
CONNECTIVITY LOSS IN LAKE ON FISH COMMUNITIES ——A CASE STUDY OF BAO’AN LAKE
-
摘要: 为探讨湖泊内部阻隔引发的生态问题, 阐释鱼类群落聚合对湖泊内部阻隔的响应, 研究选取长江中游典型阻隔湖泊保安湖为研究对象, 于2019—2020年夏季和秋季对其肖四海湖区(完全阻隔)、扁担塘湖区(半阻隔)、桥墩湖区(半阻隔)和主体湖区的鱼类群落结构进行了调查, 应用单因素方差分析、置换多元方差分析和非度量多维尺度等多重统计方法分析了湖区间鱼类群落组成和多样性的差异。结果表明, 完全阻隔湖区肖四海鱼类群落结构发生了明显变化, 其种类数[(15±3)种]显著低于半阻隔湖区[(22±3)种]和主体湖区[(23±3)种, P<0.05], 鱼类丰度高而生物量低, 物种丰富度指数、Shannon指数和Simpson指数显著低于主体湖区(P<0.05), 功能丰富度指数、功能分散度指数和功能均匀度指数也显著较低(P<0.05); 而半阻隔湖区鱼类群落结构与主体湖区无显著性差异。置换多元方差分析和非度量多维尺度分析也显示出完全阻隔湖区鱼类群落与半阻隔湖区和主体湖区鱼类群落具有显著性差异(P<0.05), 半阻隔湖区与主体湖区无显著性差异。研究发现湖泊内部水文阻隔对鱼类群落组成也有着重要影响, 恢复湖泊内部水文自由连通对湖泊生态管理和生物多样性保护有着重要作用。Abstract: Hydrological connection plays an important role in maintaining biodiversity and ecosystem function. In order to explain the effect of internal hydrological disconnection and response of fish communities, we selected a typical hydrological disconnection lake in middle reaches of Yangtze River-Bao’an Lake as the study object. Field sampling was conducted in Xiaosihai Lake (part of Bao’an Lake, complete disconnection), Biandantang Lake (part of Bao’an Lake, semi-disconnection), Qiaodun Lake (part of Bao’an Lake, semi-disconnection) and the main lake in both summer and autumn from 2019 to 2020. We compared and analyzed the differences of fish communities and functional diversity among those lakes using the multiple statistical methods. Result showed that the fish community structure in the complete disconnection lake had been changed significantly, which species number (15±3) was significantly lower than that in the semi- disconnection lake (22±3) and the main lake (23±3; P<0.05); abundance increased while the biomass decreased; the species richness index, the Shannon index and the Simpson index were significantly lower; and functional richness index, functional dispersion index and functional evenness index were also significantly lower the other lakes (P<0.05). However, the fish community structure in the semi-disconnection lake had no significant difference with that in the main lake. The Permutational multivariate analysis of variance and non-metric multidimensional scaling analysis also showed that the fish community in the complete disconnection lake significantly distinguished from the semi-disconnection lakes and the main lake (P<0.05), while there was no significant difference between the fish communities of semi-disconnection lakes and main lake was closer. Our study found that the internal hydrological disconnection in lake also has an important impact on the composition of fish communities, and restoring the free hydrological connectivity within lake was crucial to ecological management and biodiversity conservation.
-
Keywords:
- Hydrological disconnection /
- Fish community /
- Diversity /
- Ecological impact /
- Bao’an Lake
-
拒马河原名涞水河, 是海河流域大清河北部支流, 发源于河北省涞源县西北太行山麓, 流经紫荆关向北至涞水县折向东流, 至北京市张坊镇分为南、北二支, 南拒马河目前已经干涸消亡, 北拒马河于白沟镇流入大清河, 拒马河干流长254 km, 张坊镇以上流域面积5115 km2, 是中国北方最大的冬季不结冰河流[1—4]。拒马河流域地貌复杂, 不同河段气候与环境相差悬殊, 生境的多样性造就了丰富的水生生物资源, 是华北地区内陆水域生物多样性的重要组成部分, 具有极高的科研及保护价值[5, 6]。
河流及湖泊水文环境的改变影响其中鱼类群落的结构组成, 而鱼类群落的结构组成也是水生生态系统健康的重要指标, 可通过鱼类群落的变动来分析水环境状况的受污染程度[7, 8]。近年来, 拒马河生态环境发生了巨大变化, 生态流量逐年下降, 水生态环境受人类影响较大[5], 水质、水文流通性发生改变, 改变了生活在其中的鱼类的栖息环境, 增加了鱼类的生存压力[9]。拒马河拦河坝的建设导致河流片段化严重, 纵向连通几乎丧失, 鱼类洄游通道被阻断, 部分河段形成小型静水湖泊, 洄游性、亲流性和因拦河坝建设而失去栖息及越冬场所的鱼类减少而喜静水性鱼类增加[10, 11]。加深鱼类群落在空间和时间上对生境改变做出反应的认知对制定有效的生态环境保护策略非常重要[12], 国内关于鱼类群落与环境因子的关系的研究多集中于近海和河口, 对于内陆河流的研究较为匮乏[13—15]。目前, 拒马河鱼类资源的调查虽有不少资料, 但了解得仍然不够深入和系统, 多仅限于北京段, 如2008年杨文波等[16]对拒马河北京段的鱼类资源调查, 记录鱼类24种; 张春光等[17]在《北京及其邻近地区的鱼类》中整理历史资料, 记录拒马河历史上出现鱼类42种; 袁立来等[6]利用鱼类生物完整性指数对拒马河北京段进行了河流健康评价, 发现拒马河北京段河流健康整体处于较差水平。对拒马河上游区域的物种组成和地理分布等基本问题仍缺乏准确的数据, 因此, 系统开展拒马河鱼类资源调查, 以掌握拒马河鱼类时空分布及影响因子, 是拒马河鱼类资源开发利用和资源保护首要解决的问题。
本研究基于2019—2021年间6次拒马河鱼类资源调查数据, 从物种组成、优势种及群落结构特征等方面分析了鱼类群落结构现状, 并讨论了鱼类资源的时空分布与环境因子的关系, 为拒马河流域内的鱼类资源的保护、修复和管理提供理论基础和科学依据。
1. 材料与方法
1.1 样品采集
本次调查范围自涞源县拒马河源头至张坊镇龙安大桥, 采样点的选择参考《内陆水域渔业自然资源调查规范》[18], 综合考虑拒马河河流特点及采样点的代表性, 设置了15个采样点, 采样点的经纬度和位置见表 1和图 1。于2019年5月 (春季)、8月 (夏季)、10月 (秋季), 2020年8月、10月和2021年5月进行采样调查, 每个采样点设置两条多目刺网和地笼, 刺网规格20 m×1 m和25 m×1.5 m, 网目为3和5 cm, 地笼规格为10 m×35 cm×30 cm和10 m×25 cm×20 cm, 网目为1 cm。刺网和地笼于16:00—18:00时下网, 次日5:00—7:00起网, 放置时间约12h。依据最新鱼类学专著对采集的渔获物分类[19—21], 对新鲜鱼类样本现场进行物种鉴定和个体测量, 采用游标卡尺和电子天平测量体长(精确到1 mm)和体重(精确到0.1 g)数据, 现场无法鉴定的个体用95%的酒精保存后带回实验室进一步分类鉴定。
表 1 拒马河采样点分布Table 1. Distribution of sampling stations in the Juma River采样点
Sampling station编号
Serial number经度
Longitude纬度
Latitude拒马河源头
The source of Juma RiverS1 114°45′04.115″ 39°19′57.115″ 刁江汇
DiaojianghuiS2 115°01′36.775″ 39°25′39.702″ 紫荆关大桥
Zijingguan BridgeS3 115°10′03.338″ 39°25′41.776″ 清凉涧
QingliangjianS4 115°15′38.952″ 39°37′44.969″ 小丰口桥
Xiaofengkou BridgeS5 115°20′35.686″ 39°42′05.548″ 别岸
Bie’anS6 115°27′38.115″ 39°39′06.187″ 琅琊河
Langya RiverS7 115°29′18.773″ 39°39′08.849″ 天花板
TianhuabanS8 115°30′40.557″ 39°39′47.696″ 北石门
BeishimenS9 115°32′44.611″ 39°38′33.555″ 西河口
XihekouS10 115°34′09.356″ 39°38′27.367″ 九渡Jiudu S11 115°35′22.049″ 39°37′41.080″ 六渡Liudu S12 115°37′59.485″ 39°38′19.132″ 穆家口
MujiakouS13 115°39′48.970″ 39°37′43.089″ 千河口
QianhekouS14 115°39′57.071″ 39°36′38.520″ 龙安大桥
Longan BridgeS15 115°41′14.921″ 39°34′32.855″ 对鱼类资源调查的同时, 对15个采样点进行海拔(ASL)、水温(Tem)、水深(Dep)、溶解氧(DO)、透明度(SD)、浊度(FNU)、pH和叶绿素a(Chl.a)等环境因子的测定(水科院, 未发表数据)。
1.2 数据处理分析
采用以下多样性指数来分析拒马河鱼类群落多样性: (1)Margalef丰富度指数(DMa)[22]: DMa =(S–1)/ln N; (2)Shannon-Wiener多样性指数(H') [23]: H′= –∑Pi× ln Pi; (3)Pielou均匀度指数(J′) [24]: J′=H′/lnS。式中, S为采集到的鱼类种类数, N为采集到鱼类的个体数, Pi为样品中第i种鱼类的个体数占渔获物中全部个体数的比例。选择Pinkas等[25]提出的相对重要指数(Index of relative importance, IRI)计算拒马河鱼类群落优势种: IRI=(Ni+Wi) Fi。式中, Ni为渔获物中第i种鱼类的个体数占渔获物总个体数的百分比, Wi为第i种鱼类的生物量占渔获物总生物量的百分比, Fi为第i种鱼类在采样点出现的频率。定义IRI≥500的为优势种, 100≤IRI<500的为重要种, 10≤IRI<100的为常见种, 1≤IRI<10的为一般种, IRI<1的为稀有种[26]。
采用丰度生物量比较曲线(Abundance-biomass comparison curves, ABC曲线) [27]判断鱼类群落稳定性及受干扰程度, ABC曲线的统计量用W表示, 当生物量优势度曲线位于丰度优势度曲线之上时, W为正, 表明群落结构稳定未受干扰, 反之W为负[28]: W=∑(Bi–Ai)/50(S–1)。式中, Ai为第i种鱼类相对应的丰度累积百分比, Bi为第i种鱼类相对应的生物量累积百分比, S为物种数。
为满足数据的正态齐性和方差齐性, 分析前对鱼类丰度数据及多样性数据进行lg(x+1)对数转换, 运用SPSS 25.0通过单因素方差分析(One-way ANOVE)检验不同季节、海拔鱼类丰度及多样性的差异。若存在显著性差异, 进一步使用SNK(Student-newman-Keuls)多重比较分析不同季节、海拔间鱼类丰度及多样性的变化[33]。
在Primer 5.0软件中, 以采集到的鱼类丰度数据为原始矩阵, 进行lg(x+1)对数转换, 运用等级聚类分析(Cluster)和非参数多变量排序(NMDS)将鱼类群落划分不同的组分。运用相似性百分比分析(Similarity percentages, SIMPER)确定维持不同组分间结构相似性的关键物种[29]。
利用Canoco 5.0软件对15个采样点的鱼类物种组成和环境因子进行去趋势对应分析(Detrended correspondence analysis, DCA), 根据分析结果中各排序轴的大小选择线性模型(Redundancy analysis, RDA)或单峰模型(Canonical correspondence analysis, CCA)分析鱼类物种和环境因子的相关性[30]。如果排序轴大于4选择CCA分析, 排序轴小于3选择RDA分析, 介于3和4之间, 两种分析方法均可。
2. 结果
2.1 拒马河鱼类组成及优势种
2019—2021年在拒马河共采集鱼类5486尾, 隶属5目11科37种 (表 2)。渔获物中鲤形目占绝对优势, 有3科24种, 占总种数的64.86%; 鲈形目3科6种, 占16.22%; 鲇形目2科4种, 占10.81%; 合鳃鱼目2科2种, 占5.41%; 颌针鱼目1科1种, 占2.70%。
表 2 拒马河鱼类组成、各月份相对重要指数(IRI)及生态类型Table 2. The fish composition, index of relative important (IRI) of each month and ecological types in the Juma River物种Species 相对重要指数IRI 生态类型 2019.5 2019.8 2019.10 2020.8 2020.10 2021.5 Ecological types 鲤形目Cypriniformes 鲤科Cyprinidae 鲤亚科Cyprininae 鲫Carassius auratus 209.63 3024.43 371.53 1050.27 245.62 273.74 S, Omn, De, V 鱊亚科Acheilognathinae 兴凯鱊Acheilognathus chankaensis 8.36 — — — — — S, Omn, U,Sp 中华鳑鲏Rhodeus sinensis 8.08 — — 3.87 — — S, Omn, U, Sp 高体鳑鲏Rhodeus ocellatus 39.02 148.43 — 62.34 — 4.60 S, Omn, U, Sp 雅罗鱼亚科Leuciscinae 尖头鱥Rhynchocypris oxycephalus 668.20 216.15 101.76 476.61 44.59 90.97 R, Omn, Lo, Dr 拉氏鱥Rhynchocypris lagowskii 317.45 10.61 450.36 54.01 215.97 108.09 R, Omn, Lo, Dr 襁亚科Danioninae 宽鳍鱲Zacco platypus 1928.30 494.22 282.76 2497.27 1558.08 413.94 R, Omn, U, D 马口鱼Opsariichthys bidens 5.48 4.53 14.86 235.48 — — R, Car, U, D 亚科Gobioninae 麦穗鱼Pseudorasbora parva 2291.93 3463.52 1546.54 2742.93 2605.65 5382.96 S, Omn, De, V 点纹银Squalidus wolterstorffi 1162.26 121.92 24.68 92.90 1.49 22.05 S, Omn, Lo, Dr 中间银Squalidus intermedius — 53.15 — 34.37 — — R, Omn, Lo, Dr 兴隆山小鳔Microphysogobio hsinglungshanensis 217.79 53.69 29.31 — 12.42 369.75 R, Omn, Lo, Dr 黑鳍鳈Sarcochilichthys nigripinnis 548.01 622.04 198.44 685.56 383.17 884.47 S, Omn, Lo, Sp 棒花Gobio rivuloides 1.47 183.58 — — — 7.15 R, Omn, De, V 棒花鱼Abbottina rivularis 263.72 38.68 385.70 275.44 532.21 — R, Omn, De, V 蛇Saurogobio dabryi 0.82 1.83 26.13 40.43 337.43 46.09 R, Omn, Lo, Dr 花鳅科Cobitidae 泥鳅Misgurnus anguillicaudatus 1044.35 580.91 55.37 41.34 16.14 50.20 S, Omn, De, Dr 北方泥鳅Misgurnus bipartitus — 1.77 — — — — R, Omn, De, Dr 大鳞副泥鳅Paramisgurnus dabryanus 109.14 144.85 19.20 53.78 21.18 12.62 S, Omn, De, Dr 花斑花鳅Cobitis melanoleuca 95.38 115.57 — 12.07 1.72 71.63 S, Omn, De, Dr 条鳅科Nemacheilidae 北鳅Lefua costata — — — — 2.28 1.41 R, Omn, De, Dr 赛丽高原鳅Triplophysa sellaefer 11.33 — 5.83 — 104.93 37.93 R, Omn, De, Dr 尖头高原鳅TriplophysaCuneicephala — — 2.16 — — — R, Omn, De, Dr 达里湖高原鳅Triplophysa dalaica 97.01 12.11 80.15 — 168.26 127.34 R, Omn, De, Dr 合鳃鱼目Symbranchiformes 刺鳅科Mastacembelidae 刺鳅Sinobdella sinensis 50.12 26.09 — 59.92 — 2.85 S, Car, De, D 合鳃鱼科Symbranchidae 黄鳝Monopterus albus 12.24 — — — — — S, Car, De, Dr 鲈形目Percoidei 鰕虎鱼科Gobiidae 子陵吻鰕虎鱼Rhinogobius giurinus 2.65 8.25 4.56 53.91 38.20 5.10 R, Car, De, D 林氏吻鰕虎鱼Rhinogobius lindbergi 68.08 19.58 9.97 — 0.01 26.13 R, Car, De, D 波氏吻鰕虎鱼Rhinogobius cliffordpopei 44.63 0.96 1.70 — 1.31 — R, Car, De, D 福岛吻鰕虎鱼Rhinogobius fukushimai 30.47 0.63 0.43 1.83 — 2.40 R, Car, De, D 沙塘鳢科Odontobuidae 小黄䱂鱼Micropercops swinhonis 1250.14 308.40 786.30 453.05 386.69 663.59 S, Omn, De, D 丝足鲈科Osphronemidae 圆尾斗鱼Macropodus chinensis 1.40 — 0.44 0.98 1.42 2.62 S, Car, Lo, Dr 鲇形目Siluriformes 鲿科Bagridae 黄颡鱼Pelteobagrus fulvidraco 313.28 861.57 449.50 1265.35 191.14 10.27 S, Car, De, D 瓦氏黄颡鱼Pelteobagrus vachellii 7.92 37.05 0.58 — 9.54 — R, Car, De, D 乌苏里黄颡鱼Pelteobagrus ussuriensis — 287.03 — 37.59 — 20.41 R, Car, De, D 鲇科Siluridae 鲇Silurus asotus 10.83 203.88 198.89 26.22 — — S, Car, De, V 颌针鱼目Beloniformes 青鳉科Adrianichthyidae 青鳉Oryzias sinensis — 1.13 — — — — S, Omn, U, V 注: S. 喜缓流或静水; R. 亲流性; Car. 肉食性; Omn. 杂食性; U. 中上层; Lo. 中下层; De. 底栖。V. 黏性卵; D. 沉性卵; Dr. 漂流性卵; Sp. 喜贝类性卵Note: S. slow flow; R. riffle; Car. carnivore; Omn. omnivore; U. upper; Lo. lower; De. demersal; V. viscid egg; D. demersal egg; Dr. drifting egg; Sp. spawning in shellfish 鱼类生态类型(表 2): 按生活习性可以将采集到的鱼类划分为亲流性鱼类(R)和喜缓流或静水性鱼类(S), 其中亲流性鱼类有20种, 占采集到的鱼类总种数的54.05%, 喜缓流或静水性鱼类有17种, 占总种数的45.95%; 按食性可以划分为杂食性鱼类(Omn)和肉食性鱼类(Car), 杂食性鱼类有25种, 占总种数的67.57%, 肉食性鱼类有12种, 占总种数的32.43%; 按不同栖息水层划分, 底栖鱼类(De)有23种, 占62.16%, 中下层鱼类(Lo)有8种, 占21.62%, 中上层鱼类(Up)有6种, 占16.22%; 按产卵类型划分, 产漂流性卵的鱼类(Dr)有16种, 占43.24%, 产沉性卵的鱼类(D)有11种, 占29.73%, 产黏性卵的鱼类(V)6种, 占16.22%, 喜贝类性产卵的鱼类(Sp)4种, 占10.81%。
以相对重要指数IRI≥500的为优势种, 在6次采样调查中, 拒马河优势种有10种, 重要种有12种, 主要种有8种, 一般种有7种。各月份优势种存在差异, 2019年5月优势种为尖头鱥Rhynchocypris oxycephalus Sauvage、宽鳍鱲Zacco platypus Temminck & Schlegel、麦穗鱼Pseudorasbora parva Temminck & Schlegel、点纹银Squalidus wolterstorffi Regan、黑鳍鳈Sarcochilichthys nigripinnis Günther、泥鳅Misgurnus anguillicaudatus Cantor、小黄䱂鱼Micropercops swinhonis Günther; 2019年8月优势种为鲫Carassius auratus Linnaeus、麦穗鱼、黑鳍鳈、泥鳅、黄颡鱼Tachysurus fulvidraco Richardson; 2019年10月优势种为麦穗鱼、小黄䱂鱼; 2020年8月优势种为鲫、宽鳍鱲、麦穗鱼、黑鳍鳈、黄颡鱼; 2020年10月优势种为宽鳍鱲、麦穗鱼、棒花鱼Abbottina rivularis Basilewsky; 2021年5月优势种为麦穗鱼、黑鳍鳈、小黄䱂鱼。麦穗鱼在6次采样调查中均作为优势种出现。
2.2 拒马河鱼类群落特征
单因素方差分析显示(表 3), 不同季节间个体数(N)、物种数(S)、Shannon-Wiener多样性指数(H')、Margalef丰富度指数(DMa)呈现显著性差异(P<0.05), Pielou均匀度指数(J')无显著性差异(P>0.05)。SNK多重比较分析显示, 物种数、个体数均以2019年5月最多, 15个采样点平均捕获鱼类91.00尾9.67种, 2021年10月最少, 平均38.73尾5.07种; Shannon-Wiener多样性指数(H')、Margalef丰富度指数(DMa)均以2019年8月份最高, 分别是1.76(1.09—2.20)和2.06(1.12—2.98)。2019—2021年调查显示拒马河夏季的Shannon-Wiener多样性指数(H')、Margalef丰富度指数(DMa)和Pielou均匀度指数(J')均值高于春、秋两季(图 2)。
表 3 基于单因素方差分析检验(F值)拒马河鱼类多样性指数不同季节、不同海拔的变化Table 3. Changes of fish diversity index in different seasons and different altitudes in the Juma River based on One-way ANOVA test (F value)项目Item 个体数
Number of
individuals (N)物种数
Number of
species (S)多样性指数
Shannon-Wiener
diversity (H')均匀度指数
Pielou
evenness (J')丰富度指数
Margalef
richness (Dma)季节Season 2.935* 7.986** 6.504** 1.022ns 4.9** 海拔Altitude 12.206** 0.122ns 1.616ns 7.368ns 3.765ns 注: ns、*和**分别代表P>0.05、P<0.05、和P<0.01Note: ns, * and ** represent P>0.05, P<0.05 and P<0.01 ![]() 图 2 拒马河鱼类多样性指数在不同季节、不同海拔差异Figure 2. Differences of fish diversity index in different seasons and different altitudes in the Juma River
图 2 拒马河鱼类多样性指数在不同季节、不同海拔差异Figure 2. Differences of fish diversity index in different seasons and different altitudes in the Juma River物种数(S)、Shannon-Wiener多样性指数(H')、Margalef丰富度指数(DMa)、Pielou均匀度指数(J')在不同海拔无显著性差异(P>0.05), 只有个体数(N)呈现显著性差异(P<0.05), 海拔高于500 m的位点平均可采集鱼类143.91尾, 海拔低于500 m的位点可采集48.19尾(图 2)。
NMDS排序分析和聚类分析结果一致, 拒马河鱼类群落自上游至下游可划分为3组, 即S1和S2为组Ⅰ, S3和S4为组Ⅱ, 其余采样点为组Ⅲ(图 3)。相似性百分比分析SIMPER(表 4)显示, 组Ⅰ平均相似度为51.01%, 主要贡献种有麦穗鱼、鲫、小黄䱂鱼、泥鳅、拉氏鱥Rhynchocypris lagowskii Dybowski、赛丽高原鳅Triplophysa sellaefer Nichols、尖头鱥、达里湖高原鳅Triplophysa dalaica Kessler和林氏吻鰕虎鱼Rhinogobius lindbergi Berg, 累积贡献度为90.07%; 组Ⅱ平均相似度为66.92%, 主要贡献种有麦穗鱼、小黄䱂鱼、棒花鱼、黄颡鱼、鲫、子陵吻鰕虎鱼Rhinogobius giurinus Rutter、中间银Squalidus intermedius Nichols和林氏吻鰕虎鱼, 累积贡献度为90.16%; 组Ⅲ平均相似度为61.00%, 主要贡献种有麦穗鱼、宽鳍鱲、小黄䱂鱼、黑鳍鳈、鲫、黄颡鱼、泥鳅、棒花鱼、高体鳑鲏Rhodeus ocellatus Kner、兴隆山小鳔Microphysogobio hsinglungshanensis Mori、点纹银、刺鳅Sinobdella sinensis Bleeker和棒花Gobio rivuloides Nichols, 累积贡献度为91.23%。
![]() 图 3 拒马河鱼类群落Cluster聚类和NMDS排序图Figure 3. Clustering and NMDS sequencing analysis of fish community structure in the Juma River表 4 拒马河不同位点的主要贡献种相似性百分比分析Table 4. The main contributionspecies of Juma River in different locals of similarity percentage analysis
图 3 拒马河鱼类群落Cluster聚类和NMDS排序图Figure 3. Clustering and NMDS sequencing analysis of fish community structure in the Juma River表 4 拒马河不同位点的主要贡献种相似性百分比分析Table 4. The main contributionspecies of Juma River in different locals of similarity percentage analysis物种Species 组Ⅰ GroupⅠ 组Ⅱ GroupⅡ 组Ⅲ GroupⅢ AA AS (%) Con (%) AA AS (%) Con (%) AA AS (%) Con (%) 麦穗鱼Pseudorasbora parva 277.50 10.87 21.30 145.00 14.29 21.36 51.91 9.14 14.99 鲫Carassius auratus 73.00 8.74 17.14 9.00 6.23 9.31 13.09 5.24 8.59 小黄䱂鱼Micropercops swinhonis 24.50 6.56 12.86 41.00 10.97 16.40 31.91 7.28 11.93 泥鳅Misgurnus anguillicaudatus 23.50 6.00 11.76 9.09 3.63 5.95 拉氏鱥Rhynchocypris lagowskii 78.00 3.65 7.16 赛丽高原鳅Triplophysa sellaefer 17.50 2.82 5.54 尖头鱥Rhynchocypris oxycephalus 135.00 2.82 5.54 达里湖高原鳅Triplophysa dalaica 78.50 2.24 4.39 林氏吻鰕虎鱼Rhinogobius lindbergi 3.50 2.24 4.39 3.00 3.29 4.92 棒花鱼Abbottina rivularis 39.50 9.39 14.04 4.73 2.43 3.98 黄颡鱼Pelteobagrus fulvidraco 15.00 7.18 10.73 6.91 3.74 6.12 子陵吻鰕虎鱼Rhinogobius giurinus 8.50 4.82 7.20 中间银Squalidus intermedius 9.00 4.15 6.21 宽鳍鱲Zacco platypus 55.91 8.07 13.22 黑鳍鳈Sarcochilichthys nigripinnis 32.55 7.26 4.33 高体鳑鲏Rhodeus ocellatus 5.55 2.31 3.79 兴隆山小鳔Microphysogobio hsinglungshanensis 13.82 2.16 3.54 点纹银Squalidus wolterstorffi 18.27 2.11 3.45 刺鳅Sinobdella sinensis 2.45 1.21 1.99 棒花Gobio rivuloides 2.36 1.09 1.78 注: AA. 平均多度, AS. 平均相似度, Con. 贡献度Note: AA. average abund, AS. average similarity, Con. contribution 2.3 丰度生物量比较曲线
鱼类群落的优势度曲线变化趋势(图 4)显示, 2020年8月拒马河鱼类群落的生物量优势度曲线位于丰度优势度曲线之上, 统计值W>0, 表示该月份鱼类群落结构稳定未受干扰, 其余月份的丰度累积百分比与生物量累积百分比的大小交错分布, ABC曲线中丰度优势度曲线与生物量优势度曲线相交, 统计量W<0, 鱼类群落受到中等程度干扰。2020年夏季数量优势度曲线和丰度优势度曲线均排在前5位的优势种类为麦穗鱼、宽鳍鱲和尖头鱥, 该季度所采集渔获物中, 平均个体生物量为8.99 g, 平均个体生物量最大的是黄颡鱼, 为30.86 g, 平均个体生物量最小的是福岛吻鰕虎鱼Rhinogobius fukushimai Mori, 为0.50 g。
![]() 图 4 拒马河不同季节鱼类群落ABC曲线Figure 4. ABC curves of fish community in the Juma River in different seasons
图 4 拒马河不同季节鱼类群落ABC曲线Figure 4. ABC curves of fish community in the Juma River in different seasons2.4 拒马河鱼类群落结构与环境因子的关系
DCA排序结果显示四个排序轴长度均小于3(表 5), 其中一、二轴长分别为2.84和2.28, 累积解释物种变化率为13.74%和22.72%, 物种和环境与排序轴相关性为94.14%、87.37%, 拒马河鱼类群落参数对环境因子的响应呈线性关系, 因此拒马河鱼类群落与环境因子分析适用于RDA分析。
表 5 拒马河鱼类群落DCA分析Table 5. Detrended correspondence analysis of fish community in the Juma River排序轴编号No. Axis 1 Axis 2 Axis 3 Axis 4 特征值Eigenvalues 0.3306 0.2159 0.1402 0.0736 排序轴梯度长度Gradient length 2.84 2.28 2.13 1.81 物种累计解释变量百分比
Cumulative percentage explained variation of species (%)13.74 22.72 28.54 31.60 物种和环境因子与排序轴相关系数
Species-environment correlations0.9414 0.8737 0.8688 0.8913 RDA分析中第一轴和第二轴为主成分轴(表 6), 第一轴解释变化率为21.73%, 前两轴累计解释变化率为29.03%, 物种和环境同前两轴相关关系分别为94.66%和81.29%。拒马河鱼类群落与环境因子RDA排序图结果(图 5)显示, 影响拒马河鱼类群落结构组成影响最大的环境变量为海拔(ASL), 对主要种类的解释量为20.60%, 是显著影响因子(蒙特卡洛检验F=8.8, P=0.002), 海拔与pH(蒙特卡洛检验F=0.2, P=0.024)同第一轴的相关性较强。海拔对尖头高原鳅Triplophysa Cuneicephala Shaw & Tchang、达里湖高原鳅、尖头鱥、拉氏鱥、马口鱼Opsariichthys bidens Günther、蛇Saurogobio dabryi Bleeker、棒花鱼等影响较大。黑鳍鳈、刺鳅、瓦氏黄颡鱼Pelteobagrus vachellii Richardson等对pH的变化呈负相关, 而兴凯鱊Acanthorhodeus chankaensis Dybowski对pH的变化呈正相关。水温(蒙特卡洛检验F=2.8, P=0.008)与第二轴相关性较强, 对鲫、高体鳑鲏和乌苏里黄颡鱼Pelteobagrus ussuriensis Dybowski等的影响较大。
表 6 拒马河鱼类群落RDA分析Table 6. Redundancy analysis of fish community in the Juma River排序轴编号No. Axis 1 Axis 2 Axis 3 Axis 4 特征值Eigenvalues 0.217 0.073 0.047 0.034 累计解释变化率
Cumulative percentage explained variation (%)21.73 29.03 33.69 37.09 物种和环境因子与排序轴相关系数
Species-environment correlations0.9466 0.8129 0.7315 0.5583 累计解释拟合变化率
Explained fitted variation (cumulative)50.19 67.05 77.82 85.67 ![]() 图 5 拒马河鱼类与环境因子的RDA图Tem. 水温; Dep. 水深; DO. 溶解氧; Chl.a. 叶绿素a; SD. 透明度; ASL. 海拔; FNU. 浊度; PH. 酸碱度; sp1. 鲫; sp2. 兴凯鱊; sp3. 中华鳑鲏; sp4. 高体鳑鲏; sp5. 尖头鱥; sp6. 拉氏鱥; sp7. 宽鳍鱲; sp8. 马口鱼; sp9. 麦穗鱼; sp10. 点纹银; sp11. 中间银; sp12. 兴隆山小鳔; sp13. 黑鳍鳈; sp14. 棒花; sp15. 棒花鱼;sp16. 蛇; sp17. 泥鳅; sp18. 北方泥鳅; sp19. 大鳞副泥鳅; sp20. 花斑花鳅; sp21. 北鳅; sp22. 塞丽高原鳅; sp23. 尖头高原鳅; sp24. 达里湖高原鳅; sp25. 刺鳅; sp26. 黄鳝; sp27. 子陵吻鰕虎鱼; sp28. 林氏吻鰕虎鱼; sp29. 波氏吻鰕虎鱼; sp30. 福岛吻鰕虎鱼; sp31. 小黄䱂鱼; sp32. 圆尾斗鱼; sp33. 黄颡鱼; sp34. 瓦氏黄颡鱼; sp35. 乌苏里黄颡鱼; sp36. 鲇; sp37. 青鳉Figure 5. Redundancy analysis of fish community and environmental factors in the Juma RiverTem. temperature; Dep. depth; DO. dissolved oxygen; Chl.a. chlorophyll a; SD. transparency; ASL. altitude; FNU. formazin unit; pH. potential of hydrogen sp1. Carassius auratus; sp2. Acheilognathus chankaensis; sp3. Rhodeus sinensis; sp4. Rhodeus ocellatus; sp5. Rhynchocypris oxycephalus; sp6. Rhynchocypris lagowskii; sp7. Zacco platypus; sp8. Opsariichthys bidens; sp9. Pseudorasbora parva; sp10. Squalidus wolterstorffi; sp11. Squalidus intermedius; sp12. Microphysogobio hsinglungshanensis; sp13. Sarcochilichthys nigripinnis; sp14. Gobio rivuloides; sp15. Abbottina rivularis; sp16. Saurogobio dabryi; sp17. Misgurnus anguillicaudatus; sp18. Misgurnus bipartitus; sp19. Misgurnus dabryanus; sp20. Cobitis melanoleuca; sp21. Lefua costata; sp22. Triplophysa sellaefer; sp23. Triplophysa cuneicephala; sp24. Triplophysa dalaica; sp25. Sinobdella sinensis; sp26. Monopterus albus; sp27. Rhinogobius similis; sp28. Rhinogobius lindbergi; sp29. Rhinogobius cliffordpopei; sp30. Rhinogobius fukushimai; sp31. Micropercops swinhonis; sp32. Macropodus ocellatus; sp33. Tachysurus fulvidraco; sp34. Pelteobagrus vachellii; sp35. Pelteobagrus ussuriensis; sp36. Silurus asotus; sp37. Oryzias sinensis
图 5 拒马河鱼类与环境因子的RDA图Tem. 水温; Dep. 水深; DO. 溶解氧; Chl.a. 叶绿素a; SD. 透明度; ASL. 海拔; FNU. 浊度; PH. 酸碱度; sp1. 鲫; sp2. 兴凯鱊; sp3. 中华鳑鲏; sp4. 高体鳑鲏; sp5. 尖头鱥; sp6. 拉氏鱥; sp7. 宽鳍鱲; sp8. 马口鱼; sp9. 麦穗鱼; sp10. 点纹银; sp11. 中间银; sp12. 兴隆山小鳔; sp13. 黑鳍鳈; sp14. 棒花; sp15. 棒花鱼;sp16. 蛇; sp17. 泥鳅; sp18. 北方泥鳅; sp19. 大鳞副泥鳅; sp20. 花斑花鳅; sp21. 北鳅; sp22. 塞丽高原鳅; sp23. 尖头高原鳅; sp24. 达里湖高原鳅; sp25. 刺鳅; sp26. 黄鳝; sp27. 子陵吻鰕虎鱼; sp28. 林氏吻鰕虎鱼; sp29. 波氏吻鰕虎鱼; sp30. 福岛吻鰕虎鱼; sp31. 小黄䱂鱼; sp32. 圆尾斗鱼; sp33. 黄颡鱼; sp34. 瓦氏黄颡鱼; sp35. 乌苏里黄颡鱼; sp36. 鲇; sp37. 青鳉Figure 5. Redundancy analysis of fish community and environmental factors in the Juma RiverTem. temperature; Dep. depth; DO. dissolved oxygen; Chl.a. chlorophyll a; SD. transparency; ASL. altitude; FNU. formazin unit; pH. potential of hydrogen sp1. Carassius auratus; sp2. Acheilognathus chankaensis; sp3. Rhodeus sinensis; sp4. Rhodeus ocellatus; sp5. Rhynchocypris oxycephalus; sp6. Rhynchocypris lagowskii; sp7. Zacco platypus; sp8. Opsariichthys bidens; sp9. Pseudorasbora parva; sp10. Squalidus wolterstorffi; sp11. Squalidus intermedius; sp12. Microphysogobio hsinglungshanensis; sp13. Sarcochilichthys nigripinnis; sp14. Gobio rivuloides; sp15. Abbottina rivularis; sp16. Saurogobio dabryi; sp17. Misgurnus anguillicaudatus; sp18. Misgurnus bipartitus; sp19. Misgurnus dabryanus; sp20. Cobitis melanoleuca; sp21. Lefua costata; sp22. Triplophysa sellaefer; sp23. Triplophysa cuneicephala; sp24. Triplophysa dalaica; sp25. Sinobdella sinensis; sp26. Monopterus albus; sp27. Rhinogobius similis; sp28. Rhinogobius lindbergi; sp29. Rhinogobius cliffordpopei; sp30. Rhinogobius fukushimai; sp31. Micropercops swinhonis; sp32. Macropodus ocellatus; sp33. Tachysurus fulvidraco; sp34. Pelteobagrus vachellii; sp35. Pelteobagrus ussuriensis; sp36. Silurus asotus; sp37. Oryzias sinensis3. 讨论
3.1 拒马河鱼类群落组成结构特征
拒马河位于太行山迎风区北部, 北连西北山间盆地和北京西山地区, 西部以山西省为邻, 南部与大清河南支山区相接[31], 根据李国良[32]对河北淡水鱼类地理区系的划分, 拒马河流域大部分属于冀西山地, 海拔高度由500 m增至1000 m以上, 鱼类资源以条鳅科(赛丽高原鳅、达里湖高原鳅)、花鳅科(北方泥鳅Misgurnus bipartitus Dybowski、泥鳅)和鲤科中的雅罗鱼亚科(尖头鱥、拉氏鱥、马口鱼、宽鳍鱲)、亚科(麦穗鱼、点纹银、中间银)、鲃亚科(多鳞铲颌鱼Onychostoma macrolepis Bleeker)鱼类为主。根据历史记载, 拒马河鱼类资源并不丰富, 李国良[33]和王所安[20]记录了9种, 杨文波等[16]在2008年对拒马河北京段调查中监测到24种, 袁立来等[6]在2019—2020年对拒马河北京段调查中监测到33种。然而, 针对整个拒马河流域的调查, 近几十年未见报道。本研究在2019—2021年6次对拒马河鱼类资源调查中共发现37种, 与王鸿媛等[34]的记录相比减少了多鳞铲颌鱼、花䱻Hemibarbus maculatus Bleeker、黄线薄鳅Leptobotia flavolineata Wang、东方薄鳅Leptobotia orierntalis Xu, Fang & Wang和花斑副沙鳅Parabotia fasciata Dabry & Thiersant等, 这些鱼类对生态环境的改变极为敏感, 仅分布于水质清澈、溶氧量高和无污染的狭窄水域。综合之前调查[5, 6, 16], 这几种鱼类很可能已经在拒马河流域消失。受地理环境因素和人类活动的影响, 拒马河鱼类分布具有明显的区域性, 海拔在500 m以上的上游区域(组Ⅰ), 渔获物以花鳅科和条鳅科鱼类为主, 包括北鳅Lefua costata Kessler、达里湖高原鳅、赛丽高原鳅、尖头高原鳅、花斑花鳅Cobitis melanoleuca Nichols和北方泥鳅等; 海拔在200m以上的中游区域(组Ⅱ), 水生态环境受人类影响相对较小, 如S3和S4, 渔获物以棒花鱼、中间银为主要组成; 海拔在200 m以下的下游区域(组Ⅲ), 位于旅游景区, 水生态环境受到人类影响较大, 如自S5至S15, 渔获物以黑鳍鳈、麦穗鱼、小黄䱂鱼、高体鳑鲏和泥鳅等喜栖息小河流水环境具有一定耐受力的鱼类为主。组Ⅰ的平均相似性为51.01%, 小于组Ⅱ的66.92%和组Ⅲ的61.00%, 体现了拒马河上游地区相较于中下游地区更具生境多样性。
2019—2021年拒马河优势种有鲫、麦穗鱼、黑鳍鳈和宽鳍鱲等, 按照初次性成熟小于2龄, 最大体长小于24 cm的鱼类划分为小型鱼类的标准[35], 拒马河鱼类群落优势种几乎都是小型鱼类。从食性上看, 拒马河鱼类以杂食性为主, 马口鱼、黄颡鱼和刺鳅等偏肉食性鱼类较少, 表明拒马河鱼类群落高位营养级少, 水生态环境更适合杂食性鱼类的生存。生态类型方面, 亲流性的鱼类物种数较多, 符合内陆河流鱼类组成特点, 但优势种中喜缓流及静水的鱼类较多, 原因可能是拒马河拦河坝的修建导致河流片段化, 部分河段水流速度减慢甚至形成小型静水湖泊, 破坏了亲流性鱼类的捕食、产卵场所同时也为喜静水的鱼类提供了更合适的栖息环境所导致[9, 36, 37], 在设计河坝时, 应优先考虑设计过鱼通道及保护可能因河坝建设而受到影响的鱼类, 降低因河坝的建设而导致的水文环境的改变对其造成的影响。在此次鱼类资源调查中, 麦穗鱼在所有月份都作为鱼类群落的优势种出现, 具有较高的生态优势度。麦穗鱼适应能力强、繁殖力高、食性广, 同拒马河土著鱼类构成了竞争关系, 且吞食鱼卵, 干扰产卵场, 对土著鱼类的生存构成极大威胁[38]。对于麦穗鱼多的河段, 应定期监控其种群动态, 必要情况下可采取人为措施控制其种群数量。
多样性指数(H')和均匀度指数(J')是群落结构稳定的重要评价指标[39], 物种丰富度越高, 个体数分布越均匀, 群落结构越稳定, 多样性指数和均匀度指数也就较大, 反之, 物种受到环境的胁迫, 群落结构不稳定, 多样性指数则低。在本研究中, 拒马河鱼类群落夏季的多样性指数高于春、秋两季, 这可能与夏季水温上升鱼类的活动频率增加有关。鱼类群落结构及物种多样性可以衡量水体健康情况, 水生态环境恶化会直接影响鱼类的物种多样性, 拒马河鱼类群落的Shannon-Wiener多样性指数(H')、Margalef丰富度指数(DMa)及Pielou均匀度指数(J')均表明拒马河的水环境受到了一定程度的污染[40]。
3.2 拒马河鱼类群落稳定性
Warwick[41]于1986年提出丰度生物量比较曲线用于判断鱼类群落稳定性。ABC曲线将生物量优势度曲线和丰度优势度曲线放置在同一坐标系中, 通过两条曲线的分布来分析鱼类群落的受干扰程度[42]。ABC曲线基于r选择和k选择的传统进化理论, 群落结构稳定时, 群落以k选择种类(生长慢, 性成熟晚的大个体种类)为主要组成, 生物量优势度曲线位于丰度优势度曲线之上。随着干扰的增加, k选择的物种逐渐减少而r选择的物种(生长快, 性成熟早的小个体种类)逐渐增加, 丰度生物量比较曲线随之发生改变, 群落受到中度干扰时, 生物量优势度曲线与丰度优势度曲线相交; 群落受到严重干扰时, 生物量优势度曲线位于丰度优势度曲线之下[43]。根据李胜法等[43]的研究标准, 拒马河鱼类群落整体只有2020年夏季结构稳定未受干扰, 其他月份鱼类群落结构受到中等程度的干扰, 但在拒马河不同河段受干扰程度存在差异, 海拔500 m以上的两个位点, 平均采集鱼类143.91尾, 显著高于海拔低于500 m位点的48.19尾, 并采集到数量可观的尖头鱥、拉氏鱥、赛丽高原鳅等对水质要求高的鱼类, 可看出该河段鱼类群落受扰动程度较小。
3.3 拒马河鱼类群落结构与环境因子关系
拒马河鱼类多样性指数中个体数、物种数、多样性指数、丰富度指数在不同季节间呈现显著性差异, 表明拒马河鱼类群落主要受非生物因子的影响。Kadye等[44]研究证明, 温度、海拔、pH、溶解氧和距河口距离等是影响河流鱼类群落结构的主要环境因子。王卓等[45]在研究汉江平川段鱼类群落结构与环境因子的关系中发现电导率(Cond)、5日生化需氧量(BOD5)、pH和硫酸浓度是影响该江段鱼类群落结构的主要环境因子。本文通过RDA分析得出海拔是影响拒马河鱼类群落结构的主要环境因子, 对达里湖高原鳅、尖头高原鳅和北鳅等鳅科鱼类影响较大。水温作用于鱼类的分布、生长、繁殖和迁移, 对拒马河鱼类群落结构的影响仅次于海拔, 自拒马河源头至龙安大桥水温逐渐上升, 鲫和高体鳑鲏等温水性鱼类数量也逐渐增加。兴凯鱊与pH的变化呈正相关, 而棒花鱼、刺鳅和瓦氏黄颡鱼等同pH的变化呈负相关, pH会对鱼的摄食和生长产生影响, 也会影响鱼的感官、代谢、呼吸等生理过程[46]。溶解氧与波氏吻鰕虎鱼Rhinogobius cliffordpopei Nichols、福岛吻鰕虎鱼等小型肉食性鱼类相关性较强, 符合肉食性鱼类耗氧高的特点。叶绿素a也是影响拒马河鱼类群落结构的重要环境因子, 叶绿素a与浮游植物的密度密切相关, 对小黄䱂鱼和棒花鱼等杂食性鱼类影响较大。
4. 结论
本研究系统地调查了拒马河鱼类资源, 共发现鱼类37种, 隶属于5目11科, 补充了拒马河上游区域的物种组成和地理分布等缺乏的基本问题, 为拒马河鱼类资源开发利用和资源保护提供了基础数据。基于等级聚类分析(Cluster)和非参数多变量排序(NMDS)分析发现, 拒马河鱼类分布在空间上具有明显的区域性, 上游以适应清澈流水环境的花鳅科和条鳅科鱼类为主; 中上游以棒花鱼、中间银为主; 中下游以黑鳍鳈、麦穗鱼、小黄䱂鱼和泥鳅等具有一定耐受力的鱼类为主, 这对拒马河生态修复和水利工程修建能提供理论依据。通过群落优势种和丰度生物量比较曲线分析发现, 拒马河以耐污性较强的小型鱼类为主, 主要为鲫、麦穗鱼、黑鳍鳈和宽鳍鱲等, 鱼类群落结构受到一定程度的干扰; 冗余分析显示, 海拔是影响拒马河鱼类分布的主要环境因子。拒马河是大清河水系的主要河流, 目前已有“北京市房山区拒马河水生野生动物自然保护区”, 建议进一步根据拒马河独特的水生态环境划定生态保护红线, 加强对该流域生物多样性的保护。
附表 S1 拒马河鱼类分布Appendix S1. Fish species distribution in Juma River物种
Species采样地点Sampling site 拒马河
源头刁江
汇紫荆关
大桥清凉
涧小丰
口桥别岸 琅琊
河天花
板北石
门西河
口九渡 六渡 穆家
口千河
口龙安
大桥鲤形目Cypriniformes 鲤科Cyprinidae 鲤亚科Cyprininae 鲫Carassius auratus + + + + + + + + + + + + + + + 鱊亚科AcheilognatSpnae 兴凯鱊Acanthorhodeus chankaensis + + 中华鳑鲏Rhodeus sinensis + + + 高体鳑鲏Rhodeus ocellatus + + + + + + + + + + 雅罗鱼亚科Leuciscinae 尖头鱥Rhynchocypris oxycephalus + + + + + + + + + 拉氏鱥Rhynchocypris lagowskii + + + + + + + 襁亚科Danioninae 宽鳍鱲Zacco platypus + + + + + + + + + + + + 马口鱼Opsariichthys bidens + + + + + + 亚科Gobioninae 麦穗鱼Pseudorasbora parva + + + + + + + + + + + + + + + 点纹银Squalidus wolterstorffi + + + + + + + + + + 中间银Squalidus intermedius + + + + + 兴隆山小鳔Microphysogobio hsinglungshanensis + + + + + + + + + 黑鳍鳈Sarcochilichthys nigripinnis + + + + + + + + + + + 棒花Gobio rivuloides + + + + + + + + + + 棒花鱼Abbottina rivularis + + + + + + + + + + + + + 蛇Saurogobio dabryi + + + + 花鳅科Cobitidae 泥鳅Misgurnus anguillicaudatus + + + + + + + + + + + + + + 北方泥鳅Misgurnus bipartitus + 大鳞副泥鳅Paramisgurnus dabryanus + + + + + + 花斑花鳅Cobitis melanoleuca + + + + 条鳅科Nemacheilidae 北鳅Lefua costata + 赛丽高原鳅Triplophysa sellaefer + + + + 尖头高原鳅Triplophysacuneicephala + 达里湖高原鳅Triplophysa dalaica + + + + 合鳃鱼目SymbrancSpformes 刺鳅科Mastacembelidae 刺鳅Sinobdella sinensis + + + + + + + 合鳃鱼科Symbranchidae 黄鳝Monopterus albus + 鲈形目Percoidei 鰕虎鱼科Gobiidae 子陵吻鰕虎鱼Rhinogobius giurinus + + + + 林氏吻鰕虎鱼Rhinogobius lindbergi + + + + + + + + + 波士吻鰕虎鱼Rhinogobius cliffordpopei + + + + + + + 福岛吻鰕虎鱼Rhinogobius fukushimai + + + + + + + 沙塘鳢科Odontobuidae 小黄䱂鱼Micropercops swinhonis + + + + + + + + + + + + + + + 丝足鲈科Osphronemidae 圆尾斗鱼Macropodus chinensis + + + 鲇形目Siluriformes 鲿科Bagridae 黄颡鱼Pelteobagrus fulvidraco + + + + + + + + + + + + + 瓦氏黄颡鱼Pelteobagrus vachellii + + + + 乌苏里黄颡鱼Pelteobagrus ussuriensis + + + + + 鲇科Siluridae 鲇Silurus asotus + + + + + + + 颌针鱼Beloniformes 青鳉科Adrianichthyidae 青鳉Oryzias sinensis + 注: “+”表示现场调查采集到样本Note: “+” represents collected fish species in the surveys -
![]()
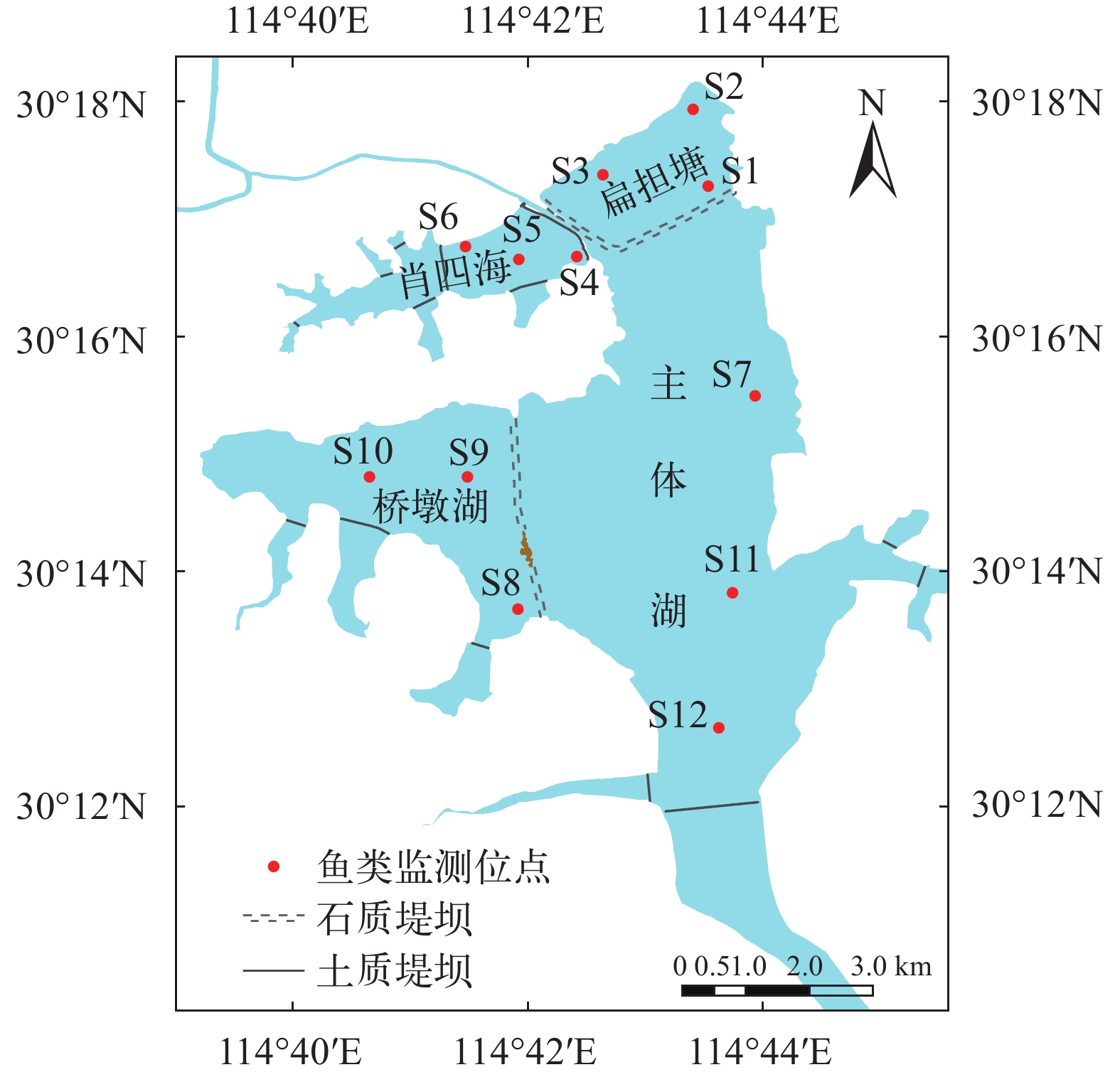
图 1 2019—2020年保安湖调查区域与样点分布
Figure 1. Study area and sampling sites in Bao’an Lake during 2019—2020
![]()
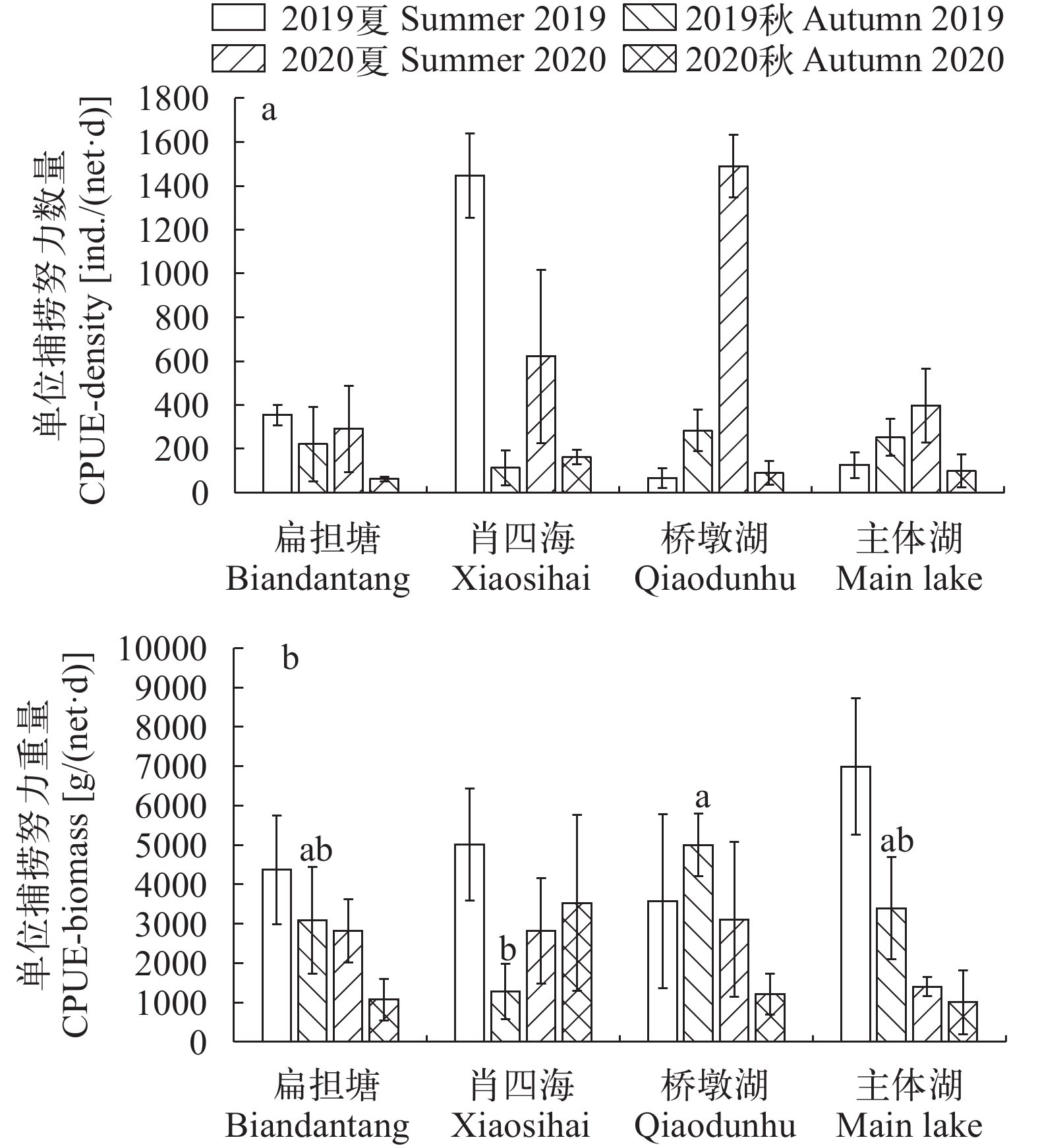
图 2 保安湖不同湖区鱼类单位捕捞努力量
a. 数量, b. 重量
Figure 2. Catch per unit effort of fish
a. density and b. biomass in sub-lakes of Bao’an Lake
![]()
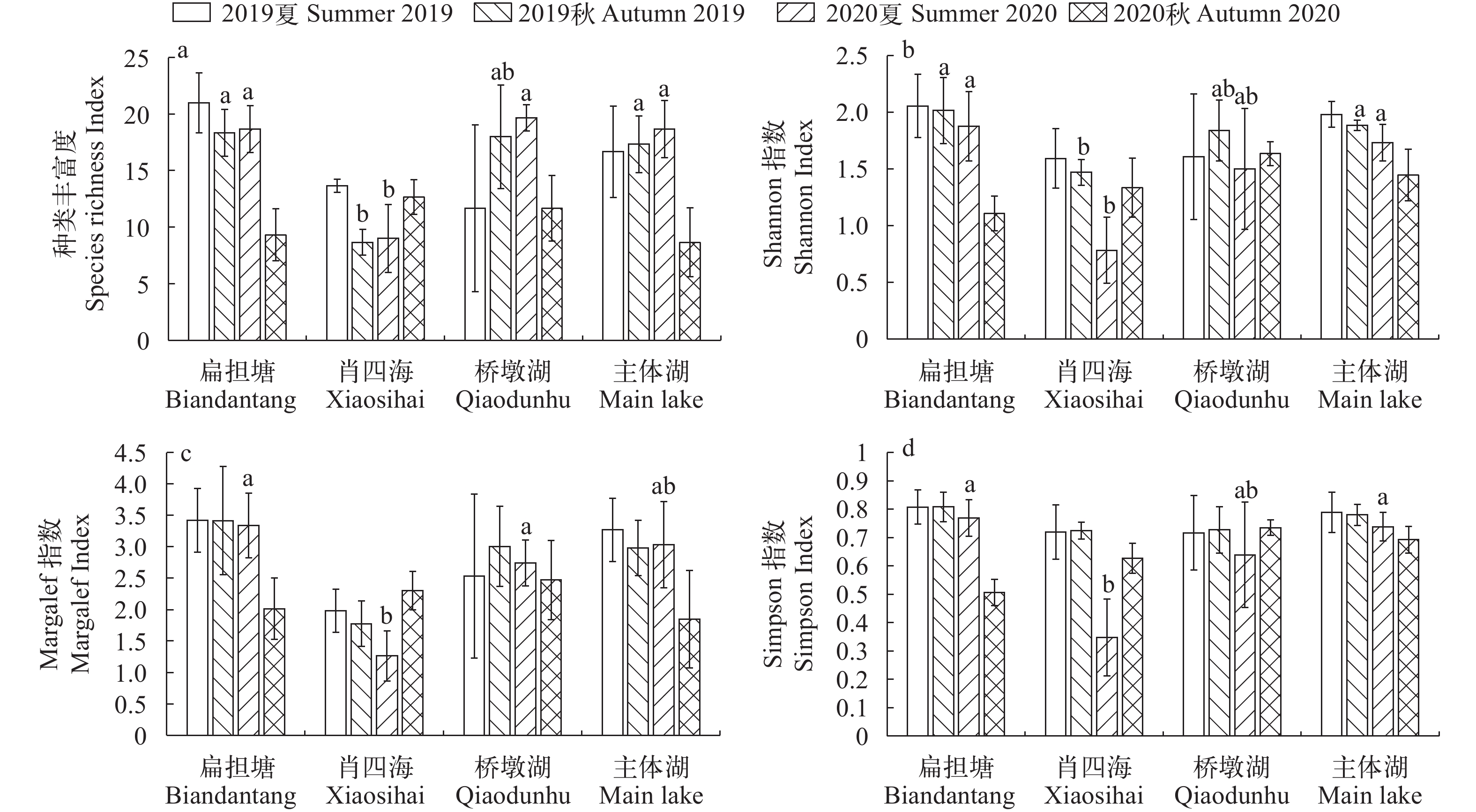
图 3 保安湖不同湖区鱼类物种多样性指数
a. 种类丰富度指数; b. Shannon多样性指数; c. Margalef丰富度指数; d. Simpson优势度指数。柱状图上边缘不同小写字母表示湖区间有显著性差异; 下同
Figure 3. Fish species diversity in sub-lakes of Bao’an Lake
a. index of species richness; b. Shannon index; c. Margalef index; d. Simpson index. Different lowercase letters on the upper of histograms indicate statistically significant differences between sub-lakes. The same applies below
![]()
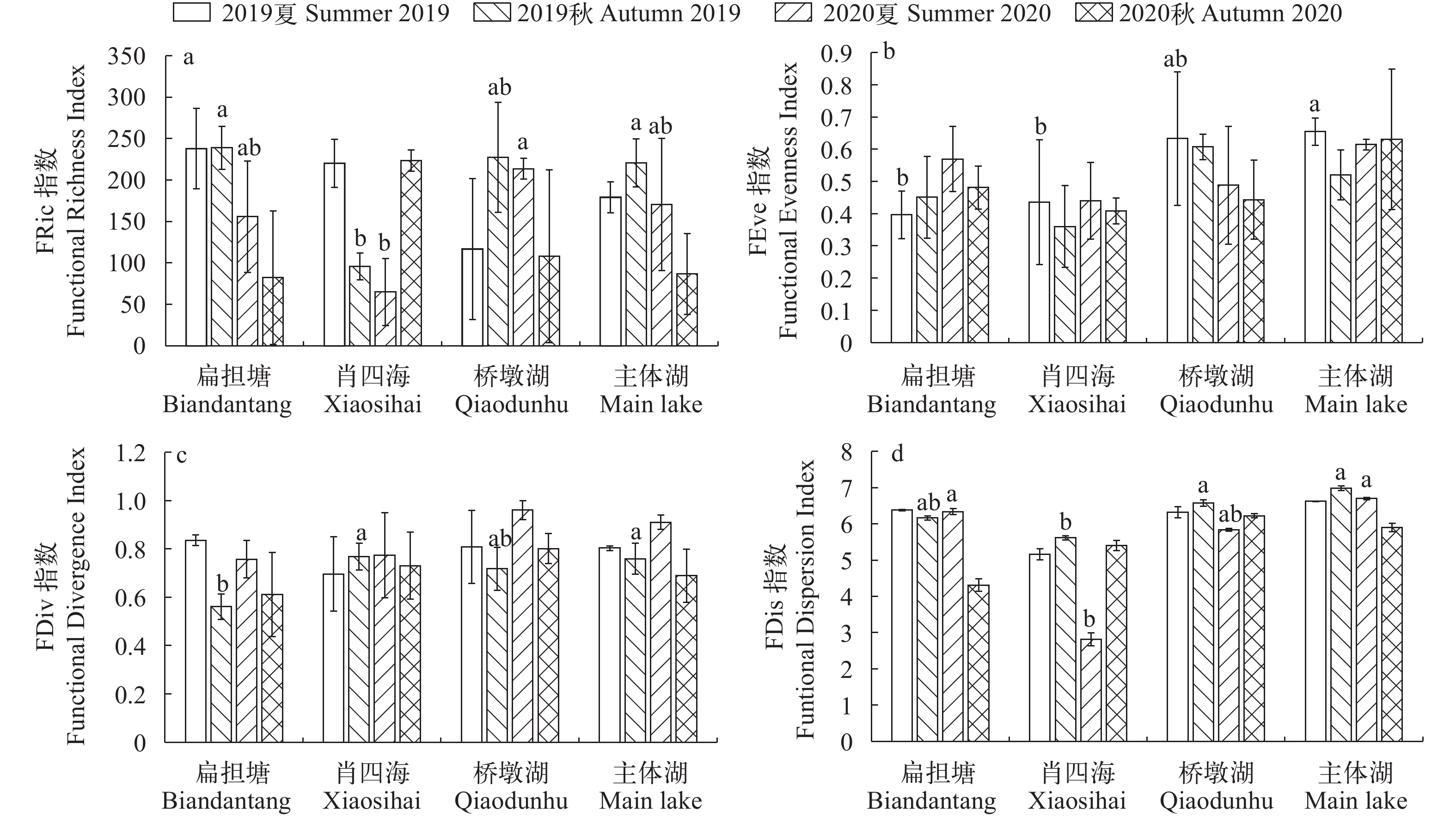
图 4 保安湖不同湖区鱼类功能多样性指数
a. 功能丰富度指数; b. 功能均匀度指数; c. 功能趋异度指数; d. 功能均匀度指数
Figure 4. Fish functional diversity in sub-lakes of Bao’an Lake
a. functional richness index; b. functional evenness index; c. functional divergence index; d. functional dispersion index
![]()
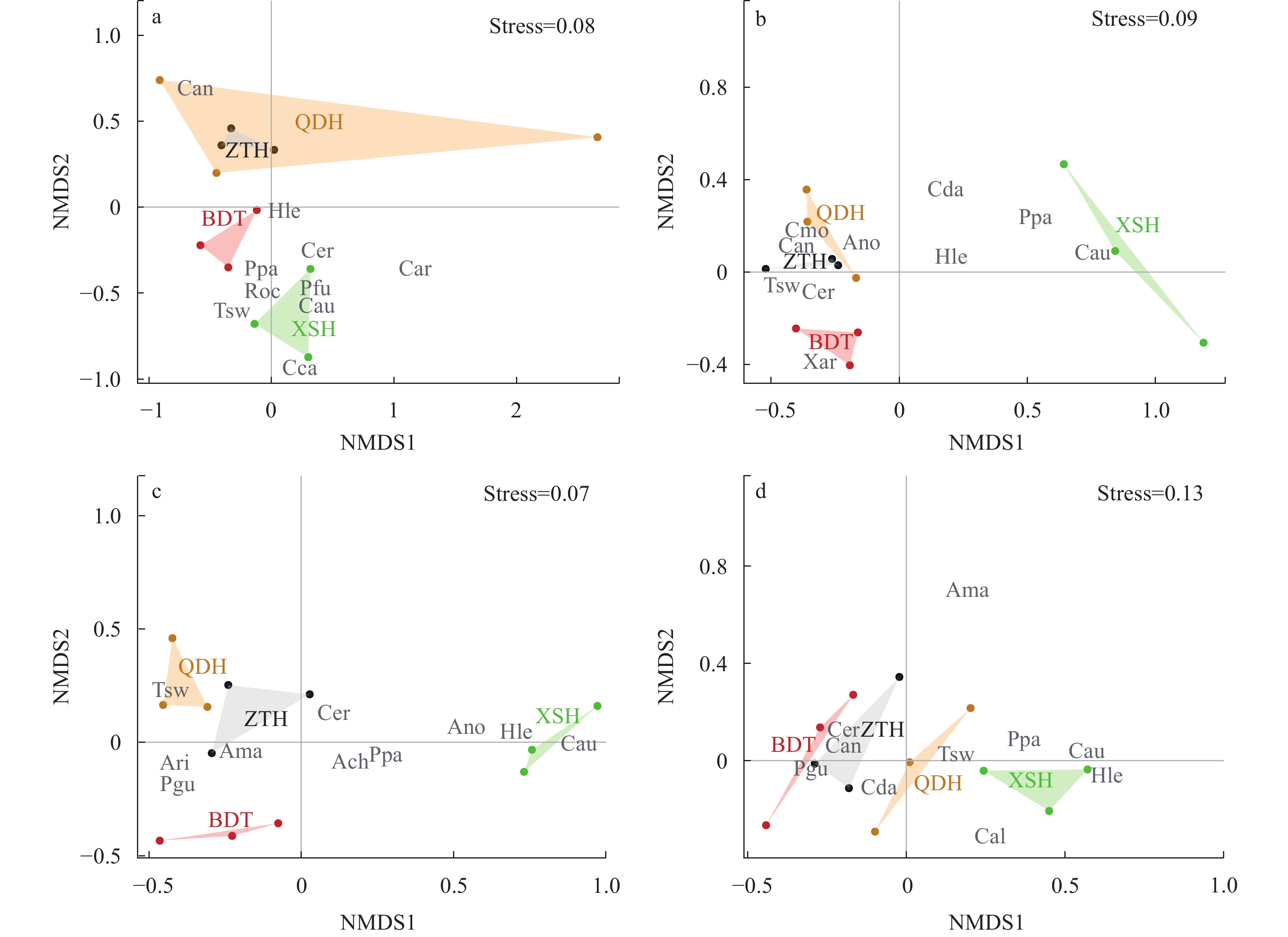
图 5 保安湖不同湖区鱼类群落NMDS分析
a. 2019夏季; b. 2019秋季; c. 2020夏季; d. 2020秋季
Figure 5. Non-metric multidimensional scaling analysis of fish communities among sub—lakes of Bao’an Lake
a. summer 2019; b. autumn 2019; c. summer 2020; d. autumn 2020
表 1 鱼类功能性状及性状分类
Table 1 Fish functional traits and categories
类型
Type性状组成
Traits性状分类
Trait categories类型
Type性状组成
Traits性状分类
Trait categories生长 栖息地偏好 山溪河流 摄食 食性 无脊椎动物食性 湖泊定居 浮游生物食性 江湖洄游 杂食性 河海洄游 植食性 栖息水层 表层 鱼食性 中上层 口型 上位口 中下层 端位口 底层 亚下位 体型 鳗型 下位口 圆筒型 营养级 纺锤型 其他 群落恢复力稳定性 侧扁型 群落抵抗力稳定性 平扁型 系统发育多样性指数 最大体长 土著/外来 体宽 易损率 性成熟体长 尾柄长 尾柄宽 眼间距 繁殖 产卵类型 沉性卵 黏性卵 浮性卵 漂流性卵 相对繁殖力 绝对繁殖力 卵径 护幼行为  下载: 导出CSV
下载: 导出CSV
表 2 保安湖历史记录和现状各湖区鱼类种类名录
Table 2 List of fish species in historical and current records in different sub-lakes in Bao’an Lake
种类
Species扁担塘
Biandantang肖四海
Xiaosihai桥墩湖
Qiaodunhu主体湖
Main Lake现状
Current全湖历史
History鳀科Engraulidae 1.短颌鲚Coilia brachygnathus + + + + + 银鱼科Salangidae 2.寡齿新银鱼Neosalanx oligodontis + + + + + 鲤科Cyprinidae 3.鲤Cyprinus carpio + + + + + 4.鲫Carassius auratus auratus + + + + + + 5.棒花鱼Abbottina rivularis + + + + + + 6.麦穗鱼Pseudorasbora parva + + + + + + 7.黑鳍鳈Sarcocheilichthys nigripinnis + + + + + 8.华鳈S. sinensis + + + 9.唇䱻Hemibarbus labeo + 10.花䱻H. maculatus + + + + 11.蛇 Saurogobio dabryi + + 12.亮银Squalidus nitens + + + + 13.西湖银 S. sihuensis + 14.似刺鳊Paracanthobrama guichenoti + + + + + 15.草鱼Ctenopharyngodon idellus + + + + + + 16.青鱼Mylopharyngodon piceus + + + + + 17.鳡Elopichthys bambusa + 18.鳤Ochetobius elongatus + 19.赤眼鳟Squaliobarbus curriculus + + + + 20.短须鱊Acheilognathus barbatulus + 21.兴凯鱊A. chankaensis + + + + + + 22.大鳍鱊A. macropterus + + + + + + 23.越南鱊A. tonkinensis + 24.彩副鱊Paracheilognathus imberbis + 25.方氏鳑鲏Rhodeus fangi + 26.彩石鳑鲏R. lighti + 27.高体鳑鲏R. ocellatus + + + + + 28.中华鳑鲏R.sinensis + 29.似鱎Toxabramis swinhonis + + + + + + 30.贝氏䱗Hemiculter bleekeri + 31.䱗H. leucisculus + + + + + + 32.飘鱼Pseudolaubuca sinensis + 33.翘嘴鲌Culter alburnus + + + + + + 34.达氏鲌C. dabryi + + + + + + 35.蒙古鲌C. mongolicus + + + + + 36.尖头鲌C. oxycephalus + + + + 37.红鳍原鲌Cultrichthys erythropterus + + + + + + 38.团头鲂Megalobrama amblycephala + 39.鲂M. skolkovii + 40.三角鲂 M. terminalis + 41.鳊Parabramis pekinensis + + + 42.似鳊Pseudobrama simoni + + + + + + 43.银鲴Xenocypris argentea + + + + + 44.黄尾鲴X. davidi + + + + + + 45.细鳞鲴X. microlepis + 46.鲢Hypophthalmichthys molitrix + + + + + + 47.鳙Hypophthalmichthys nobilis + + + + + + 鳅科Cobitidae 48.泥鳅Misgurnus anguillicaudatus + + + + 49.大鳞副泥鳅Paramisgurnus dabryanus + + + 50.中华沙鳅 Botia superciliaris + 51.花鳅 Cobitis sinensis + 鲿科Bagridae 52.黄颡鱼Pelteobagrus fulvidraco + + + + + + 53.瓦氏黄颡鱼P. vachelli + + + 54.长须黄颡鱼P. eupogon + + 鲇科Siluridae 55.鲇Silurus asotus + + + 青鳉科Oryziatidae 56.青鳉 Oryzias latipes + 57.中华青鳉O. latipes sinensis + 鱵科Hemirhamphidae 58.间下鱵Hyporhamphus intermedius + + + + + 合鳃鱼科Synbranchidae 59.黄鳝Monopterus albus + 斗鱼科Belontiidae 60.圆尾斗鱼Macropodus chinensis + + + 鳢科Channidae 61.乌鳢Channa argus + + + + + 鮨科Serranidae 62.鳜Siniperca chuatsi + + + + + 63.大眼鳜S. kneri + 塘鳢科Eleotridae 64.黄䱂鱼Hypseleotris swinhonis + 65.沙塘鳢Odontobutis potamophila + + + 66.小黄䱂鱼 Micropercops swinhonis + + + + 鰕虎鱼科Gobiidae 67.波氏吻鰕虎鱼Rhinogobius cliffordpopei + 68.子陵吻鰕虎鱼R. giurinus + + + + + 69.粘皮鲻鰕虎鱼Mugilogobius myxodermus + 鳗鲡科Anguillidae 70.鳗鲡Anguilla japonica + 刺鳅科Mastacembelidae 71.刺鳅Mastacembelus aculeatus + 72.中华刺鳅Sinobdella sinensis + 总计Total 32 25 35 33 44 68 注: 全湖鱼类历史记录参考胡传林和黄祥飞[32]、梁彦龄和刘伙泉[33]、张堂林[34]、Xie 等[35]、李为[36]、Li 等 [37, 38]Note: Historical records of fish in Bao’an Lake are from Hu and Huang[32], Liang and Liu[33], Zhang[34], Xie, et al.[35], Li[36], Li, et al.[37, 38]
下载: 导出CSV
表 3 保安湖不同湖区前五位鱼类优势种
Table 3 Top five dominant species of fish in sub-lakes of Bao’an Lake
湖区
Sub-lake2019夏
Summer 20192019秋
Autumn 20192020夏
Summer 20202020秋
Autumn 2020扁担塘
Biandantang高体鳑鲏、鳙、红鳍原鲌、
麦穗鱼、草鱼短颌鲚、银鲴、䱗、
红鳍原鲌、似鱎似鱎、鲫、似刺鳊、
大鳍鱊、红鳍原鲌短颌鲚、达氏鲌、
似鱎、红鳍原鲌肖四海
Xiaosihai鲫、红鳍原鲌、䱗、鳙、鲤 达氏鲌、麦穗鱼、䱗、
鲫、鳙鲫、鳙、䱗、大鳍鱊 似鱎、䱗、草鱼、
达氏鲌、鲫桥墩湖
Qiaodunhu红鳍原鲌、短颌鲚、鳙、䱗 短颌鲚、蒙古鲌、鲢、
似鳊、鳙似鱎、红鳍原鲌、鲫、
大鳍鱊、䱗短颌鲚、似鱎、达氏鲌、
红鳍原鲌主体湖
Main lake草鱼、䱗、短颌鲚、
高体鳑鲏、鳙短颌鲚、似鱎、䱗、
红鳍原鲌、鳙似鱎、红鳍原鲌,似刺鳊、
鲫、大鳍鱊短颌鲚、达氏鲌、
似鱎、红鳍原鲌注: 以相对重要性指数大小进行鱼类优势种排列顺序Note: The order of the dominant fish species is based on the index of relative importance
下载: 导出CSV
表 4 保安湖不同湖区鱼类群落差异的主要种类组成(贡献率>5%)
Table 4 Main species composition of fish community differences in sub-lakes of Bao’an Lake (contribution rate>5%)
湖区Sub-lake 2019夏Summer 2019 2019秋Autumn 2019 2020夏Summer 2020 2020秋Autumn 2020 扁担塘&
肖四海鲤*、鲫、高体鳑鲏、黄颡鱼、似鱎、䱗、兴凯鱊 银鲴*、红鳍原鲌*、短颌鲚、似鱎*、鲫*、麦穗
鱼*、青鱼、达氏鲌似鱎、鲫、大鳍鱊、棒花鱼*、似刺鳊*、鳙、
䱗*、麦穗鱼䱗*、似鱎*、短颌鲚、
鲫*、麦穗鱼、翘嘴鲌、
红鳍原鲌扁担塘&
桥墩湖高体鳑鲏*、麦穗鱼*、
似鱎*、红鳍原鲌、鲫、
兴凯鱊似鳊*、银鲴、短颌鲚、
蒙古鲌、鳙红鳍原鲌*、似鱎、鲫、大鳍鱊、间下鱵、兴凯鱊 红鳍原鲌、似鱎、短颌鲚、达氏鲌、䱗、翘嘴鲌、麦穗鱼、大鳍鱊 扁担塘&
主体湖高体鳑鲏*、似鱎*、麦穗鱼、䱗、鲫、兴凯鱊 鳙、短颌鲚、银鲴、似鱎、大鳍鱊 红鳍原鲌、似鱎、短颌
鲚*、大鳍鱊、鲫、麦穗鱼短颌鲚、达氏鲌、似鱎、麦穗鱼、翘嘴鲌、红鳍原鲌、䱗、似刺鳊* 肖四海&
桥墩湖鲤*、鲫*、红鳍原鲌*、麦穗鱼*、䱗、高体鳑鲏、黄颡鱼*、短颌鲚 短颌鲚*、蒙古鲌*、
似鳊*、红鳍原鲌、鲫*、似鱎、达氏鲌、麦穗鱼似鱎*、鲫、大鳍鱊、棒花鱼*、鳙、红鳍原鲌 䱗、红鳍原鲌、似鱎、短颌鲚、鲫、麦穗鱼、达氏鲌 肖四海&
主体湖鲤*、鲫*、短颌鲚、高体鳑鲏、䱗、黄颡鱼*、
银鲴*、乌鳢*、麦穗鱼短颌鲚*、似鱎*、红鳍原鲌*、鳙*、青鱼*、鲫、
达氏鲌、麦穗鱼似鱎*、鲫*、大鳍鱊*、似刺鳊*、短颌鲚*、棒花鱼 䱗*、短颌鲚、似鱎、麦穗鱼、达氏鲌、红鳍原鲌、鲫* 桥墩湖&
主体湖䱗、银鲴*、红鳍原鲌、草鱼*、短颌鲚、兴凯鱊、鲫 似鳊*、蒙古鲌、鳙、似鱎、达氏鲌、翘嘴鲌、䱗、 银鲴 大鳍鱊、麦穗鱼、 鲫、鳙、兴凯鱊、间下鱵、似刺鳊、鲤 红鳍原鲌、达氏鲌、短颌鲚、翘嘴鲌、大鳍鱊、似鱎、䱗、麦穗鱼 注: 种类排序以对群落差异的贡献性大小、*代表为显著性物种Note: Species are sorted by the contribution to community differences、*represents significant species
下载: 导出CSV
-
[1] Poff N L, Allan J D, Bain M B, et al. The natural flow regime [J]. BioScience, 1997, 47(11): 769-784. doi: 10.2307/1313099
[2] Bunn S E, Arthington A H. Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity [J]. Environmental Management, 2002, 30(4): 492-507. doi: 10.1007/s00267-002-2737-0
[3] Lytle D A, Poff N L. Adaptation to natural flow regimes [J]. Trends in Ecology & Evolution, 2004, 19(2): 94-100.
[4] Olden J D, Konrad C P, Melis T S, et al. Are large-scale flow experiments informing the science and management of freshwater ecosystems [J]? Frontiers in Ecology and the Environment, 2014, 12(3): 176-185. doi: 10.1890/130076
[5] 王洪铸, 刘学勤, 王海军. 长江河流-泛滥平原生态系统面临的威胁与整体保护对策 [J]. 水生生物学报, 2019, 43(S1): 157-182. doi: 10.7541/2019.178 Wang H Z, Liu X Q, Wang H J. The Yangtze River-floodplain ecosystem: multiple threats and holistic conservation [J]. Acta Hydrobiologica Sinica, 2019, 43(S1): 157-182. doi: 10.7541/2019.178
[6] Ward J V, Tockner K, Arscott D B, et al. Riverine landscape diversity [J]. Freshwater Biology, 2002, 47(4): 517-539. doi: 10.1046/j.1365-2427.2002.00893.x
[7] Amoros C, Bornette G. Connectivity and biocomplexity in waterbodies of riverine floodplains [J]. Freshwater Biology, 2002, 47(4): 761-776. doi: 10.1046/j.1365-2427.2002.00905.x
[8] Pan B Z, Wang H J, Liang X M, et al. Macrozoobenthos in Yangtze floodplain lakes: patterns of density, biomass, and production in relation to river connectivity [J]. Journal of the North American Benthological Society, 2011, 30(2): 589-602. doi: 10.1899/10-025.1
[9] Zhang X K, Liu X Q, Wang H Z. Developing water level regulation strategies for macrophytes restoration of a large river-disconnected lake, China [J]. Ecological Engineering, 2014(68): 25-31.
[10] 王利民, 胡慧建, 王丁. 江湖阻隔对涨渡湖区鱼类资源的生态影响 [J]. 长江流域资源与环境, 2005, 14(3): 287-292. doi: 10.3969/j.issn.1004-8227.2005.03.005 Wang L M, Hu H J, Wang D. Ecological impacts of disconnection from the Yangtze on fish resources in Zhangdu Lake [J]. Resources and Environment in the Yangtze Basin, 2005, 14(3): 287-292. doi: 10.3969/j.issn.1004-8227.2005.03.005
[11] 屈霄, 郭传波, 熊芳园, 等. 梯级开发下金沙江下游鱼类群落结构特征及环境驱动因子 [J]. 水生态学杂志, 2020, 41(6): 46-56. doi: 10.15928/j.1674-3075.2020.06.006 Qu X, Guo C B, Xiong F Y, et al. Characterization of the fish community and environmental driving factors during development of cascading dams in the lower Jinsha River [J]. Journal of Hydroecology, 2020, 41(6): 46-56. doi: 10.15928/j.1674-3075.2020.06.006
[12] 王洪铸, 王海军. 长江中下游江湖阻隔的生态学效应及修复方略 [A]//王兆印, 邵东国, 邵学军, 等. 长江流域水沙生态综合管理 [M]. 北京: 科学出版社, 2009: 379-396. Wang H Z, Wang H J. Ecological Effects of River-lake Disconnection and Restoration Strategies in the Mid-lower Yangtze Basin [A]//Wang Z Y, Shao D G, Shao X J, et al (Eds.), Comprehensive ecological management of water and sediment in the Yangtze River Basin. Beijing: Science Press, 2009: 379-396.
[13] Liu X Q, Wang H Z. Estimation of minimum area requirement of river-connected lakes for fish diversity conservation in the Yangtze River floodplain [J]. Diversity and Distributions, 2010, 16(6): 932-940. doi: 10.1111/j.1472-4642.2010.00706.x
[14] Liu X Q, Wang H Z. Food web of benthic macroinvertebrates in a large Yangtze River-connected lake: the role of flood disturbance [J]. Fundamental and Applied Limnology, 2008, 171(4): 297-309. doi: 10.1127/1863-9135/2008/0171-0297
[15] Olson D M, Dinerstein E. The global 200: a representation approach to conserving the earth’s most biologically valuable ecoregions [J]. Conservation Biology, 1998, 12(3): 502-515. doi: 10.1046/j.1523-1739.1998.012003502.x
[16] Chen Y S, Zhang S H, Huang D, et al. The development of China’s Yangtze River Economic Belt: how to make it in a green way [J]? Science Bulletin, 2017, 62(9): 648-651. doi: 10.1016/j.scib.2017.04.009
[17] Chen Y S, Qu X, Xiong F Y, et al. Challenges to saving China’s freshwater biodiversity: fishery exploitation and landscape pressures [J]. Ambio, 2020, 49(4): 926-938. doi: 10.1007/s13280-019-01246-2
[18] 陈宇顺. 多重人类干扰下长江流域的水生态系统健康修复 [J]. 人民长江, 2019, 50(2): 19-23. doi: 10.16232/j.cnki.1001-4179.2019.02.004 Chen Y S. Restoring aquatic ecosystem health of Yangtze River Basin under multiple human disturbances [J]. Yangtze River, 2019, 50(2): 19-23. doi: 10.16232/j.cnki.1001-4179.2019.02.004
[19] Appelberg M, Berger H M, Hesthagen T, et al. Development and intercalibration of methods in Nordic freshwater fish monitoring [J]. Water, Air, and Soil Pollution, 1995, 85(2): 401-406. doi: 10.1007/BF00476862
[20] 汪振华, 龚甫贤, 吴祖立, 等. 多网目单层和三重组合刺网在岩礁和沙地生境中的鱼类采集效果比较 [J]. 生态学杂志, 2013, 32(2): 507-514. doi: 10.13292/j.1000-4890.2013.0151 Wang Z H, Gong F X, Wu Z L, et al. Efficiency of multi-mesh gillnets and multi-mesh trammel nets in collecting fish from rocky reef and sandy beach habitats [J]. Chinese Journal of Ecology, 2013, 32(2): 507-514. doi: 10.13292/j.1000-4890.2013.0151
[21] Guo C, Wang R, Qu X, et al. Assessing fish assemblages in a shallow Yangtze River lake using multi-mesh gillnets and weirs [J]. Acta Hydrobiologica Sinica, 2018, 42(6): 44-51.
[22] Shannon, C E. A mathematical theory of communication [J]. Bell System Technical Journal, 1948, 27(3): 379-423. doi: 10.1002/j.1538-7305.1948.tb01338.x
[23] Margalef R. Perspectives in Ecological Theory [M]. Chicago: University of Chicago Press, 1968.
[24] Simpson E H. Measurement of diversity [J]. Nature, 1949, 163(4148): 688. doi: 10.1038/163688a0
[25] Villéger S, Brosse S, Mouchet M, et al. Functional ecology of fish: current approaches and future challenges [J]. Aquatic Sciences, 2017, 79(4): 783-801. doi: 10.1007/s00027-017-0546-z
[26] Winemiller K O, Fitzgerald D B, Bower L M, et al. Functional traits, convergent evolution, and periodic tables of niches [J]. Ecology Letters, 2015, 18(8): 737-751. doi: 10.1111/ele.12462
[27] Froese R. FishBase. world wide web electronic publication, 2009.
[28] Villéger S, Mason N W H, Mouillot D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology [J]. Ecology, 2008, 89(8): 2290-2301. doi: 10.1890/07-1206.1
[29] Mason N W H, Mouillot D, Lee W G, et al. Functional richness, functional evenness and functional divergence: the primary components of functional diversity [J]. Oikos, 2005, 111(1): 112-118. doi: 10.1111/j.0030-1299.2005.13886.x
[30] Laliberté E, Legendre P. A distance-based framework for measuring functional diversity from multiple traits [J]. Ecology, 2010, 91(1): 299-305. doi: 10.1890/08-2244.1
[31] Pinkas L, Oliphant M S, Iverson I L K. Food Habits of Albacore, Bluefin Tuna and Bonito in California Waters [C]. California Department of Fish and Game Fish Bulletin, 1971.
[32] 胡传林, 黄祥飞. 保安湖渔业生态和渔业开发技术研究文集 [M]. 北京: 科学出版社, 1991. Hu C L, Huang X F. Collection of Research Papers on Fishery Ecology and Fishery Development Technology of Bao’an Lake [M]. Beijing: Science Press, 1991.
[33] 梁彦龄, 刘伙泉. 草型湖泊资源、环境与渔业生态学管理(一) [M]. 北京: 科学出版社, 1995. Liang Y L, Liu H Q. Resources, Environment and Fishery Ecological Management of Macrophytic Lakes [M]. Beijing: Science Press, 1995.
[34] 张堂林. 扁担塘鱼类生活史策略、营养特征及群落结构研究 [D]. 武汉: 中国科学院水生生物研究所, 2005: 1-5. Zhang T L. Life-history strategies, trophic patterns and community structure in the fishes of Lake Biandantang [D]. Wuhan: Institute of Hydrobiology, Chinese Academy of Sciences, 2005: 1-5.
[35] Xie S, Cui Y, Zhang T, et al. The spatial pattern of the small fish community in the Biandantang Lake–A small shallow lake along the middle reach of the Yangtze River, China [J]. Environmental Biology of Fishes, 2000, 57(2): 179-190. doi: 10.1023/A:1007541031222
[36] 李为. 长江中游浅水湖泊鳜放养渔业的生态学研究 [D]. 武汉: 中国科学院水生生物研究所, 2011, 32-35. Li W. Ecological studies on the stocking fishery of mandarin fish Siniperca chuatsi (Basilewsky) in shallow lakes along the middle reach of the Yangtze River [D]. Wuhan: Institute of Hydrobiology, Chinese Academy of Sciences, 2011, 32-35.
[37] Li W, Zhang T, Li Z. Spatial distribution and abundance of small fishes in Xiaosihai Lake, a shallow lake along the Changjiang (Yangtze) River, China [J]. Chinese Journal of Oceanology and Limnology, 2010, 28(3): 470-477. doi: 10.1007/s00343-010-9082-6
[38] Li W, Hicks B J, Lin M, et al. Impacts of hatchery-reared mandarin fish Siniperca chuatsi stocking on wild fish community and water quality in a shallow Yangtze Lake [J]. Scientific Reports, 2018(8): 11481. doi: 10.1038/s41598-018-29758-z
[39] 李为, 朱挺兵, 郭传波, 等. 肖四海不同渔业发展阶段的水环境特征分析 [J]. 长江流域资源与环境, 2014, 23(8): 1090-1096. doi: 10.11870/cjlyzyyhj201408007 Li W, Zhu T B, Guo C B, et al. Analysis of water environmental characteristics at different fisheries stages in Lake Xiaosihai [J]. Resources and Environment in the Yangtze Basin, 2014, 23(8): 1090-1096. doi: 10.11870/cjlyzyyhj201408007
[40] 张春兰. 长江中游江湖阻隔后鱼类资源的变动 [D]. 北京: 北京大学, 2004. Zhang C L. Changes of fish resources after isolation of rivers and lakes in middle reaches of the Yangtze River [D]. Beijing: Peking University, 2004.
[41] 胡军华, 张春兰, 胡慧建. 长江中游湖泊鱼类物种多样性结构及动态 [J]. 水生态学杂志, 2008, 29(1): 47-51. doi: 10.15928/j.1674-3075.2008.05.008 Hu J H, Zhang C L, Hu H J. Structure and variation of fish diversity of lakes in the central Yangtze River [J]. Journal of Hydroecology, 2008, 29(1): 47-51. doi: 10.15928/j.1674-3075.2008.05.008
[42] 郑鹏, 蒋小明, 曹亮, 等. 江湖阻隔背景下东部平原湖泊鱼类功能特征及多样性变化 [J]. 湖泊科学, 2022, 34(1): 151-168. doi: 10.18307/2022.0114 Zheng P, Jiang X M, Cao L, et al. Long-term changes in the functional trait composition and diversity of fish assemblages in eastern plain lakes under the regime of river-lake connectivity loss [J]. Journal of Lake Sciences, 2022, 34(1): 151-168. doi: 10.18307/2022.0114
[43] Vandermeer J H. Niche theory [J]. Annual Review of Ecology Evolution and Systematics, 1972(3): 107-132.
[44] Zuppinger-Dingley D, Schmid B, Petermann J S, et al. Selection for niche differentiation in plant communities increases biodiversity effects [J]. Nature, 2014, 515(7525): 108-111. doi: 10.1038/nature13869
[45] Liu X, Wang H. Effects of loss of lateral hydrological connectivity on fish functional diversity [J]. Conservation Biology, 2018, 32(6): 1336-1345. doi: 10.1111/cobi.13142
[46] Jiang X, Zheng P, Cao L, et al. Effects of long-term floodplain disconnection on multiple facets of lake fish biodiversity: decline of alpha diversity leads to a regional differentiation through time [J]. Science of the Total Environment, 2021(763): 144177.
[47] Sullivan S M P, Watzin M C. Stream-floodplain connectivity and fish assemblage diversity in the Champlain Valley, Vermont, USA [J]. Journal of Fish Biology, 2009, 74(7): 1394-1418. doi: 10.1111/j.1095-8649.2009.02205.x
[48] Bolland J D, Nunn A D, Lucas M C, et al. The importance of variable lateral connectivity between artificial floodplain waterbodies and river channels [J]. River Research and Applications, 2012, 28(8): 1189-1199. doi: 10.1002/rra.1498
[49] Hawkins C P, Mykrä H, Oksanen J, et al. Environmental disturbance can increase beta diversity of stream macroinvertebrate assemblages [J]. Global Ecology and Biogeography, 2015, 24(4): 483-494. doi: 10.1111/geb.12254
[50] Penha J, Landeiro V L, Ortega J C, et al. Interchange between flooding and drying, and spatial connectivity control the fish metacommunity structure in lakes of the Pantanal wetland [J]. Hydrobiologia, 2017, 797(1): 115-126. doi: 10.1007/s10750-017-3164-9
[51] 陈俊伊, 王康, 郭钰伦, 等. 基于稳定同位素技术的保安湖食物网结构特征研究 [J]. 水生生物学报, 2022, 46(5): 699-706. doi: 10.7541/2021.2021.057 Chen J Y, Wang K, Guo Y L, et al. Food web structure of the Bao’an Lake by analysis of stable carbon and nitrogen isotopes analysis [J]. Acta Hydrobiologica Sinica, 2022, 46(5): 699-706. doi: 10.7541/2021.2021.057
-
期刊类型引用(4)
1. 金禹成,王晓玲,秦瑜,肖满义,周禹廷,祝贵兵. 拒马河沉积物N_2O产生潜势、途径与微生物群落结构的研究. 环境科学学报. 2024(08): 311-322 .  百度学术
百度学术
2. 李君轶,吴金明,杜浩,张辉,邵俭,危起伟. 嘉陵江源鱼类群落结构特征与环境因子的关系. 淡水渔业. 2024(05): 24-32 . 百度学术
3. 康玲玲,马超,孙光,盛祥锐,韩静,罗遵兰. 潮白河怀柔段鱼类多样性特征及水生态系统健康评价. 环境科学研究. 2024(10): 2214-2224 . 百度学术
4. 杨姣姣,陈冬,黄立成,李杨,董晋延,黄超,王纯波,刘永定,杜劲松,潘珉. 滇池不同湖区浮游动物群落稳定性及其驱动因子分析. 湖泊科学. 2023(05): 1752-1766 . 百度学术
其他类型引用(4)





计量
- 文章访问数: 1858
- HTML全文浏览量: 670
- PDF下载量: 93
- 被引次数: 8