
TOXIC EFFECTS OF DIHYDROARTEMISININ ON TETRAHYMENA THERMOPHILA
-
摘要: 以嗜热四膜虫(Tetrahymena thermophila)为试验对象, 双氢青蒿素(Dihydroartemisinin, DHA)以终浓度为0(对照组)、40、80、160和320 μmol/L 分别加入到嗜热四膜虫细胞培养液中, 探讨双氢青蒿素对嗜热四膜虫的毒性作用。采用 CCK-8 法检测嗜热四膜虫细胞增殖, 倒置显微镜和荧光显微镜观察细胞的形态结构及运动, 采用流式细胞术检测线粒体膜电位, 检测细胞内抗氧化还原酶活力和线粒体酶活力。结果表明, DHA显著抑制嗜热四膜虫增殖(P<0.05), 在一定暴露时间内增殖活力和浓度呈负相关。双氢青蒿素作用嗜热四膜虫48h后各暴露组细胞皱缩变圆, 对照组细胞呈椭圆状。其中在160和320 μmol/L DHA暴露下, 嗜热四膜虫在培养基中的活动减弱, 细胞核出现固缩和浓染等特征, 线粒体膜电位显著下降(P<0.05)。随着 DHA浓度增加, 细胞内超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GSH-Px)和谷胱甘肽硫转移酶(GST)活性先增强后下降。线粒体内琥珀酸脱氢酶(SDH)活性逐渐降低, 与对照组相比, 差异均有统计学意义(P<0.05)。上述结果表明, 双氢青蒿素对嗜热四膜虫具有毒性作用, 抗氧化酶在一定程度上能抵抗双氢青蒿素暴露导致的氧化损伤。氧化应激和线粒体损伤可能是双氢青蒿素对嗜热四膜虫产生毒性效应的重要机制。Abstract: To explore the toxicity of dihydroartemisinin (DHA) to Tetrahymena thermophila, T. thermophila was exposed to DHA (0, 40, 80, 160 and 320 μmol/L), and then cell proliferation, cell structures and movement, antioxidase activity, and mitochondrial membrane potential (MMP) were measured. The results showed that DHA inhibited the growth of T. thermophila in a concentration-dependent pattern. Observation through inverted microscope showed that after 48h treatment with DHA, T. thermophila became round and shrunken compared with the cells in the control group, which were oval. Meanwhile, high concentration treatment group (160 and 320 μmol/L) reduced cell movement, induced nucleus pycnosis, and decreased MMP significantly (P<0.05). With the increase of DHA concentration, the activities of the superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GSH-Px), and glutathione S-transferase (GST) increased at first and then decreased, and the level of succinate dehydrogenase (SDH) decreased significantly (P<0.05). The results indicated that the antioxidant enzymes in T. thermophila could prevent the oxidative damage caused by DHA exposure. Based on our results, the toxic effects of DHA on T. thermophila might be related with oxidative damage and mitochondrial damage.
-
青蒿素是中国科学家从传统中药青蒿中分离得到的倍半萜内酯类化合物的一种, 是抗疟疾特效药物[1]。双氢青蒿素是青蒿素在体内的有效活性代谢产物之一, 在体外可由四氢硼钠还原青蒿素而得到。双氢青蒿素不仅能有效防治疟原虫[2], 对其他寄生虫同样具有较好的杀灭效果[3-8]。很多寄生虫病分布范围广, 对人类健康和社会经济造成巨大危害。如车轮虫和多子小瓜虫是引起水生动物患病和死亡的主要寄生虫, 对水生生态和养殖业造成极大的损害[8]; 血吸虫能引起人兽共患的血吸虫病, 对该病的防治在公共卫生方面的重要性仅次于疟疾[9]。由于寄生虫离体培养困难, 并且寄生虫材料来源难以稳定获得, 因此采用与寄生虫分类地位相近的替代模式生物, 对于研究青蒿素类药物抗寄生虫的作用机制具有重要作用。
嗜热四膜虫(Tetrahymena thermophila)隶属于原生动物门纤毛亚门, 可在淡水环境中营自由生活。嗜热四膜虫是重要的单细胞真核模式生物, 遗传背景清楚且基因组序列的数据库完备, 以其为对象在细胞学和分子生物学的研究领域取得了一系列成果[10, 11]。此外嗜热四膜虫能在无菌液体培养基中培养, 培养方法简单, 个体小且繁殖周期短。大量研究表明, 嗜热四膜虫是研究药物[12]、有机化合物[13]和无机物[14]等外源化学物质潜在毒性的良好模式生物。
线粒体是机体能量代谢的中心, 还参与调控细胞增殖、凋亡等多种细胞活动[15]。线粒体介导的途径在青蒿素类药物诱导疟原虫细胞毒性中发挥重要作用[2]。线粒体通路途径可通过改变线粒体膜通透性, 引起线粒体膜电位降低和活性氧类增加, 促进氧化应激反应从而诱导细胞凋亡或死亡 [16, 17]。本文利用能体外培养的嗜热四膜虫为试验对象, 在嗜热四膜虫细胞培养液中定量加入不同剂量的双氢青蒿素, 研究DHA对细胞形态、运动行为等生理方面和细胞增殖、线粒体功能、氧化还原水平等生化方面的影响, 探讨DHA对嗜热四膜虫的毒性作用, 为评估青蒿素类药物在抗虫方面潜在的药用价值提供基础数据和科学依据。
1. 材料与方法
1.1 实验材料
嗜热四膜虫(B2086.2株由美国康奈尔大学赠送)虫种以5% 的体积分数接种于装有5 mL Neff培养液(0.25% 蛋白胨、0.25% 酵母提取物、0.5%葡萄糖和 9 g/L的FeCl3 母液按0.1% 体积比加入)的试管中, 于30℃、150 r/min 摇床培养48h后, 将处于对数生长期时的嗜热四膜虫虫液转移扩大培养。
双氢青蒿素的配制: 0.1 g DHA(99.9%, 中国食品药品检定研究院)用少量二甲基亚砜(DMSO)溶解, 配制成0.2 mol/L DHA储备液, 使用时用适量培养液稀释至实验所需浓度。
1.2 CCK8法检测细胞增殖活力
采用CCK8法测定双氢青蒿素对嗜热四膜虫细胞增殖的影响。移取对数生长期的嗜热四膜虫按1×104 个细胞/mL接种于10 mL新鲜培养液中。根据预实验结果, 设置DHA暴露组(浓度分别为40、80、160和320 μmol/L), 阴性对照组(0.01% DMSO)和空白对照组(无虫体的培养液), 在30℃、150 r/min条件下摇床培养24h、48h和72h。吸取100 μL至96孔板, 参考CCK8试剂盒(CK04-500T, 东仁化学科技上海有限公司)说明书所述方法, 用酶标仪(800TS, 美国伯腾仪器有限公司)于450 nm处测定吸光度(A)值。细胞增殖活力(%)=(A1–A0)/(A2–A0)×100%(其中A1、A2和A0分别为实验组、阴性对照组和空白组的A值)。每个浓度设4个复孔, 实验重复3次。
1.3 嗜热四膜虫细胞形态观察
移取对数生长期的嗜热四膜虫按1×104 个细胞/mL接种于10 mL新鲜培养液中。设置DHA暴露组(浓度分别为40、80、160和320 μmol/L)和对照组(0.01% DMSO)。于30℃、150 r/min 摇床培养48h。各组分别取200 μL依次加入24孔板中, 倒置显微镜(Ti-S, 日本Nikon公司)下观察细胞数目, 形态和运动状况。为了进一步观察细胞凋亡特征, 于培养48h 后, 3200 r/min
离心5min收集细胞, 弃上清。加沉淀2—3倍体积的Hoechst 33258染色液(C1017, 上海碧云天生物技术有限公司), 室温避光培养20min, 3200 r/min离心5min弃染液。用磷酸盐缓冲溶液(PBS, pH 7.4)洗2次, 取20 μL细胞悬液滴在载玻片上, 加入抗荧光淬灭封片液(P0126, 上海碧云天生物技术有限公司)。荧光显微镜(X73, 日本Olympus公司)下观察细胞核形态。 1.4 线粒体膜电位分析
细胞培养和实验分组同1.3。取DHA处理48h后的细胞, 经自动细胞计数仪计数, 收集4×104个细胞。按 JC-1试剂盒(C2006, 上海碧云天生物技术有限公司)使用说明书要求操作, 配置JC-1工作液及JC-1缓冲液(使用前稀释至1×), 四膜虫细胞置于JC-1工作液, 37℃ 染色20min, 离心取上清, 加JC-1缓冲液重悬离心, 重复洗涤2次后用适量JC-1缓冲液重悬细胞, 流式细胞仪(FACSCanto Ⅱ, 美国BD公司)检测, 分别采集红色荧光(激发波长525 nm, 发射波长585 nm)和绿色荧光(激发波长490 nm, 发射波长 530 nm)下的图像。横坐标为荧光强度, 纵坐标为细胞数量, 以红/绿荧光强度比值表示线粒体膜电位。实验重复3次。
1.5 嗜热四膜虫抗氧化酶活力
细胞培养和实验分组同1.3。取DHA处理48h后的细胞, 以3200 r/min
离心5min收集细胞, 弃上清。用生理盐水洗2次后加入一定量的PBS使细胞重新悬浮, 经液氮反复冻融使细胞裂解。将细胞悬液在4℃ 3200 r/min 离心5min, 取上清液待测。超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GSH-Px)、谷胱甘肽硫转移酶(GST)和琥珀酸脱氢酶(SDH)测定均采用南京建成生物工程研究所研制试剂盒, 按照相关说明书操作经分光光度计测定。 1.6 统计学分析
实验数据用平均值±标准差(mean±SD)表示, 采用 SPSS 13.0 软件进行统计处理分析。单因素方差分析(One way ANOVA)检验组间是否存在显著性差异, 如果存在显著性差异, 通过Tukey分析法进行多重比较, 显著性差异水平采用P<0.05。
2. 结果
2.1 双氢青蒿素对嗜热四膜虫细胞增殖的影响
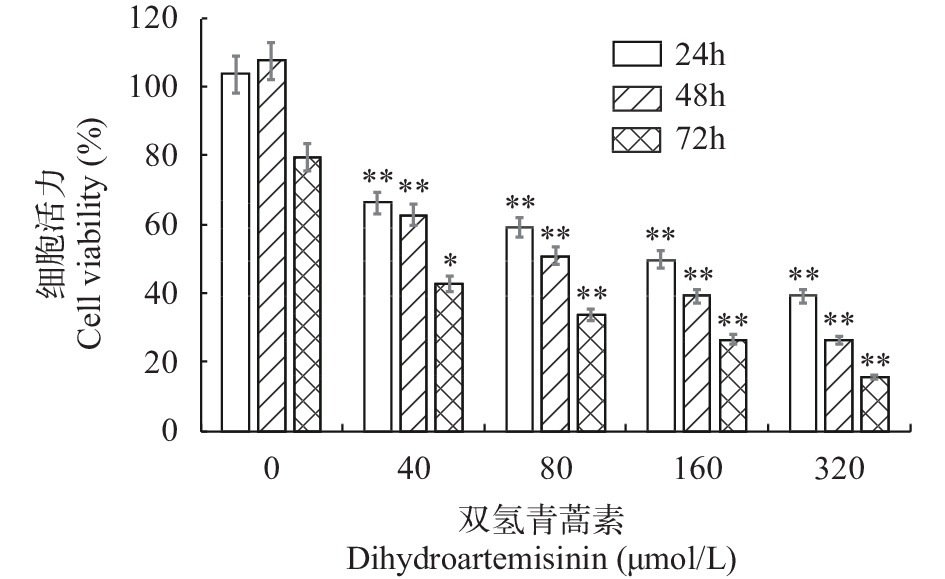
DHA作用嗜热四膜虫24h、48h和72h后, 采用 CCK-8法检测各浓度组、各时间点细胞增殖情况(图 1)。随着浓度增加, 嗜热四膜虫细胞增殖活力下降, 与对照组比较差异有统计学意义(P<0.05)。24h下, 40与80 μmol/L组, 80与160 μmol/L组的差异没有统计学意义。在48h和72h下各时间点组两两比较差异有统计学意义(P<0.05), 表现为增殖活力与浓度呈负相关。在相同DHA作用浓度下, 细胞增殖随着时间增加表现为逐渐降低, 72h时细胞增殖活力最低。根据实验结果, 选取DHA暴露48h作为时间样点进行后续试验。
![]() 图 1 双氢青蒿素对嗜热四膜虫细胞增殖的影响实验数据以平均值±标准差表示, *表示与对照组相比差异显著(P<0.05), **表示与对照组相比差异极显著(P<0.01)Figure 1. The effect of DHA on the proliferation of Tetrahymena thermophilaData are expressed as mean±standard deviation. The difference between the experimental group with the superscript and the control group is significant (*P<0.05, **P<0.01)
图 1 双氢青蒿素对嗜热四膜虫细胞增殖的影响实验数据以平均值±标准差表示, *表示与对照组相比差异显著(P<0.05), **表示与对照组相比差异极显著(P<0.01)Figure 1. The effect of DHA on the proliferation of Tetrahymena thermophilaData are expressed as mean±standard deviation. The difference between the experimental group with the superscript and the control group is significant (*P<0.05, **P<0.01)2.2 双氢青蒿素对嗜热四膜虫形态及运动的影响
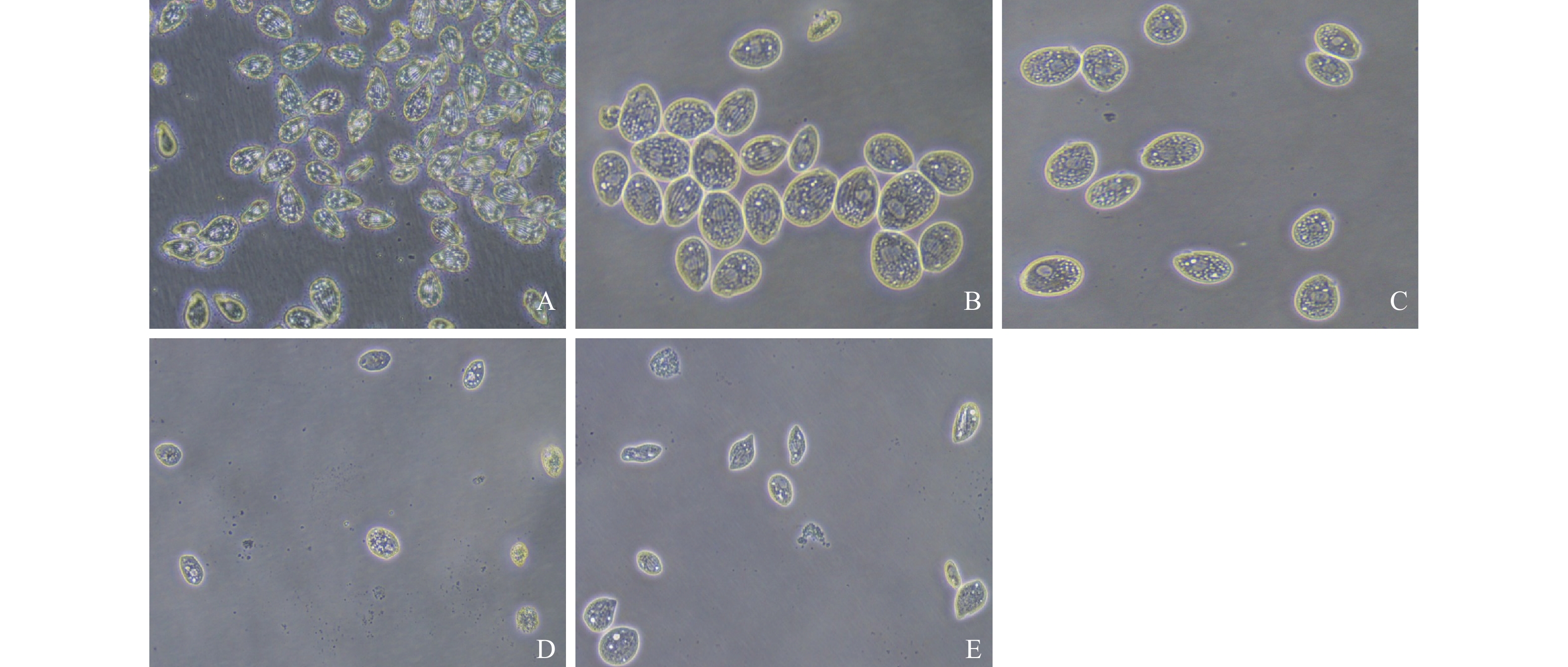
倒置显微镜下观察结果见图 2, 对照组嗜热四膜虫细胞呈椭圆长梨状, 细胞数目多, 活动速度快。DHA作用下各浓度组嗜热四膜虫形态发生改变, 细胞皱缩变圆, 胞质内空泡明显; 同时随着浓度增大, 细胞生长明显受到抑制, 活动速度逐渐减慢。与对照组比较, 160和320 μmol/L浓度组DHA处理细胞可见数量明显减少, 出现细胞破裂和异常细胞形态。
![]() 图 2 双氢青蒿素对嗜热四膜虫形态的影响 (比例尺=20 μm)A. 对照组; B、C、D和E 分别为40、80、160和320 μmol/L双氢青蒿素暴露组; 下同Figure 2. The effect of dihydroartemisinin on morphology of Tetrahymena thermophila (Scale bar=20 μm)A. Control group; B, C, D, E are experimental groups with the dihydroartemisinin concentrations of 40, 80, 160, and 320 μmol/L, respectively. The same applies below
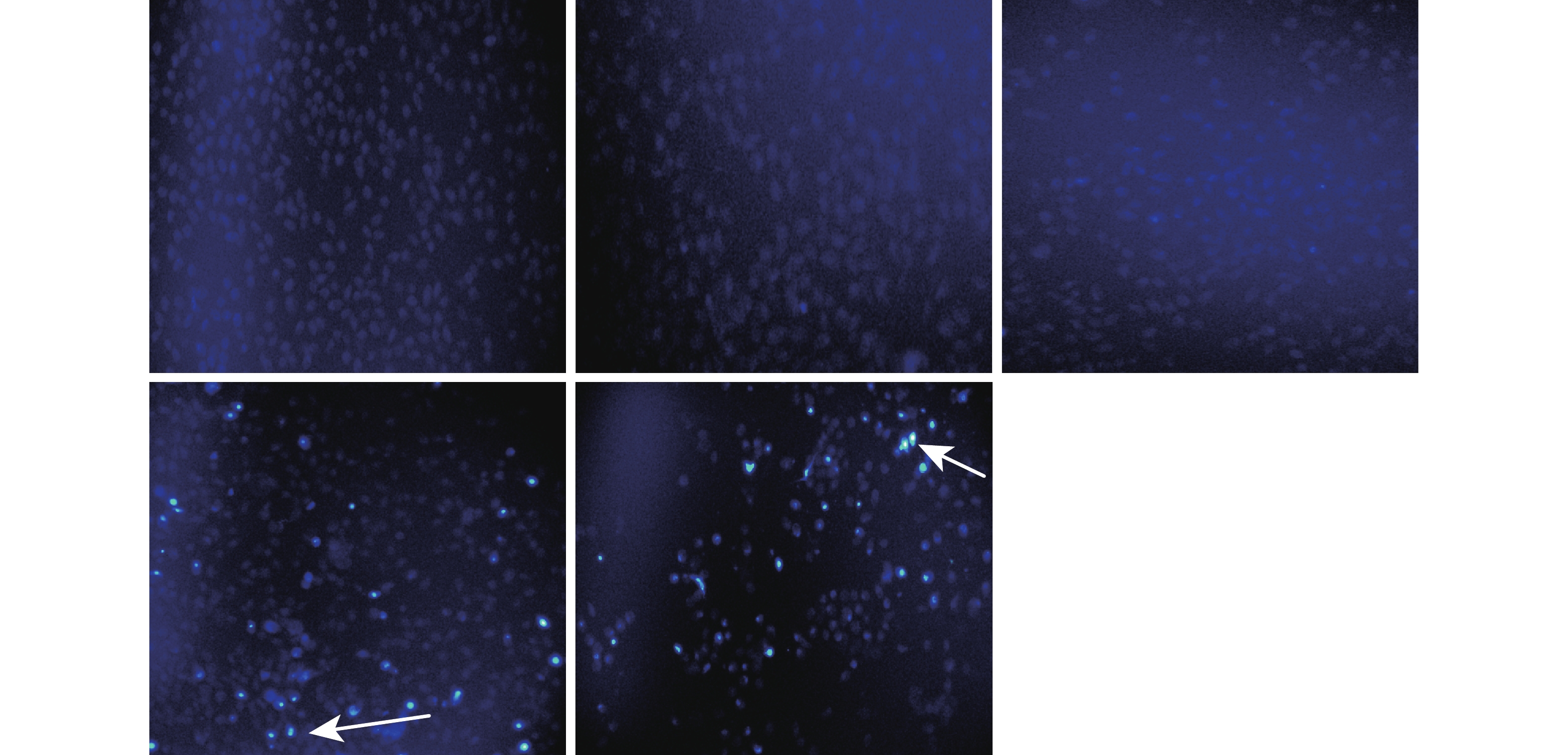
图 2 双氢青蒿素对嗜热四膜虫形态的影响 (比例尺=20 μm)A. 对照组; B、C、D和E 分别为40、80、160和320 μmol/L双氢青蒿素暴露组; 下同Figure 2. The effect of dihydroartemisinin on morphology of Tetrahymena thermophila (Scale bar=20 μm)A. Control group; B, C, D, E are experimental groups with the dihydroartemisinin concentrations of 40, 80, 160, and 320 μmol/L, respectively. The same applies belowHoechst 33258核染色结果见图 3。与对照组比较, 40和80 μmol/L浓度组DHA处理后细胞核形态没有发生明显变化。160和320 μmol/L浓度组DHA处理后, 细胞核呈浓染致密的颗粒状荧光, 且胞核固缩变小, 并出现凋亡小体(图 3箭头指示)。
![]() 图 3 双氢青蒿素对嗜热四膜虫细胞凋亡形态的影响(×400)Figure 3. The effect of dihydroartemisinin on apoptotic morphology of Tetrahymena thermophila (×400)
图 3 双氢青蒿素对嗜热四膜虫细胞凋亡形态的影响(×400)Figure 3. The effect of dihydroartemisinin on apoptotic morphology of Tetrahymena thermophila (×400)2.3 双氢青蒿素对嗜热四膜虫线粒体膜电位水平的影响
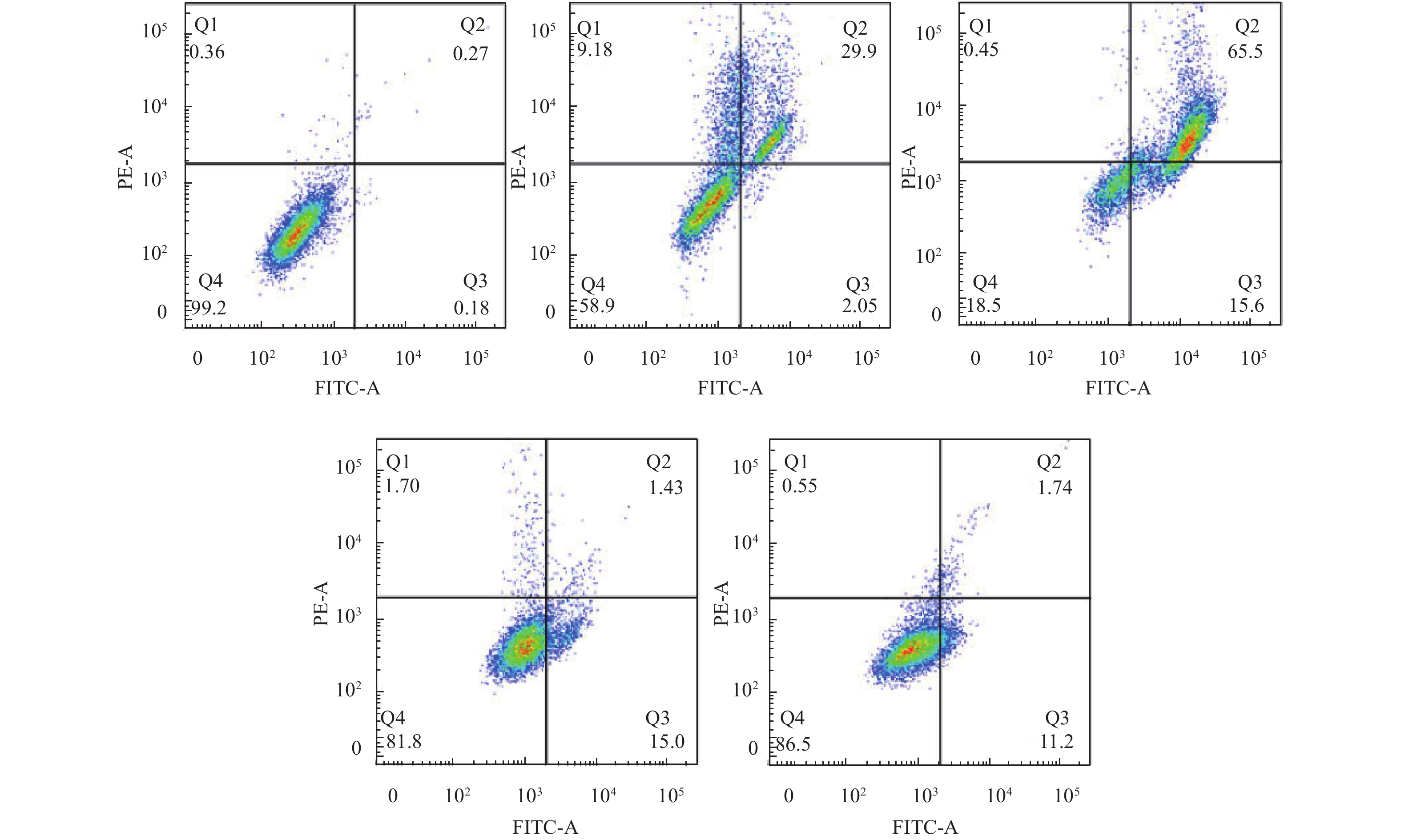
图 4中的细胞散点图所示, 对照组细胞集中在左下区, 红色荧光强, 线粒体膜电位高。随着DHA浓度的增加, 绿色荧光强度增加, 右下区细胞逐渐增多。对照组与40、80、160和320 μmol/L DHA实验组的红/绿荧光比值分别为(1.13±0.28)、(0.93±0.13)、(0.93±0.27)、(0.14±0.18)和(0.12±0.31)。与对照组相比, 160和320 μmol/L浓度组DHA处理后细胞的红/绿荧光比值显著升高, 差异有统计学意义(P<0.05)。这说明在一定浓度下双氢青蒿素可使嗜热四膜虫细胞线粒体膜电位下降。
![]() 图 4 双氢青蒿素对嗜热四膜虫细胞线粒体膜电位的影响Figure 4. The effect of dihydroartemisinin on mitochondrial membrane potential of Tetrahymena thermophila
图 4 双氢青蒿素对嗜热四膜虫细胞线粒体膜电位的影响Figure 4. The effect of dihydroartemisinin on mitochondrial membrane potential of Tetrahymena thermophila2.4 双氢青蒿素对嗜热四膜虫酶活力的影响
不同浓度DHA作用嗜热四膜虫后, 其抗氧化酶活力和SDH水平见表 1。在40、80和160 μmol/L DHA作用下, 细胞内抗氧化酶SOD、GSH-Px、GST和CAT活性明显高于对照组(P<0.05), 320 μmol/L DHA作用下细胞内抗氧化酶SOD、GSH-PX活性明显低于对照组(P<0.05), GST和CAT活性与对照组相比无显著性差异。随着DHA浓度增加, 抗氧化酶活力呈先升高后降低的趋势。DHA作用下各浓度组与对照组相比, 琥珀酸脱氢酶活力均显著降低(P<0.05)。
表 1 双氢青蒿素作用对嗜热四膜虫SOD、GSH-PX、GST、CAT和SDH活力的影响Table 1. The effect of dihydroartemisinin on enzyme activities in Tetrahymena thermophila (mean±SD, n=3; U/mg Prot)浓度
Concentration (μmol/L)SOD GSH-Px GST CAT SDH 0 20.16±0.34 10.37±1.68 61.42±0.05 18.30±3.45 43.67±5.05 40 23.24±1.58* 13.01±0.72** 67.50±1.12** 19.91±1.41* 41.80±3.59* 80 26.06±1.04** 14.44±1.16** 72.84±1.51** 21.18±2.52** 40.74±4.41* 160 36.82±2.35** 17.53±0.72** 78.03±1.40** 23.07±1.43** 37.62±2.45** 320 15.70±4.05** 8.86±2.53* 60.61±1.33 17.08±1.92 29.43±3.51** 注: 表中数据为3个重复的平均值; 同一列数据有上标符号的表示该实验组与同处理时间的对照组比较差异显著, 其中 *P<0.05, **P<0.01Note: Data are mean of triplicate.The difference between the experimental group with the superscript and the control group are significant (*P<0.05, ** P<0.01) 3. 讨论
DHA 是青蒿素的衍生物, 最先用于治疗疟疾, 同时也具有抗肿瘤、抑菌杀虫和免疫调节的功用。关于DHA抗寄生虫作用的研究, 发现DHA通过破坏原虫表膜-线粒体系统[3]来抑制伯氏疟原虫的生长; 通过对毛虫滋养体的蛋白质及细胞骨架的损伤作用, 来抑制蓝氏贾第鞭毛虫的生长[4]; 此外DHA 能破坏卡氏肺孢子虫滋养体和包囊[5]、虫膜系结构[6]而起到杀虫作用。伴随抗寄生虫药物耐药性挑战, 双氢青蒿素有望成为抗寄生虫新药, 但其杀虫机制尚待进一步阐明。因此我们采用能体外培养的嗜热四膜虫来研究双氢青蒿素的抗虫作用机制。本实验首先观察双氢青蒿素对嗜热四膜虫增殖和形态的影响, 结果显示与对照组相比, DHA作用下细胞增殖受到抑制, 细胞形态改变且运动速度减慢。研究表明青蒿琥脂、双氢青蒿素对嗜热四膜虫生长代谢有抑制作用, 但双氢青蒿素的抑制作用强于青蒿琥脂[18], 这可能与两者的构效不同有关。
当生物体暴露于外界毒物时, 抗氧化防御系统会构成机体的第一道防线, 抗氧化酶活力的改变可反映机体的氧化应激水平。关于外源化学物质对嗜热四膜虫生物胁迫作用的研究显示, H2O2处理后细胞线粒体膜电位丧失, 发生凋亡样死亡[19]。有机紫外防晒剂[20]和纳米材料[21]均可诱导嗜热四膜虫产生氧化应激, 导致抗氧化物质如SOD和GST等活力降低。在本研究中, DHA在40、80和160 μmol/L浓度下诱导细胞内抗氧化酶升高, 说明嗜热四膜虫表现出氧化应激。随着DHA浓度升高至320 μmol/L, 抗氧化酶水平降低。推测中低浓度DHA暴露下机体抗氧化防御酶活性增强以减轻DHA对细胞造成的氧化胁迫, 而高浓度DHA暴露下细胞内氧化损伤加重导致细胞活力降低, 进而破坏抗氧化酶导致酶水平降低。Ye等[12]研究发现, 高浓度钩吻素子(中药断肠草中的活性成分)作用嗜热四膜虫可引起氧化应激和凋亡, 低浓度下胁迫作用不明显。这与本实验结果存在一定差异, 不同化学物质对嗜热四膜虫的毒性大小有所区别, 因此细胞对氧化应激的适应性响应不同。
线粒体膜电位的稳定有利于维持细胞的正常生理功能[22]。有研究表明青蒿素在快速杀灭寄生虫过程中线粒体膜电位下降是主要生理事件, 而膜电位下降又与氧化应激催化生成的活性氧(ROS)有关[23]。琥珀酸脱氢酶参与线粒体能量代谢, 是反映线粒体功能的标志酶[24]。本实验结果显示, 一定浓度的DHA(160和320 μmol/L)暴露可降低嗜热四膜虫线粒体膜电位和琥珀酸脱氢酶活力(P<0.05), 这表明DHA暴露可造成线粒体功能损伤。在神经元细胞中也发现线粒体跨膜电位下降是蒿甲醚引起神经毒性的重要环节[25]。本实验结果与上述研究有相似之处, 青蒿素类药物对细胞的毒性作用可能与线粒体损伤有关[26]。结合Hoechst染色结果, 包括细胞核出现浓缩和染色体凝集等典型凋亡形态学改变, 故推测DHA暴露导致氧化应激, 引起活性氧过度堆积, 造成线粒体损伤, 最终诱导细胞凋亡。然而考虑到线粒体通路产生复杂的细胞传导通路和级联反应, 双氢青蒿素抗嗜热四膜虫的机制有待进一步研究。本研究现有结果表明DHA对嗜热四膜虫具有毒性效应, 氧化应激和线粒体损伤可能是毒性作用途径。
-
![]()
图 1 双氢青蒿素对嗜热四膜虫细胞增殖的影响
实验数据以平均值±标准差表示, *表示与对照组相比差异显著(P<0.05), **表示与对照组相比差异极显著(P<0.01)
Figure 1. The effect of DHA on the proliferation of Tetrahymena thermophila
Data are expressed as mean±standard deviation. The difference between the experimental group with the superscript and the control group is significant (*P<0.05, **P<0.01)
![]()
图 2 双氢青蒿素对嗜热四膜虫形态的影响 (比例尺=20 μm)
A. 对照组; B、C、D和E 分别为40、80、160和320 μmol/L双氢青蒿素暴露组; 下同
Figure 2. The effect of dihydroartemisinin on morphology of Tetrahymena thermophila (Scale bar=20 μm)
A. Control group; B, C, D, E are experimental groups with the dihydroartemisinin concentrations of 40, 80, 160, and 320 μmol/L, respectively. The same applies below
![]()
图 3 双氢青蒿素对嗜热四膜虫细胞凋亡形态的影响(×400)
Figure 3. The effect of dihydroartemisinin on apoptotic morphology of Tetrahymena thermophila (×400)
![]()
图 4 双氢青蒿素对嗜热四膜虫细胞线粒体膜电位的影响
Figure 4. The effect of dihydroartemisinin on mitochondrial membrane potential of Tetrahymena thermophila
表 1 双氢青蒿素作用对嗜热四膜虫SOD、GSH-PX、GST、CAT和SDH活力的影响
Table 1 The effect of dihydroartemisinin on enzyme activities in Tetrahymena thermophila (mean±SD, n=3; U/mg Prot)
浓度
Concentration (μmol/L)SOD GSH-Px GST CAT SDH 0 20.16±0.34 10.37±1.68 61.42±0.05 18.30±3.45 43.67±5.05 40 23.24±1.58* 13.01±0.72** 67.50±1.12** 19.91±1.41* 41.80±3.59* 80 26.06±1.04** 14.44±1.16** 72.84±1.51** 21.18±2.52** 40.74±4.41* 160 36.82±2.35** 17.53±0.72** 78.03±1.40** 23.07±1.43** 37.62±2.45** 320 15.70±4.05** 8.86±2.53* 60.61±1.33 17.08±1.92 29.43±3.51** 注: 表中数据为3个重复的平均值; 同一列数据有上标符号的表示该实验组与同处理时间的对照组比较差异显著, 其中 *P<0.05, **P<0.01Note: Data are mean of triplicate.The difference between the experimental group with the superscript and the control group are significant (*P<0.05, ** P<0.01)  下载: 导出CSV
下载: 导出CSV
-
[1] 李海波, 秦大鹏, 葛雯, 等. 青蒿化学成分及药理作用研究进展 [J]. 中草药, 2019, 50(14): 3461-3470. doi: 10.7501/j.issn.0253-2670.2019.14.029 Li H B, Qin D P, Ge W, et al. Research progress on chemical constituents of Artemisia annua and its pharmacological activities [J]. Chinese Traditional and Herbal Drugs, 2019, 50(14): 3461-3470. doi: 10.7501/j.issn.0253-2670.2019.14.029
[2] 吴芷静, 魏子巍, 李娇, 等. 抗疟疾药物研究进展 [J]. 动物医学进展, 2020, 41(3): 106-109. doi: 10.3969/j.issn.1007-5038.2020.03.021 Wu Z J, Wei Z W, Li J, et al. Progress on antimalarial drugs [J]. Progress in Veterinary Medicine, 2020, 41(3): 106-109. doi: 10.3969/j.issn.1007-5038.2020.03.021
[3] Chen P Q, Yuan J, Du Q Y, et al. Effects of dihydroartemisinin on fine structure of erythrocytic stages of Plasmodium berghei ANKA strain [J]. Acta Pharmacologica Sinica, 2000, 21(3): 234-238.
[4] 余源, 陈阳, 王卫亮, 等. 双氢青蒿素对蓝氏贾第鞭毛虫滋养体蛋白质的损伤 [J]. 中国人兽共患病学报, 2010, 26(11): 995-998. doi: 10.3969/j.issn.1002-2694.2010.11.002 Yu Y, Chen Y, Wang W L, et al. Effects of dihydroartemisinin on protein of Giardia lamblia trophozoites [J]. Chinese Journal of Zoonoses, 2010, 26(11): 995-998. doi: 10.3969/j.issn.1002-2694.2010.11.002
[5] 李文桂, 陈雅棠, 刘成伟, 等. 双氢青蒿素对大鼠体内卡氏肺孢子虫作用的超微结构研究 [J]. 中国地方病学杂志, 2004, 23(1): 28-31. Li W G, Chen Y T, Liu C W, et al. Ultrastructural study of effects of dihydroartemisinin on Pneumocystis carinii in rats [J]. Chinese Journal of Endemiology, 2004, 23(1): 28-31.
[6] 刘成伟, 陈雅棠, 李文桂, 等. 双氢青蒿素治疗实验大鼠卡氏肺孢子虫肺炎的透射电镜观察 [J]. 重庆医学, 2003, 32(6): 701-703. doi: 10.3969/j.issn.1671-8348.2003.06.031 Liu C W, Chen Y T, Li W G, et al. Transmission electron microscopic observation of pneumocystis carinii pneumonia in the immunosuppressed rats treated by dihydroartemisinin [J]. Chongqing Medicine, 2003, 32(6): 701-703. doi: 10.3969/j.issn.1671-8348.2003.06.031
[7] 茹炜炜, 梁幼生. 青蒿素类药物抗寄生虫作用研究进展 [J]. 中国血吸虫病防治杂志, 2006, 18(1): 78-80. doi: 10.3969/j.issn.1005-6661.2006.01.024 Ru W W, Liang Y S. Progress of research on artemisine against parasitic diseases [J]. Chinese Journal of Schistosomiasis Control, 2006, 18(1): 78-80. doi: 10.3969/j.issn.1005-6661.2006.01.024
[8] 王雪芹, 阳涛. 我国鱼类寄生虫病的防治策略及研究现状 [J]. 水产养殖, 2020, 41(11): 5-10. doi: 10.3969/j.issn.1004-2091.2020.11.002 Wang X Q, Yang T. Prevention and control strategies and research status of fish parasitosis in China [J]. Journal of Aquaculture, 2020, 41(11): 5-10. doi: 10.3969/j.issn.1004-2091.2020.11.002
[9] Colley D G, Bustinduy A L, Secor W E, et al. Human schistosomiasis [J]. Lancet, 2014, 383(9936): 1106-1118.
[10] Eisen J A, Coyne R S, Wu M, et al. Macronuclear genome sequence of the ciliate Tetrahymena thermophila, a model eukaryote [J]. PLoS Biology, 2006, 4(9): 1620-1642.
[11] 缪炜. 原生动物四膜虫“小材”有“大用” [J]. 生物学通报, 2010, 45(12): 1-4. doi: 10.3969/j.issn.0006-3193.2010.12.001 Miao W. Tetrahymena thermophila is a small fish in a big pond [J]. Bulletin of Biology, 2010, 45(12): 1-4. doi: 10.3969/j.issn.0006-3193.2010.12.001
[12] Ye Q, Zhang C N, Wang Z L, et al. Induction of oxidative stress, apoptosis and DNA damage by koumine in Tetrahymena thermophila [J]. PLoS One, 2019, 14(2): 1-15.
[13] 潘厚军, 张銮妫, 梁慧丽, 等. 应用嗜热四膜虫评估苯甲酸钠的生物毒性 [J]. 食品与生物技术学报, 2016, 35(5): 517-524. doi: 10.3969/j.issn.1673-1689.2016.05.011 Pan H J, Zhang L G, Liang H L, et al. Application Tetrahymena thermophila for assessment biotoxicity of sodium benzoate [J]. Journal of Food Science and Biotechnology, 2016, 35(5): 517-524. doi: 10.3969/j.issn.1673-1689.2016.05.011
[14] Pan Y B, Lin S J, Zhang W J. Epigenetic effects of silver nanoparticles and ionic silver in Tetrahymena thermophila [J]. Science of the Total Environment, 2021(768): 144659-144665.
[15] Hernandez-Resendiz S, Buelna-Chontal M, Correa F, et al. Targeting mitochondria for cardiac protection [J]. Current Drug Targets, 2013, 14(5): 586-600. doi: 10.2174/1389450111314050008
[16] Baechler B L, Bloemberg D, Quadrilatero J. Mitophagy regulates mitochondrial network signaling, oxidative stress, and apoptosis during myoblast differentiation [J]. Autophagy, 2019, 15(9): 1606-1619. doi: 10.1080/15548627.2019.1591672
[17] 申屠路媚, 牟艳玲. 线粒体功能障碍机制及其相关疾病研究进展 [J]. 生命科学, 2018, 30(1): 87-93. Shentu L M, Mu Y L. Research progress in mitochondrial dysfunction and its related diseases [J]. Chinese Bulletin of Life Sciences, 2018, 30(1): 87-93.
[18] Shen X S, Su Q, Qiu Z P, et al. Effects of artemisinin derivative on the growth metabolism of Tetrahymena thermophila BF5 based on expression of Thermokinetics [J]. Biological Trace Element Research, 2010, 136(1): 117-125. doi: 10.1007/s12011-009-8527-2
[19] 凌子惠, 张思维, 曹怡, 等. 细胞应激条件下嗜热四膜虫线粒体膜电位变化研究 [J]. 复旦学报, 2012, 51(3): 283-289. Ling Z H, Zhang S W, Cao Y, et al. Loss of mitochondrial membrane potential induces the accumulation of ROS in Tetrahymena thermophila under cellular stress [J]. Journal of Fudan University (
Natural Science ) , 2012, 51(3): 283-289. [20] 高礼, 周川琪, 袁涛, 等. 环境相关浓度有机紫外防晒剂对嗜热四膜虫抗氧化酶活力的影响 [J]. 环境与健康杂志, 2018, 35(11): 955-958. Gao L, Zhou C Q, Yuan T, et al. Effects of organic UV filters on enzyme activities in Tetrahymena thermophila at environmentally relevant concentrations [J]. Journal of Environment and Health, 2018, 35(11): 955-958.
[21] 廖苑辰, 常叶倩, 徐晨珂, 等. 氧化石墨烯对嗜热四膜虫的毒性效应 [J]. 中国环境科学, 2019, 39(3): 1299-1305. doi: 10.3969/j.issn.1000-6923.2019.03.048 Liao Y C, Chang Y Q, Xu C K, et al. Toxicity effects of graphene oxide to Tetrahymena thermophila [J]. China Environmental Science, 2019, 39(3): 1299-1305. doi: 10.3969/j.issn.1000-6923.2019.03.048
[22] Chowra U, Yanase E, Koyama H, et al. Aluminium-induced excessive ROS causes cellular damage and metabolic shifts in black gram Vigna mungo (L.) Hepper [J]. Protoplasma, 2017, 254(1): 293-302. doi: 10.1007/s00709-016-0943-5
[23] Antoine T, Fisher N, Amewu R, et al. Rapid kill of malaria parasites by artemisinin and semi-synthetic endoperoxides involves ROS-dependent depolarization of the membrane potential [J]. Journal of Antimicrobial Chemotherapy, 2014, 69(4): 1005-1016. doi: 10.1093/jac/dkt486
[24] Miyadera H, Shiomi K, Ui H, et al. Atpenins, potent and specific inhibitors of mitochondrial complex Ⅱ (succinate-ubiquinone oxidoreductase) [J]. Proceedings of the National Academy of Sciences, 2003, 100(2): 473-477. doi: 10.1073/pnas.0237315100
[25] 赵艳红, 王京燕. 蒿甲醚引起线粒体跨膜电位下降在神经毒性中的作用 [J]. 中国寄生虫学与寄生虫病杂志, 2003, 21(6): 326-329. doi: 10.3969/j.issn.1000-7423.2003.06.003 Zhao Y H, Wang J Y. Effect of mitochondrial membrane potential in the neurotoxicity of artemether [J]. Chinese Journal of Parasitology and Parasitic Diseases, 2003, 21(6): 326-329. doi: 10.3969/j.issn.1000-7423.2003.06.003
[26] 尹纪业, 王和枚, 丁日高. 青蒿素及其衍生物毒理学研究进展 [J]. 中国药理学与毒理学杂志, 2014, 28(2): 309-314. doi: 10.3867/j.issn.1000-3002.2014.02.028 Yin J Y, Wang H M, Ding R G. Artemisinin and its derivatives: progress in toxicology [J]. Chinese Journal of Pharmacology and Toxicology, 2014, 28(2): 309-314. doi: 10.3867/j.issn.1000-3002.2014.02.028
-
期刊类型引用(4)
1. 仲悦宁,杨昊憬,张逸博,吴振兵,党晨原,付杰. 污水处理厂活性污泥工艺中微型动物的生活习性、功能作用、研究方法与毒性响应. 净水技术. 2024(05): 18-34 .  百度学术
百度学术
2. 高帆,冯佳,谢树莲. 大学生创新创业训练计划的生物科学专业实验教学改革探索. 实验室研究与探索. 2023(07): 145-150+155 . 百度学术
3. 郭玉婷,郑海荣,张钰,吴清清,朱宇红,张新新,张继业. 青蒿的化学成分及药理作用的最新研究进展. 西北药学杂志. 2023(06): 241-249 . 百度学术
4. 胡青,毕保良. 水产养殖学专业课程思政元素探索——以水生生物学为例. 现代农业科技. 2023(22): 157-161 . 百度学术
其他类型引用(0)
计量
- 文章访问数: 2957
- HTML全文浏览量: 1022
- PDF下载量: 38
- 被引次数: 4