.47(t+0.55)[1–e–0.47(t+0.55)]× [3.0783e–0.47(t+0.55)–1]
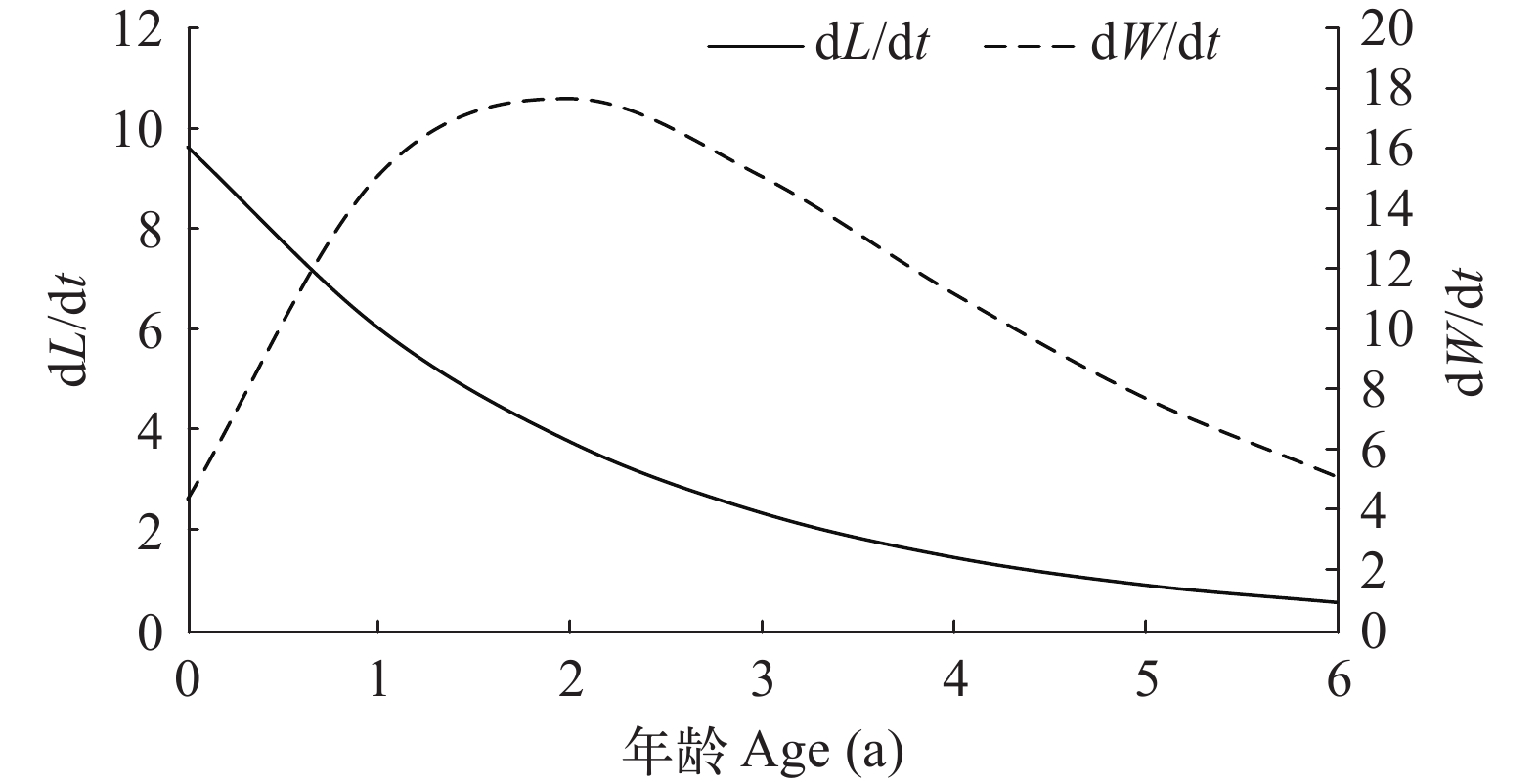
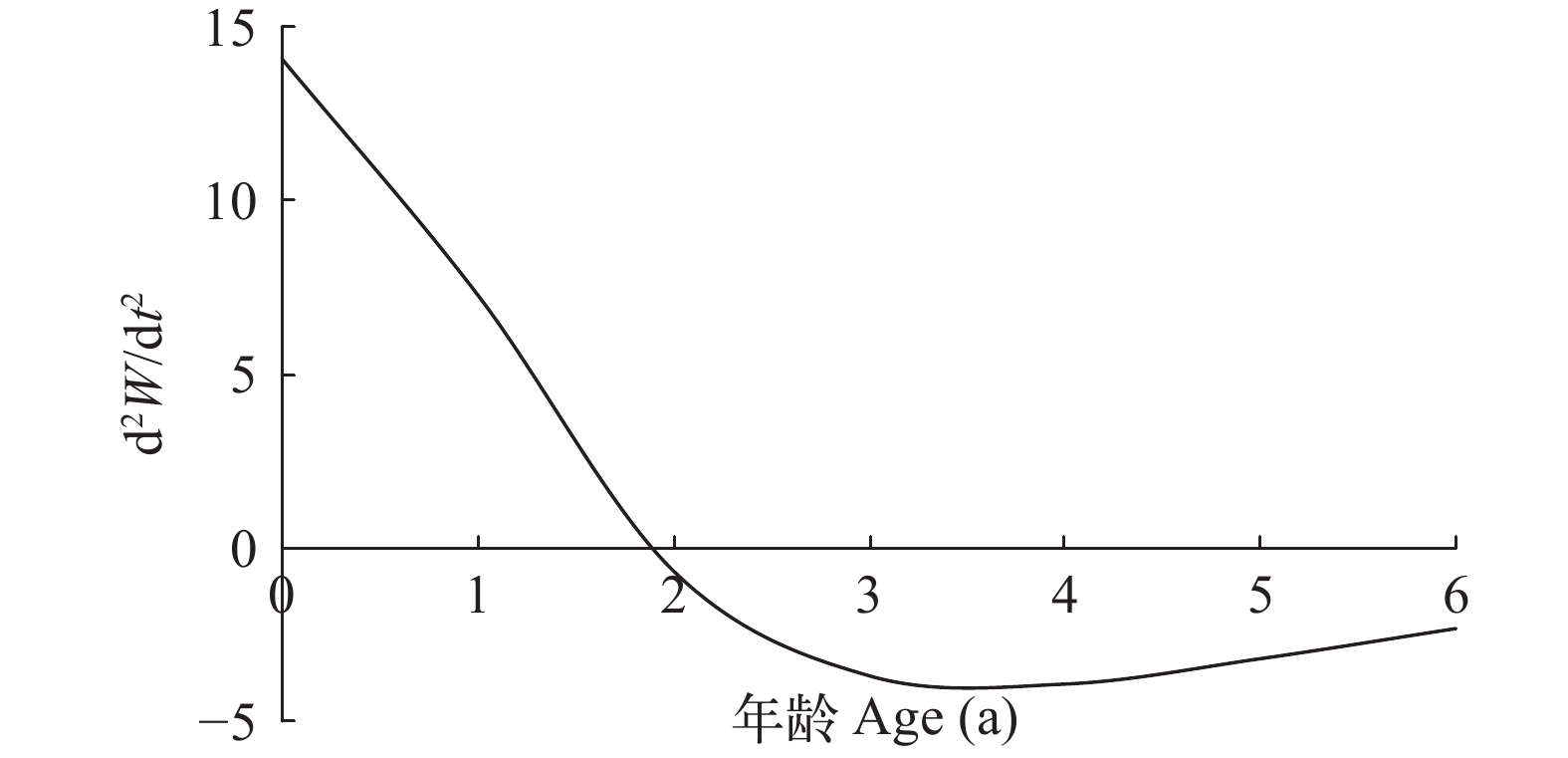
凤鲚体长生长速度曲线为渐近线, 不具有拐点(图 5), 并且生长速度随年龄的增加而减小; 体重具有一个生长拐点, 拐点年龄为1.85 a, 对应的拐点体重为25.42 g, 拐点体长为17.96 cm; 拐点年龄前体重生长速度逐渐增大, 拐点后逐渐减小。再将体重生长速度方程(dW/dt)对t求导, 进一步得到体重生长加速度方程。体重生长加速度曲线也具有一个拐点(图 6), 在体重生长拐点后, 体重生长加速度为负值。
![]() 图 5 瓯江口凤鲚的体长和体重生长速度曲线Figure 5. The growth curve of body length and body weight of Coilia mystus in Oujiang River Estuary
图 5 瓯江口凤鲚的体长和体重生长速度曲线Figure 5. The growth curve of body length and body weight of Coilia mystus in Oujiang River Estuary![]() 图 6 瓯江口凤鲚的体重生长加速度曲线Figure 6. The growth acceleration rate curve of body weight of Coilia mystus in Oujiang River Estuary
图 6 瓯江口凤鲚的体重生长加速度曲线Figure 6. The growth acceleration rate curve of body weight of Coilia mystus in Oujiang River Estuary2.3 死亡
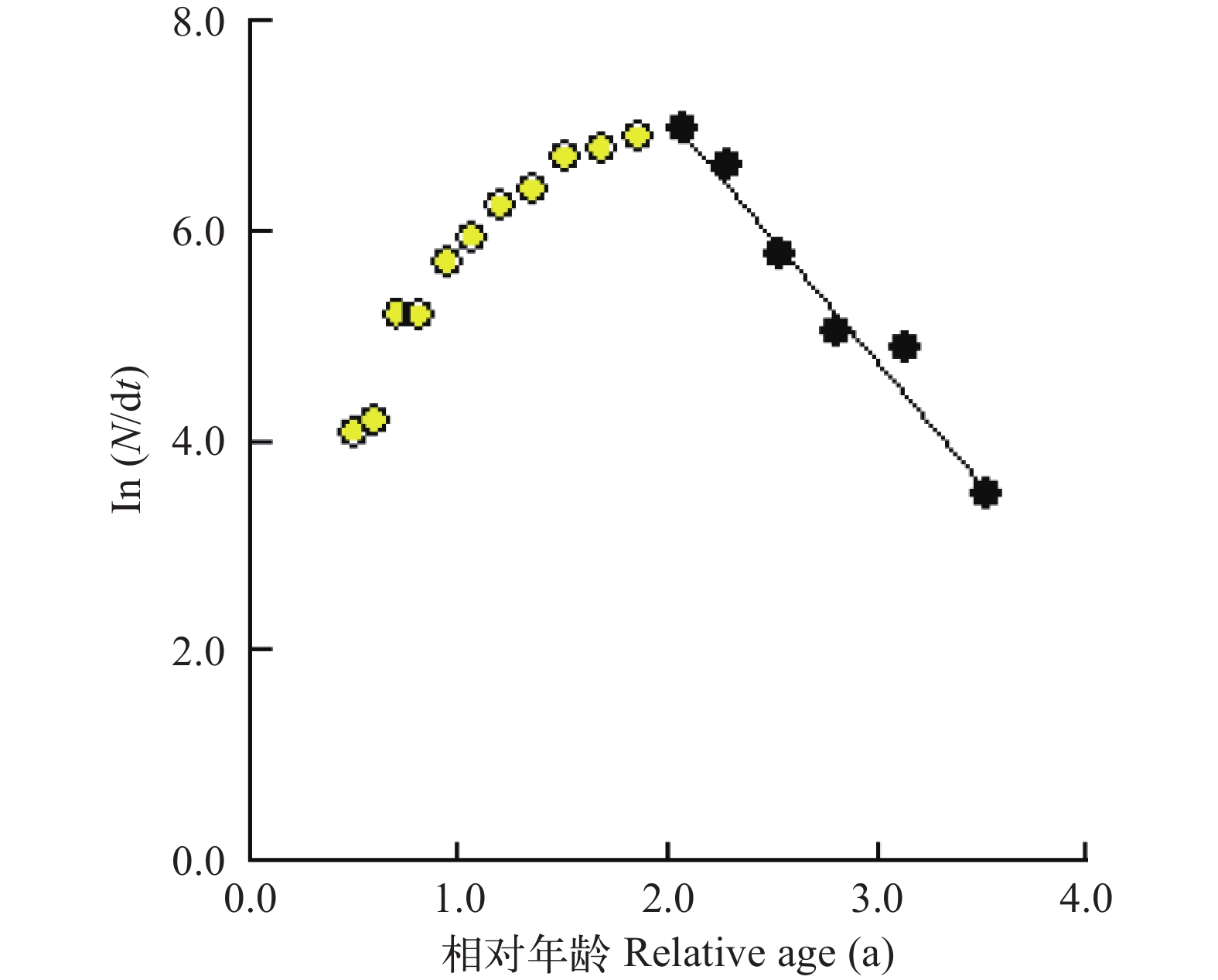
根据体长变换渔获曲线法估算总死亡系数, 本研究共选取6个点进行线性回归[29, 30](图 7)。方程的斜率为–2.30, 故所估算的瓯江口凤鲚的总死亡系数Z=2.30。根据体长-全长关系求得的渐近全长TL∞=37.76 cm, 调查期间瓯江水域平均水温为25.90℃, 代入Pauly经验公式求得自然死亡系数M=1.00。根据公式(5)求得捕捞死亡系数F=1.30, 由E=F/Z得出凤鲚的开发率为0.567。
![]() 图 7 根据体长变换渔获曲线估算瓯江口凤鲚的总死亡系数Figure 7. Estimation of total mortality parameter from length coverted catch curve of Coilia mystus in Oujiang River Estuary
图 7 根据体长变换渔获曲线估算瓯江口凤鲚的总死亡系数Figure 7. Estimation of total mortality parameter from length coverted catch curve of Coilia mystus in Oujiang River Estuary2.4 开捕体长和资源利用现状
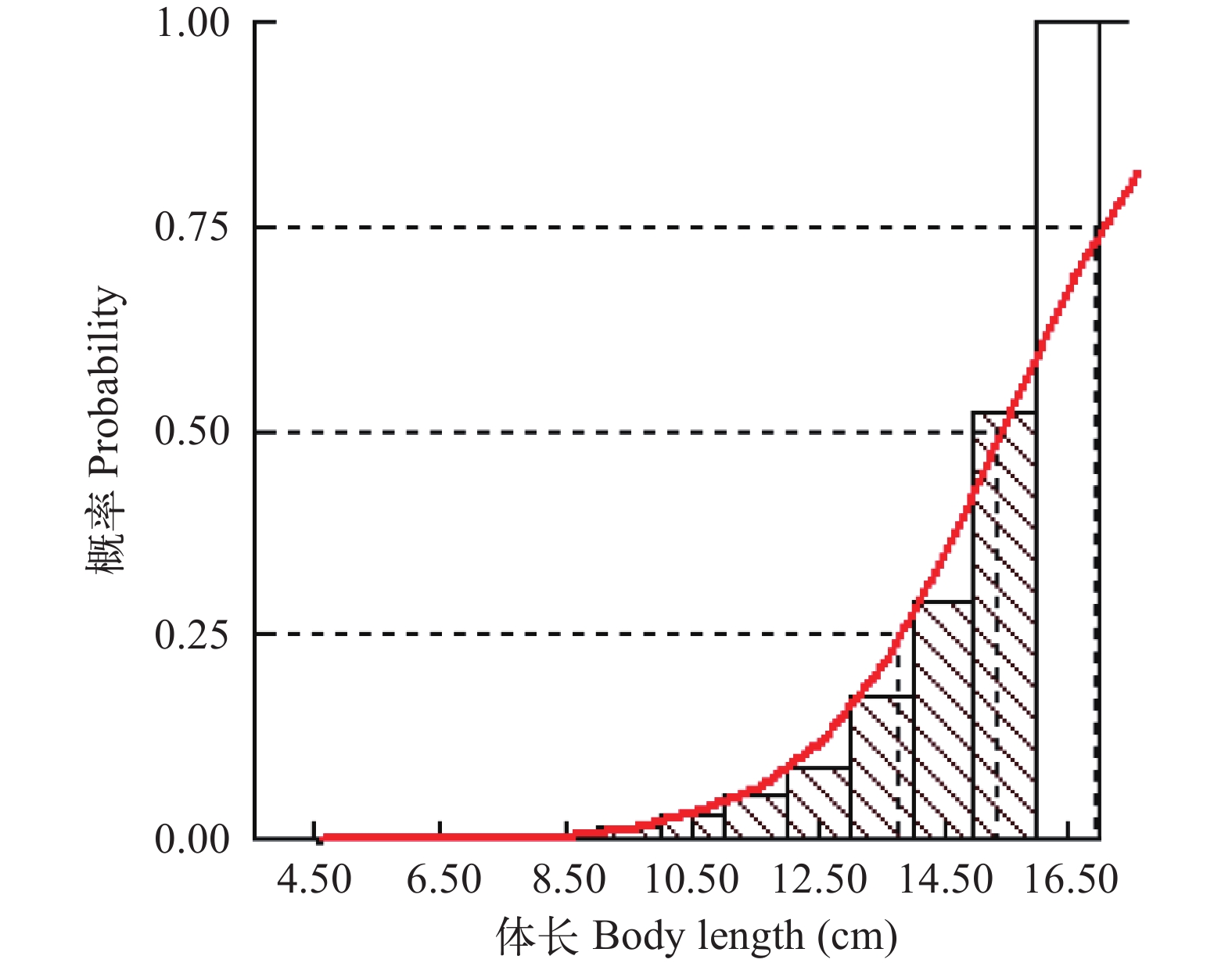
将凤鲚达到初次性成熟时的年龄作为补充年龄, 本研究实际测得的凤鲚最小性成熟体长为14.50 cm, 利用生长方程求得对应的年龄tr=1.13 a。根据渔获概率曲线图, 将期望值和观测值之比的累积率达到50%时的点所对应的体长(图 8)作为开捕体长Lc, 结果为15.35 cm, 对应的开捕年龄tc为1.28 a。
![]() 图 8 瓯江口凤鲚渔获概率曲线Figure 8. Curve of catch probability for Coilia mystus in Oujiang River Estuary
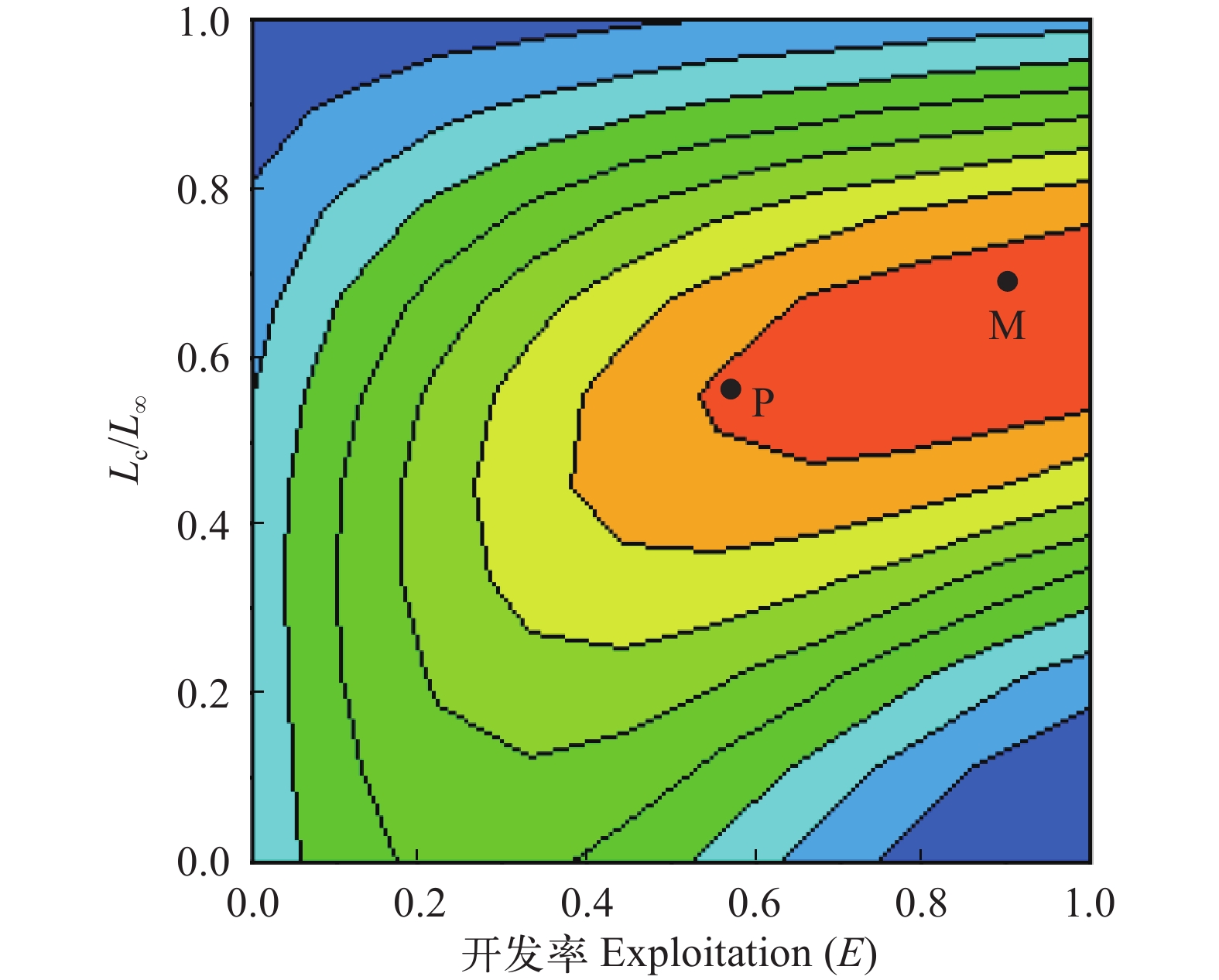
图 8 瓯江口凤鲚渔获概率曲线Figure 8. Curve of catch probability for Coilia mystus in Oujiang River Estuary应用“Beverton & Holt Y/R Analysis”模块, 输入M/k=2.12, 分析当前捕捞状态下瓯江口凤鲚Y'/R随E和Lc/L∞变化的等值线图(图 9), 当前捕捞状态P点的Lc/L∞=0.57, M/k=2.12, E=0.567, Y'/R=0.028, 而M/k=2.12时的理论最佳捕捞状态(M点)的Lc/L∞≈0.70、E≈0.90; 从P点调整到M点, Y'/R上升的比例远没有开发率大。当保持M/k=2.12, E=0.567时, Lc/L∞介于0.5—0.6时, 即Lc为13.30—15.96 cm时, Y'/R保持最大值0.028, 若继续提高开捕体长Lc, 则Y'/R随Lc/L∞的增大呈减小趋势。当前开捕体长位于Y'/R最大值的体长范围内。假设开捕体长稳定在现行状态, 从Y'/R、B'/R与E的二维分析图可见(图 10), 当E从现在的0.567增加到最大渔获量的开发率Emax=0.960, Y'/R达到最大值0.031, 但Y'/R的增幅远小于开发率的增幅, 开发率增大导致捕捞死亡系数也相应的增大。
![]() 图 9 瓯江口凤鲚相对单位补充量渔获量与开发率和开捕体长的关系M. 理论最佳开发状态Theoretic optimum exploitation; P. 当前捕捞状态Current exploitationFigure 9. Relative yield-per-recruit (Y′/R) of Coilia mystus in relation to exploitation ratio and body length at first catch
图 9 瓯江口凤鲚相对单位补充量渔获量与开发率和开捕体长的关系M. 理论最佳开发状态Theoretic optimum exploitation; P. 当前捕捞状态Current exploitationFigure 9. Relative yield-per-recruit (Y′/R) of Coilia mystus in relation to exploitation ratio and body length at first catch![]() 图 10 相对单位补充量渔获量(Y'/R)和相对单位补充资源量(B'/R)曲线图Figure 10. Two-dimensional analysis for relative yield per recruit, relative biomass per recruit and exploitation ratio of Coilia mystus in Oujiang River Estuary
图 10 相对单位补充量渔获量(Y'/R)和相对单位补充资源量(B'/R)曲线图Figure 10. Two-dimensional analysis for relative yield per recruit, relative biomass per recruit and exploitation ratio of Coilia mystus in Oujiang River Estuary3. 讨论
3.1 体长、体重
2006年5—8月用流刺网捕获的凤鲚雌性个体、雄性个体平均体长分别为(17.3±2.0)和(18.1±1.7) cm, 平均体重分别为(22.1±7.5)和(31.6±8.8) g[8]。与本研究结果对比, 雌性个体差别较小, 但雄性个体的平均体重相差较大, 可能是由于调查时间长短不同。鱼类体长-体重关系式W=a×Lb中, 幂指数b的变化与鱼类的生长阶段和对应的营养条件有关, b值一般介于2.5—3.5[12, 20]。长江口青草沙水库群体[31]、舟山群体[32]和九龙江口群体[33]的b值分别为3.0238、3.2120和3.2300, 本研究中b为3.0783, 经独立样本t检验, 不同水域的b值与3差异不显著, 说明这4个水域的凤鲚群体均呈匀速生长。
3.2 生长参数
传统的研究方法依靠鳞片[25, 34]和耳石[31, 35]开展年龄鉴定获取生长信息。近几年来随着计算机技术的发展, 体长频率分析技术因操作简便, 克服了传统年龄鉴定方法的主观性误差, 广泛应用于鱼类的生长研究, 为鱼类的生长研究提供了高效路径, 尤其适用于温带水域生长迅速、短寿命的小型鱼类[18]。当生长参数k满足e–k<1时, 表明von Bertalanffy方程能较好拟合生长, k值估算准确[36]。本研究根据ELEFAN估算出瓯江凤鲚渐近体长L∞=26.60 cm, k=0.47, e–k=0.63<1, k值的估算符合理论要求。比较各水域用体长频率估算法得出的凤鲚种群生长情况, 瓯江凤鲚L∞大于舟山[32](22.00 cm)和九龙江口[33](24.68 cm)凤鲚, 而k以九龙江口最大, 但其t0最小。不同水域凤鲚的生长参数存在明显差异, 可以解释为L∞、k和t0在计算过程中互相修正, 其组合可以体现凤鲚在不同海区生长由于种群密度、环境因子等栖息环境造成的差异[37, 38]。有学者在研究大泷六线鱼时提出利用不同研究方法得出的生长方程会导致结果有所偏差, 因此今后要对比通过凤鲚鳞片、耳石直接观测年龄和体长频率分析技术推算年龄这两种方法所得结果的准确性和差异性[39]。
3.3 死亡参数
鱼类死亡是指个体从资源群体中消失的状况, 是影响资源群体数量变动的主要因素[12, 18], 死亡程度的高低决定了资源群体数量的下降速度[12]。Beverton等[36]指出当M/K介于1.5—2.5时, 估算的M值比较准确合理, 当Z/K≤3时, 说明种群的死亡主要源于自然死亡, 反之则主要源于捕捞死亡[12]。在本研究中, M/K值为2.12, Z/K值为4.89, 表明M值符合理论要求且较为准确, 同时揭示捕捞是瓯江凤鲚种群死亡的主要影响因子。本次估算的M值(1.00)大于东海区凤鲚的M值(0.53)[40], 但生长速率小于东海区, 这可能是由于瓯江口调查期间处于夏季, 水温较高, 从而导致用Pauly公式计算的M值偏大, 这与林龙山提出的M值估算与调查水域栖息水温等环境因素有关相一致[18]。
Gulland[41]指出开发率E在0—0.5内, 说明资源群体处于轻度开发状态, 0.5为最适开发率, 大于0.5则为过度开发。本研究凤鲚的开发率为0.567, 表明目前瓯江凤鲚已处于过度开发状态, 而估算的M值表明凤鲚的死亡受捕捞影响较大。刘志坚等[5]分析认为捕捞强度过大是影响瓯江凤鲚资源衰减的重要原因之一, 与本研究的结论相一致。因此, 为降低凤鲚死亡率恢复瓯江凤鲚资源, 建议管理部门减轻捕捞压力, 降低其捕捞死亡率。
3.4 最适开捕规格
从凤鲚的生长特征出发, 由于拐点年龄前鱼类生长速度随年龄的增大而增大, 因此开捕年龄应控制在凤鲚生长最快阶段结束时, 此时能保障鱼类充分生长, 并能获得理论上最高的商业价值[15]。本研究得出凤鲚的拐点年龄tp为1.85 a(17.96 cm), 开捕年龄为1.28 a(15.35 cm), 开捕年龄小于拐点年龄; 从世代生物量变化的角度出发, 理想状态下资源生物量在临界年龄前不断增加, 其后则下降直至整个世代完全消失, 因此资源开发应控制在资源生物量下降前(即接近临界年龄)进行, 当前开捕年龄、Y'/R达到最大值时所对应的年龄均接近临界年龄(1.36 a, 15.76 cm); 从资源补充的角度出发, 要保证凤鲚一生至少有一次产卵机会, 因此开捕年龄大于最小性成熟年龄则更有利于资源的补充[32], 而本研究实际测得的凤鲚最小性成熟年龄为1.13 a(14.50 cm), 当前开捕年龄大于最小性成熟年龄。开捕年龄应控制在1.13—1.36 a, 当前开捕年龄在此范围中。对于瓯江口凤鲚当前开发状态, 规定开捕体长和降低捕捞强度均能使其处于较为合理的利用状态。依据本研究结果, 当前凤鲚的平均体长为15.06 cm, 优势体长组为14.0—18.0 cm(54.30%); 在体长组成中, 15.35 cm以上的个体占52.47%, 17.96 cm以上个体仅占14.05%, 说明目前大规格的凤鲚很少见, 多以小型的低龄个体为主。林龙山等[42]确立了凤鲚的开捕体长为14.00 cm, 本研究建议将凤鲚的开捕体长增大至15.50 cm(1.31 a), 以有效保护凤鲚资源, 保证凤鲚补充群体的数量, 防止凤鲚资源过度开发, 同时维护渔民的经济利益, 将开捕体长确立在渔民可接受范围内。
-
![]()
图 2 瓯江口凤鲚体长和体重分布
Figure 2. Body length and body weight distribution of Coilia mystus in Oujiang River Estuary
![]()
图 3 瓯江口凤鲚体长和体重的关系
Figure 3. The body length-weight relationship of Coilia mystus in Oujiang River Estuary
![]()
图 4 瓯江口凤鲚的体长和体重生长曲线
Figure 4. The growth curve in body length and body weight of Coilia mystus in Oujiang River Estuary
![]()
图 5 瓯江口凤鲚的体长和体重生长速度曲线
Figure 5. The growth curve of body length and body weight of Coilia mystus in Oujiang River Estuary
![]()
图 6 瓯江口凤鲚的体重生长加速度曲线
Figure 6. The growth acceleration rate curve of body weight of Coilia mystus in Oujiang River Estuary
![]()
图 7 根据体长变换渔获曲线估算瓯江口凤鲚的总死亡系数
Figure 7. Estimation of total mortality parameter from length coverted catch curve of Coilia mystus in Oujiang River Estuary
![]()
图 8 瓯江口凤鲚渔获概率曲线
Figure 8. Curve of catch probability for Coilia mystus in Oujiang River Estuary
![]()
图 9 瓯江口凤鲚相对单位补充量渔获量与开发率和开捕体长的关系
M. 理论最佳开发状态Theoretic optimum exploitation; P. 当前捕捞状态Current exploitation
Figure 9. Relative yield-per-recruit (Y′/R) of Coilia mystus in relation to exploitation ratio and body length at first catch
![]()
图 10 相对单位补充量渔获量(Y'/R)和相对单位补充资源量(B'/R)曲线图
Figure 10. Two-dimensional analysis for relative yield per recruit, relative biomass per recruit and exploitation ratio of Coilia mystus in Oujiang River Estuary
表 1 不同年份凤鲚的体长、体重分布
Table 1 Body length and body weight distribution in different year groups of Coilia mystus
年份
Year样本数
No. of
samples体长Body length (cm) 体重Body weight (g) 均值±
标准差
Mean±SD范围
Range均值±
标准差
Mean±SD范围
Range2015 429 16.1±2.1 8.2—21.9 19.5±9.2 2.2—53.6 2018 571 13.4±3.2 5.2—21.7 11.9±8.8 0.3—56.7 2019 121 15.7±2.8 6.8—21.9 17.6±10.0 0.9—44.0 2020 238 17.0±1.7 12.7—21.8 23.7±8.5 8.3—63.0  下载: 导出CSV
下载: 导出CSV
表 2 不同年龄组凤鲚的体长、体重分布
Table 2 Body length and body weight distribution in different age groups of Coilia mystus
年龄组
Age样本数
No. of
samples体长Body length (cm) 体重Body weight (g) 均值±
标准差
Mean±SD范围
Range均值±
标准差
Mean±SD范围
Range0 106 8.6±1.3 5.2—10.3 3.0±1.2 5.2—9.6 1 755 14.1±1.6 10.4—16.4 12.1±4.8 3.9—27.0 2 458 17.8±1.0 16.5—20.2 25.5±6.4 13.0—47.8 3 40 20.9±0.5 20.3—21.9 43.3±7.4 27.0—63.0
下载: 导出CSV
-
[1] 倪勇, 伍汉霖. 江苏鱼类志 [M]. 北京: 中国农业出版社, 2006: 204-205. Ni Y, Wu H L. Fishes of Jiangsu Province [M]. Beijing: China Agriculture Press, 2006: 204-205.
[2] 李明德, 张洪杰. 渤海鱼类生物学 [M]. 北京: 中国科学技术出版社, 1991: 21-22. Li M D, Zhang H J. Fish Biology of Bohai Sea [M]. Beijing: China Science and Technology Press, 1991: 21-22.
[3] 袁传宓, 林金榜, 秦安舲, 等. 关于我国鲚属鱼类分类的历史和现状—兼谈改造旧鱼类分类学的几点体会 [J]. 南京大学学报(自然科学版), 1976(2): 1-5. Yuan C M, Lin J B, Qin A L, et al. On the classification history and status quo of genus Coilia in China [J]. Journal of Nanjing University (Natural Science), 1976(2): 1-5
[4] 阎雪岚, 唐文乔, 杨金权. 基于线粒体控制区的序列变异分析中国东南部沿海凤鲚种群遗传结构 [J]. 生物多样性, 2009, 17(2): 143-150. doi: 10.3724/SP.J.1003.2009.08286 Yan X L, Tang W Q, Yang J Q. Population genetic structure of tapertail anchovy (Coilia mystus) in coastal waters of southeast China based on mtDNA control region sequences [J]. Biodiversity Science, 2009, 17(2): 143-150. doi: 10.3724/SP.J.1003.2009.08286
[5] 仲伟, 邵鑫斌, 胡利华, 等. 凤鲚瓯江种群的生物学特性 [J]. 温州大学学报(自然科学版), 2009, 30(4): 14-18. Zhong W, Shao X B, Hu L H, et al. Biological characteristics of Coilia mystus in the Oujiang River [J]. Journal of Wenzhou University (Natural Science), 2009, 30(4): 14-18.
[6] 刘志坚, 李德伟, 郭安托, 等. 瓯江口春夏季渔业生物种类组成及多样性 [J]. 浙江农业科学, 2016, 57(8): 1325-1327. Liu Z J, Li D W, Guo A T, et al. Species composition and diversity of fisheries in the Oujiang Estuary during spring and summer [J]. Journal of Zhejiang Agricultural Sciences, 2016, 57(8): 1325-1327.
[7] 陈锋, 赵先富, 赵进勇, 等. 瓯江鱼类资源调查及保护对策 [J]. 长江流域资源与环境, 2012, 21(8): 934-941. Chen F, Zhao X F, Zhao J Y, et al. Investigation of fish resources in the Oujiang River and its protection strategy [J]. Resources and Environment in the Yangtze Basin, 2012, 21(8): 934-941.
[8] 徐兆礼. 夏秋季瓯江口海域鱼类数量的时空分布 [J]. 动物学报, 2008, 54(6): 981-987. Xu Z L. Spatial-temporal distribution of fish density in the Oujiang Estuary during summer and autumn [J]. Acta Zoologica Sinica, 2008, 54(6): 981-987.
[9] 李凯, 范正利, 刘志坚, 等. 瓯江干流温州段鱼类群落结构的季节变化 [J]. 浙江海洋学院学报(自然科学版), 2017, 36(1): 9-13. Li K, Fan Z L, Liu Z J, et al. Seasonal variation of fish community structure in the Wenzhou section of Oujiang River mainstream [J]. Journal of Zhejiang Ocean University (Natural Science), 2017, 36(1): 9-13.
[10] 姜涛, 杨健, 刘洪波, 等. 刀鲚、凤鲚和湖鲚矢耳石的形态学比较研究 [J]. 海洋科学, 2011, 35(3): 23-31. Jiang T, Yang J, Liu H B, et al. A comparative study of the morphology of sagittal otolith in Coilia nasus, Coilia mystus and Coilia nasus taihuensis [J]. Marine Sciences, 2011, 35(3): 23-31.
[11] 刘琦, 张弛, 叶振江, 等. 东、黄海海鳗与山口海鳗生长和死亡参数的初步研究 [J]. 中国海洋大学学报(自然科学版), 2019, 49(S2): 46-52. Liu Q, Zhang C, Ye Z J, et al. A comparative study on growth and mortality of Muraenesox cinereus and M. bagio in East China Sea and Yellow Sea [J]. Periodical of Ocean University of China, 2019, 49(S2): 46-52.
[12] 詹秉义. 渔业资源评估 [M]. 北京: 中国农业出版社, 1995: 18-31. Zhan B Y. Fish Stock Assessment [M]. Beijing: China Agriculture Press, 1995: 18-31.
[13] 李大命, 唐晟凯, 刘燕山, 等. 滆湖鲢、鳙种群生长特征及起捕规格 [J]. 江苏农业科学, 2021, 49(6): 134-139. Li D M, Tang S K, Liu Y S, et al. Captured standard and growth characteristics of Hypophthalmichthys molitrix and Aristichthys mobilis in Lake Gehu [J]. Jiangsu Agricultural Sciences, 2021, 49(6): 134-139.
[14] 杨琴, 赵峰, 宋超, 等. 长江口及邻近海域凤鲚4个不同地理群体的形态差异 [J]. 海洋渔业., 2019, 41(3): 294-303. Yang Q, Zhao F, Song C, et al. Analysis of morphological variations among four different geographic populations of Coilia mystus in the Yangtze River Estuary and its adjacent waters [J]. Marine Fisheries, 2019, 41(3): 294-303.
[15] 杨琴, 赵峰, 宋超, 等. 长江口及邻近海域凤鲚生境履历重建 [J]. 中国水产科学, 2019, 26(6): 1175-1184. Yang Q, Zhao F, Song C, et al. Habitat history reconstruction of Coilia mystus from the Yangtze River Estuary and its adjacent sea area [J]. Journal of Fishery Sciences of China, 2019, 26(6): 1175-1184.
[16] 倪健夫, 郭弘艺, 唐文乔, 等. 长江靖江段沿岸凤鲚丰度的时间格局及生物学研究 [J]. 海洋渔业, 2020, 42(5): 560-570. doi: 10.3969/j.issn.1004-2490.2020.05.006 Ni J F, Guo H Y, Tang W Q, et al. Temporal distribution and biological characteristics of Coilia mystus in Jingjiang section of the Yangtze River [J]. Marine Fisheries, 2020, 42(5): 560-570. doi: 10.3969/j.issn.1004-2490.2020.05.006
[17] 孙超, 刘洪波, 姜涛, 等. 凤鲚ITS1与Cyt b基因序列的比较研究 [J]. 水产科学, 2013, 32(9): 536-540. doi: 10.3969/j.issn.1003-1111.2013.09.007 Sun C, Liu H B, Jiang T, et al. The comparison of ITS1 and Cyt b gene sequences in tapertail anchovy Coilia mystus [J]. Fisheries Science, 2013, 32(9): 536-540. doi: 10.3969/j.issn.1003-1111.2013.09.007
[18] 高春霞, 麻秋云, 田思泉, 等. 浙江南部近海小黄鱼生长、死亡和单位补充量渔获量 [J]. 中国水产科学, 2019, 26(5): 925-937. Gao C X, Ma Q Y, Tian S Q, et al. Growth, mortality and yield per recruitment of small yellow croaker in offshore waters of southern Zhejiang [J]. Journal of Fishery Sciences of China, 2019, 26(5): 925-937.
[19] Ricker W E. Linear regressions in fishery research [J]. Journal of the Fisheries Research Board of Canada, 1973, 30(3): 409-434. doi: 10.1139/f73-072
[20] Froese R. Cube law, condition factor and weight-length relationships: history, meta-analysis and recommendations [J]. Journal of Applied Ichthyology, 2006, 22(4): 241-253. doi: 10.1111/j.1439-0426.2006.00805.x
[21] Munro J L, Pauly D. A simple method for comparing the growth of fishes and invertebrates [J]. Fishbyte, 1983, 1(1): 5-6.
[22] Pauly D, Munro J L. Once more on the comparison of growth in fish and invertebrates [J]. Fishbyte, 1984, 2(1): 1-21.
[23] Von Bertalanffy L. A quantitative theory of organic growth (inquiries on growth laws. Ⅱ) [J]. Human Biology, 1938, 10(2): 181-213.
[24] Pauly D. Length-converted catch curves and the seasonal growth of fishes [J]. Fishbyte, 1990, 8(3): 33-38.
[25] 唐渝. 太湖湖鲚生长特征和临界年龄的研究 [J]. 生态学杂志, 1986, 5(3): 5-9,13. Tang Y. Study on the growth character and critical age of Coilia ectenes Taihuensis (Yuan et Lin) [J]. Chinese Journal of Ecology, 1986, 5(3): 5-9,13.
[26] 解崇友, 胡佐灿, 蔡瑞钰, 等. 三峡库区重要支流8种优势鱼类生长及其资源开发现状评估 [J]. 中国水产科学, 2019, 26(3): 504-511. doi: 10.3724/SP.J.1118.2019.18391 Xie C Y, Hu Z C, Cai R Y, et al. Assessment of the growth of eight dominant fish species and their resource development in important tributaries of the Three Gorges Reservoir area [J]. Journal of Fishery Sciences of China, 2019, 26(3): 504-511. doi: 10.3724/SP.J.1118.2019.18391
[27] Pauly D, Soriano M L. Some practical extensions to Beverton and Holt's relative yield-per-recruit model [C]//Maclean J L, Dizon L B, Hosillo L V (Eds.), The First Asian Fisheries Forum. Manila: Asian Fisheries Society, 1986: 491-496.
[28] 田辉伍, 段辛斌, 熊星, 等. 长江上游长薄鳅生长和种群参数的估算 [J]. 长江流域资源与环境, 2013, 22(10): 1305-1312. Tian H W, Duan X B, Xiong X, et al. Estimation of growth and population parameters of elongate loach (Leptobotia elongata) in the upper reaches of the Yangtze River [J]. Resources and Environment in the Yangtze Basin, 2013, 22(10): 1305-1312.
[29] 吴斌, 方春林, 傅培峰, 等. 鄱阳湖通江水道短颌鲚生长特性初探 [J]. 水生态学杂志, 2015, 36(3): 51-55. Wu B, Fang C L, Fu P F, et al. Growth characteristics of Coilia brachygnathus in the Poyang Lake-Yangtze River waterway [J]. Journal of Hydroecology, 2015, 36(3): 51-55.
[30] 孙远远, 昝肖肖, 徐宾铎, 等. 海州湾及邻近海域大泷六线鱼的生长、死亡和最适开捕体长研究 [J]. 中国海洋大学学报(自然科学版), 2014, 44(9): 46-52. Sun Y Y, Zan X X, Xu B D, et al. Growth, mortality and optimum catchable size of Hexagrammos otakii in Haizhou Bay and its adjacent waters [J]. Periodical of Ocean University of China (Natural Science), 2014, 44(9): 46-52.
[31] 于晓. 长江口凤鲚繁殖群体的生物学特性研究 [D]. 上海: 上海海洋大学, 2014: 26-27. Yu X. The research of biological characteristics of anadromous spawning Coilia mystus in Estuary of the Yangtze River [D]. Shanghai: Shanghai Ocean University, 2014: 26-27.
[32] 薛利建, 周永东, 徐开达, 等. 舟山近海凤鲚生长参数及资源量、持续渔获量分析 [J]. 福建水产, 2011, 33(2): 18-23, 32. doi: 10.3969/j.issn.1006-5601.2011.02.004 Xue L J, Zhou Y D, Xu K D, et al. Estimation of biological parameter, biomass, sustaining yield for Coilia mystus in the offshore of Zhoushan [J]. Journal of Fujian Fisheries, 2011, 33(2): 18-23, 32. doi: 10.3969/j.issn.1006-5601.2011.02.004
[33] 晁眉, 黄良敏, 李军, 等. 福建九龙江口凤鲚的生物学特征 [J]. 集美大学学报(自然科学版), 2016, 21(1): 16-20. Chao M, Huang L M, Li J, et al. Biological characteristics of Coilia mystus in Jiulong River Estuary in Fujian Province [J]. Journal of Jimei University (Natural Science), 2016, 21(1): 16-20.
[34] 王继隆, 唐富江, 刘伟. 大麻哈鱼的年龄与生长 [J]. 水生生物学报, 2012, 36(6): 1149-1155. doi: 10.7541/SP.J.1035.2012.01149 Wang J L, Tang F J, Liu W. Age and growth of chum salmon [J]. Acta Hydrobiologica Sinica, 2012, 36(6): 1149-1155. doi: 10.7541/SP.J.1035.2012.01149
[35] 水柏年. 黄海南部、东海北部小黄鱼的年龄与生长研究[J]. 浙江海洋学院学报(自然科学版), 2003, 22(1): 16-20. Shui B N. Study on the age and growth of Pseudosciaena polyactis in the south of the Yellow Sea and the north of the East China Sea [J]. Journal of Zhejiang Ocean University (Natural Science), 2003, 22(1): 16-20.
[36] Beverton R J H, Holt S J. A Review of the Lifespans and Mortality Rates of Fish in Nature, and Their Relation to Growth and Other Physiological Characteristics [C]//Wolstenholme G E W, O’Connor M (Eds.), Ciba Foundation Symposium: the Lifespan of Animals (Colloquia on Ageing), Chichester, UK: John Wiley & Sons, 1959: 142-180.
[37] Sainsbury K J. Effect of individual variability on the von Bertalanffy growth equation [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1980, 37(2): 241-247. doi: 10.1139/f80-031
[38] 陈作志, 邱永松, 黄梓荣. 南海北部白姑鱼生长和死亡参数的估算 [J]. 应用生态学报, 2005, 16(4): 712-716. doi: 10.3321/j.issn:1001-9332.2005.04.025 Chen Z Z, Qiu Y S, Huang Z R. Estimation of growth and mortality parameters of Argyrosomus argentatus in northern South China Sea [J]. Chinese Journal of Applied Ecology, 2005, 16(4): 712-716. doi: 10.3321/j.issn:1001-9332.2005.04.025
[39] 周永东, 薛利建, 徐开达. 舟山近海凤鲚Coilia mystus (Linnaeus)的生物学特性研究 [J]. 现代渔业信息, 2004, 19(8): 19-21. Zhou Y D, Xue L J, Xu K D. A study on biological characteristics of Coilia mystus (Linnaeus) offshore of Zhoushan [J]. Modern Fisheries Information, 2004, 19(8): 19-21.
[40] 林龙山, 郑元甲, 程家骅, 等. 东海区底拖网渔业主要经济鱼类渔业生物学的初步研究 [J]. 海洋科学, 2006, 30(2): 21-25. doi: 10.3969/j.issn.1000-3096.2006.02.005 Lin L S, Zheng Y J, Cheng J H, et al. A preliminary study on fishery biology of main commercial fishes surveyed from the bottom trawl fisheries in the East China Sea [J]. Marine Sciences, 2006, 30(2): 21-25. doi: 10.3969/j.issn.1000-3096.2006.02.005
[41] Gulland J A. Fish Stock Assessment: A Manual of Basic Methods [M]. New York: Wiley, 1983: 869-870.
[42] 林龙山, 程家骅, 凌建忠, 等. 东海区主要经济鱼类开捕规格的初步研究 [J]. 中国水产科学, 2006, 13(2): 250-256. doi: 10.3321/j.issn:1005-8737.2006.02.014 Lin L S, Cheng J H, Ling J Z, et al. First capture sizes of major commercial fishes in the East China Sea region [J]. Journal of Fishery Sciences of China, 2006, 13(2): 250-256. doi: 10.3321/j.issn:1005-8737.2006.02.014
-
期刊类型引用(3)
1. 张秋宏,孙浩奇,周永东,蒋日进,徐开达,徐国强,朱凯,李霞芳,蒋永生,渠遥,赵芃. 增殖放流背景下东海北部近海大黄鱼种群资源状况. 浙江海洋大学学报(自然科学版). 2024(02): 127-136 .  百度学术
百度学术
2. 王淼,洪波,张丹,晏军. 杭州湾北部凤鲚雌、雄繁殖群体的渔业生物学特征. 水产科学. 2024(05): 793-800 . 百度学术
3. 张泽鹏,金洪宇,迟淼,邵慧丽,李雷. 西藏朗错兰格湖裸鲤的生长、死亡及资源利用状况. 应用生态学报. 2023(07): 1988-1994 . 百度学术
其他类型引用(8)
计量
- 文章访问数: 1076
- HTML全文浏览量: 420
- PDF下载量: 44
- 被引次数: 11