
HYDROACOUSTIC SURVEYS ON SPATIAL-TEMPORAL DISTRIBUTION OF FISHES DURING EARLY FISHING BAN PERIOD IN POYANG LAKE
-
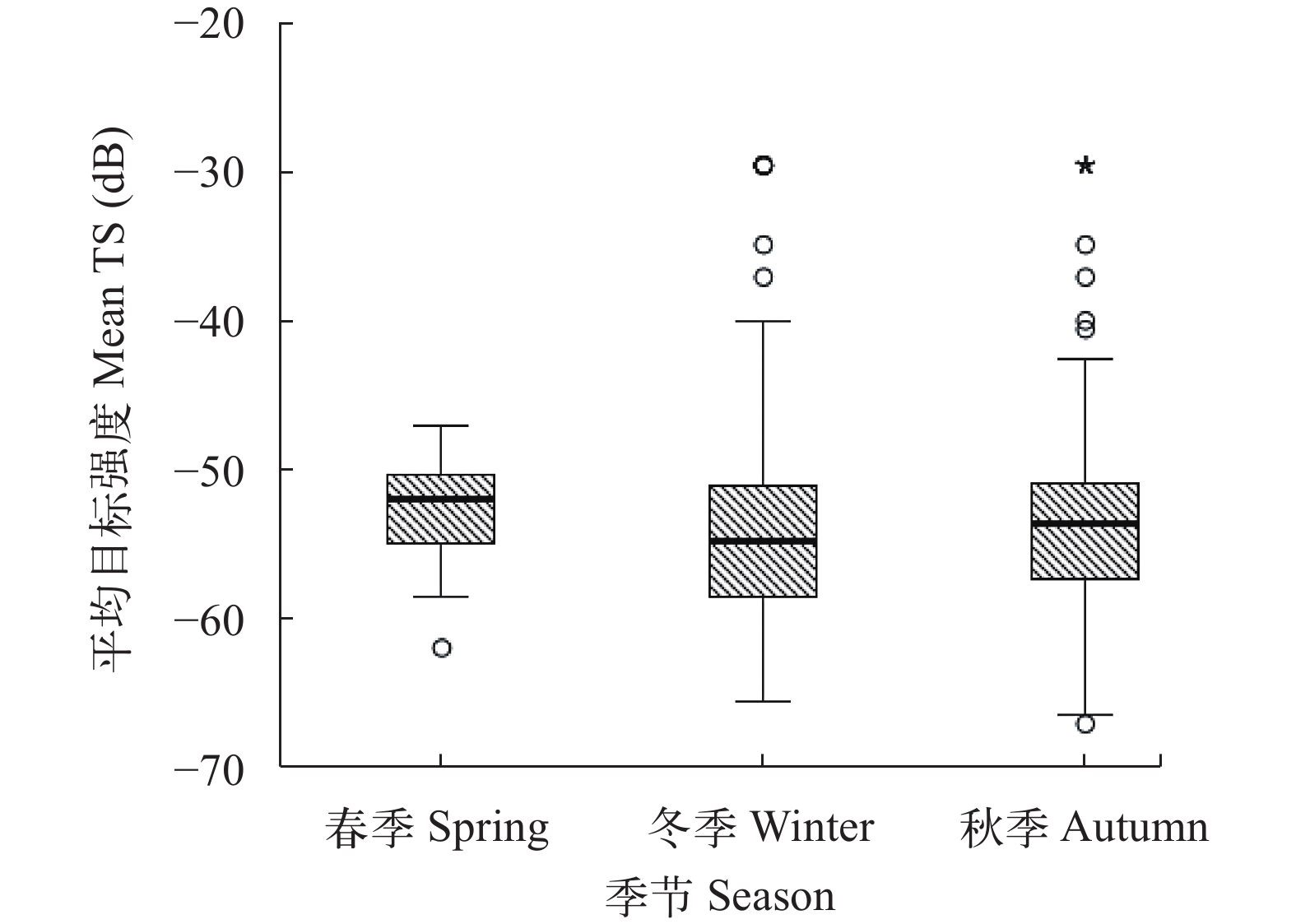
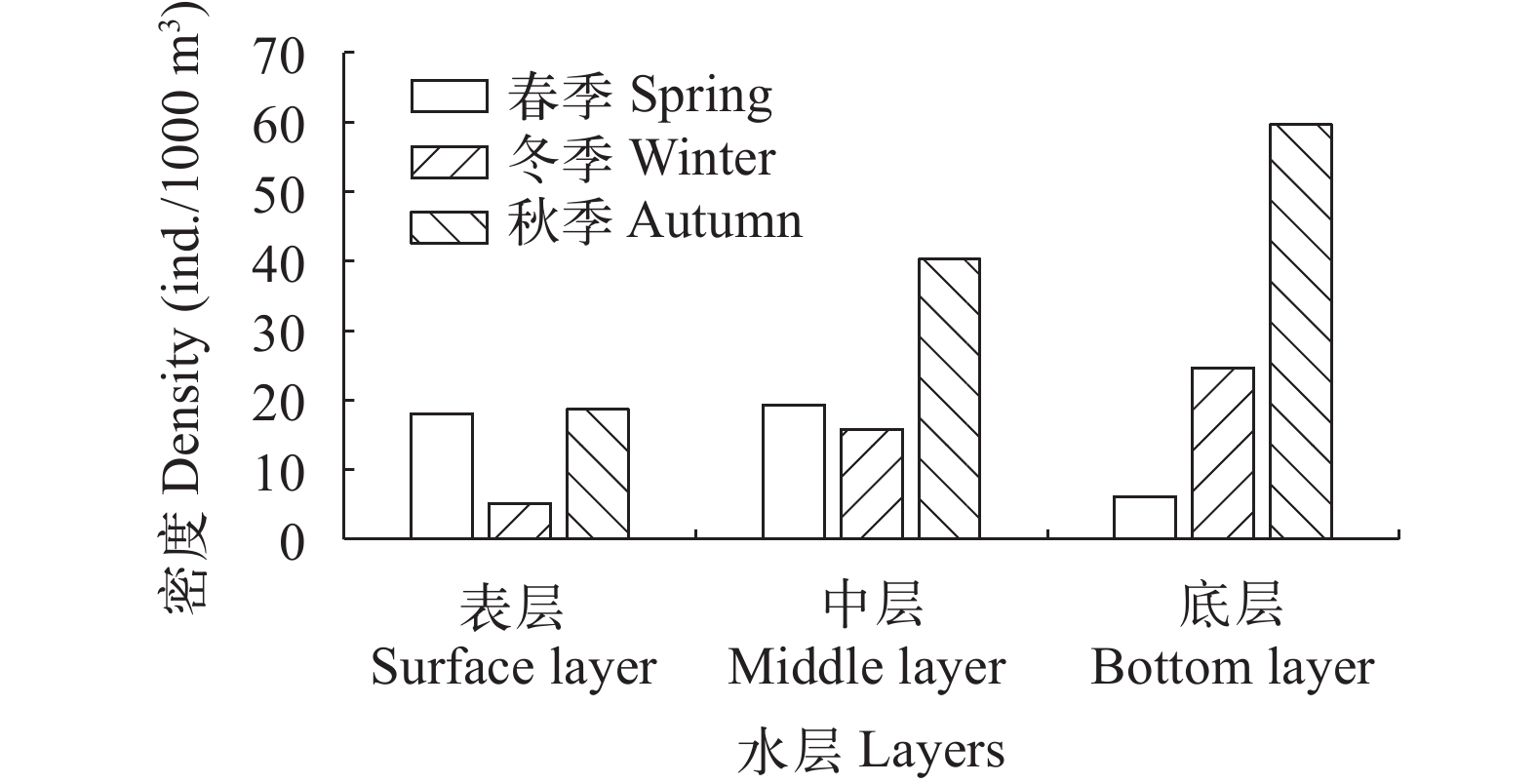
摘要: 研究于2020—2021年使用Simrad EY60鱼探仪对鄱阳湖进行了秋季、春季和冬季共3次的水声学调查, 并同步开展渔获物调查作为补充, 分析鄱阳湖鱼类资源时空分布变化特征。结果表明: 在季节分布上, 鱼类目标强度和密度值都存在显著差异(P<0.05), 在目标强度上表现为冬季[(–51.0±14.13) dB]>春季[(–52.10±4.59) dB]>秋季[(–52.71±9.95) dB], 在鱼类密度上表现为冬季(54.61 ind./1000 m3)>秋季(46.10 ind./1000 m3)>春季(18.54 ind./1000 m3); 在水平分布上, 鱼类资源空间分布不均且不同湖区间有显著差异(P<0.05), 秋季鱼类主要分布在中部湖区松门山, 冬季鱼类主要分布在北部湖区通江水道; 在垂直分布上, 秋季和冬季均表现为底层>中层>表层, 春季表现为表层>中层>底层。综合来看, 鄱阳湖鱼类时空分布与鱼类的生活习性如产卵、育肥和越冬等因素密切相关。研究结果为从宏观空间尺度分析区域鱼类时空变动特征提供参考, 也为鄱阳湖禁捕效果评估及生物完整性评价提供数据支撑。Abstract: Poyang Lake is the largest freshwater lake in China, and a typical natural lake in the middle and lower reaches of the Yangtze River. Different habitat season landscapes can meet the needs of fish in different life history stages, so that fish resources show dynamic distribution characteristics. The 10-year fishing ban have been implemented in the Yangtze River basin, including Poyang Lake since Jan 1, 2021, to conserve the aquatic organism of the Yangtze River. In this study, we applied hydroacoustic analysis with Simrad EY60 split-beam echo sounder, to detect the spatial-temporal distribution of fish in Poyang Lake from 2020 to 2021. The results showed that more than half of the fish were 10—30 cm of body length in all three seasons, deduced from the fish target strength (TS) signal. However, the TS varied significantly among seasons (P<0.05), with the highest TS in winter (–51.0±14.13) dB and the lowest (52.71±9.95) dB in autumn. The horizontal distribution of fish density was not homogeneous, and also significantly different among seasons (P<0.01). The maximum density was 440.22, 169.19, and 554.18 ind./1000 m3 in autumn, spring, and winter, respectively. Vertically, the fish density decreased gradually from the bottom to the surface in autumn and winter, while the fish density in the surface and middle layers was higher than that in the bottom in spring. The fish density varied significantly in the same water layer among seasons (P<0.01). This study obtained the spatial and temporal distribution characteristics of fish in Poyang Lake by using hydroacoustic method at the beginning of fishing ban, and provided a reference for the study of fish resource in the region at a macro spatial scale under natural conditions. The results can also support for the evaluation of the fishing ban effect and the evaluation of biological integrity during the ten-year fishing ban in Poyang Lake.
-
Keywords:
- Fish resources /
- Hydroacoustics /
- Temporal and spatial distribution /
- Fishing ban /
- Poyang Lake
-
蛋白源是水产动物配合饲料的主要成分。鱼粉因高消化率、适宜的氨基酸和脂肪酸组成, 成为水产动物饲料中最常用且最重要的蛋白源。随着水产养殖业的发展, 鱼粉的需求量不断增加, 但是随着气候环境的变化, 海洋渔业资源不断衰退, 鱼粉的供需矛盾变得极为紧张。价格低廉且来源广泛的以豆粕、棉粕、菜粕为主的植物蛋白被大量应用于水产饲料, 但这些植物蛋白中存在抗营养因子、缺乏如赖氨酸和蛋氨酸等必需氨基酸, 无法完全替代鱼粉, 使用具有一定的局限性。近年来, 以菌体和藻类为主的单细胞蛋白在水产饲料中的应用也逐步深入。
乙醇梭菌蛋白(Clostridium autoethanogenum protein, CAP)是一种价廉质优的单细胞蛋白, 其蛋白含量, 氨基酸组成相对平衡。同时, CAP是乙醇工业化生产中得到的副产品, 产量巨大, 产能稳定, 因此CAP的生产不会消耗额外的其他资源, 是一种可靠的新型饲料蛋白源[1]。研究表明, CAP可在建鲤(Cyprinus carpiovar Jian)的配合饲料中替代植物蛋白[1], 在大口黑鲈(Micropterus salmoides)饲料中替代鱼粉[2, 3], 吉富罗非鱼(Oreochromis niloticus)配合饲料中替代豆粕[4]。此外, 在甲壳类动物中, CAP也可以替代鱼粉用于凡纳滨对虾(Litopenaeus vannamei)[5]和克氏原螯虾(Procambarus clarkii)[6]的配合饲料。
日本沼虾(Macrobrachium nipponense)俗称河虾、青虾, 是我国国内广泛存在的本土淡水虾品种。因其生长快、品质高、抗病性强、经济价值高而开始被大面积养殖。2023年, 日本沼虾全国年产量已超22.6万吨, 主要产区位于江苏省。2023年江苏省日本沼虾养殖产量已经超过10万吨, 接近全国产量的一半[7]。研究表明日本沼虾的配合饲料中需要约25%的鱼粉[8]。在鱼粉使用成本过高、用量有限的当下, 若能使用CAP替代鱼粉, 则能够节省更多的成本, 让日本沼虾的养殖具有更高的经济效益。本试验以CAP替代日本沼虾配合饲料中不同的鱼粉水平, 评估CAP替代鱼粉对日本沼虾生长、蛋白质和脂肪代谢的影响, 以探讨在日本沼虾配合饲料中CAP替代鱼粉的可行性以及最适宜添加量, 为新型单细胞蛋白源CAP在水产饲料中的应用提供数据支撑。
1. 材料与方法
1.1 试验动物
日本沼虾幼虾[平均初始体重(0.102±0.001) g, 初始体长约15 mm]来自淡水渔业研究中心宜兴大浦基地养殖场(江苏无锡)。在饲养试验前, 用青虾“太湖2号”专用商品料(江苏冠乾特种水产饲料有限公司, 常州溧阳)饲喂幼虾2周, 以适应养殖环境。禁食24h后, 将日本沼虾(n=1200)随机分配到24个养殖桶, 每个桶50尾虾。
1.2 饲料制备
乙醇梭菌蛋白(CAP)由北京首朗生物科技有限公司(中国河北)提供, 干物质含量92.83%、粗蛋白含量为89.25%、粗脂肪含量为2.30%, 其中必需氨基酸赖氨酸含量为8.59%、蛋氨酸含量为3.54%。利用CAP替代不同水平鱼粉, 配制6组等氮等脂(粗蛋白37.25%, 粗脂肪7.70%)的试验饲料(表 1)。对照组D1饲料中含25%鱼粉, D2组饲料中CAP替代10%鱼粉、D3组饲料CAP替代20%鱼粉、D4组饲料CAP替代30%鱼粉、D5组饲料CAP替代40%鱼粉和D6组饲料CAP替代50%鱼粉。在各组饲料中添加L-赖氨酸和D、L-蛋氨酸以保证各组饲料中赖氨酸和蛋氨酸平衡。根据试验饲料配方表, 将所有原料粉碎过60目筛, 随后采用逐级混合扩大法将所有原料混合均匀。采用双螺杆挤条机(F-26, 广州华中光电科技有限公司, 广州, 中国)挤压混合原料, 制备直径为1.0 mm的日本沼虾饲料。饲料在室温晾干后, –20℃保存备用。
表 1 试验饲料配方和营养成分Table 1. Experimental diets formulation and proximate composition原料Ingredient 鱼粉替代水平 Fishmeal replacement (%) D1 (0) D2 (10%) D3 (20%) D4 (30%) D5 (40%) D6 (50%) 鱼粉Fish meal 25.00 22.50 20.00 17.50 15.00 12.50 乙醇梭菌蛋白CAP 0.00 1.86 3.70 5.57 7.42 9.30 豆粕Soybean meal 20.00 20.00 20.00 20.00 20.00 20.00 菜粕Canola meal 10.00 10.00 10.00 10.00 10.00 10.00 虾粉Shrimp powder 5.00 5.00 5.00 5.00 5.00 5.00 花生粕Peanut meal 8.00 8.00 8.00 8.00 8.00 8.00 乌贼膏Squid 2.00 2.00 2.00 2.00 2.00 2.00 α-淀粉α-starch 17.16 17.66 18.15 18.63 19.12 19.59 米糠Rice bran 5.00 5.00 5.00 5.00 5.00 5.00 鱼油Fish oil 1.50 1.50 1.50 1.50 1.50 1.50 豆油Soybean oil 1.20 1.39 1.59 1.79 1.99 2.19 大豆磷脂Soy lecithin 0.50 0.50 0.50 0.50 0.50 0.50 胆固醇Cholesterol 0.50 0.50 0.50 0.50 0.50 0.50 磷酸二氢钙Ca (H2PO4)2 2.00 2.00 2.00 2.00 2.00 2.00 多维多矿预混Vitamin & Mineral premix1 1.00 1.00 1.00 1.00 1.00 1.00 维生C Vitamin C 0.50 0.50 0.50 0.50 0.50 0.50 氯化胆碱Chlorine chloride 0.25 0.25 0.25 0.25 0.25 0.25 赖氨酸Lysine 0.26 0.23 0.21 0.18 0.15 0.12 蛋氨酸Met 0.13 0.11 0.10 0.08 0.07 0.05 营养成分(%干物质) Proximate Composition (% of dry matter) 粗蛋白Crude protein 37.38 37.22 37.27 37.28 37.21 37.25 粗脂肪Crude lipid 7.65 7.71 7.82 7.72 7.75 7.65 粗灰分Ash 10.54 10.76 10.76 10.59 10.42 10.01 赖氨酸Lysine 2.71 2.71 2.71 2.71 2.71 2.71 蛋氨酸Methionine 0.95 0.95 0.95 0.95 0.95 0.95 注: 1多维多矿预混料为无锡华诺威动物保健品有限公司提供的淡水虾多维多矿预混料Note: 1 Vitamin and mineral premix are provided by Wuxi Hanove Animal Health Products Co., Ltd. 1.3 饲养管理
PVC养殖桶(Φ1500 mm, 高1000 mm, 加水后含水体800 L)随机分为6组, 每组饲料投喂4个养殖桶。采用静水养殖, 桶内放置塑料假叶子作为遮蔽物和躲避场所, 持续增氧, 隔天换水。养殖期间每周测定水质指标2次, 溶解氧≥ 6 mg/L, 氨氮含量<0.2 mg/L、pH 7.0—8.5、水温 25—28℃。投喂量为虾体重量的3%, 每天投喂3次(7:00、12:00和18:00), 投喂结束后1h吸出残饵和粪便, 隔天对养殖桶进行换水。饲养试验持续8周。
1.4 样品采集
在养殖试验结束后, 禁食24h。对每个养殖桶称重, 对存活的试验虾进行计数, 并根据以下公式计算存活率(SR)、末均重(FBW)、增重率(WGR)、特定生长率(SGR)、饵料系数(FCR)、蛋白质效率(PER)等生长相关指标。
存活率(SR, %)=(n2/n1)×100
增重率(WGR, %)=(w2–w1)/w1×100
特定生长率(SGR, %/d)=(Ln w2–Ln w1)/d×100
饲料系数(FCR)= f /(w2–w1)
蛋白质效率(PER)=(w2–w1)/(f×p)×100
式中, n1为试验初虾尾数, n2为试验末虾尾数, w1为试验初虾均重, w2为试验末虾均重, d为养殖天数, f为摄食饲料量, p为饲料蛋白质含量。
每桶随机取6尾虾采血, 虾置于冰上, 0.5 mL注射器自围心腔以抗凝剂和血淋巴1﹕1的比例采血(抗凝剂: 柠檬酸三钠 13.2 g/L、柠檬酸 4.8 g/L、葡萄糖14.7 g/L), 4℃ 3000 r/min离心10min制备血清, 3尾虾混合成血淋巴混合样, –20℃保存, 以备血淋巴生化指标测定。随后采集该6尾虾的肝胰腺, 其中2尾虾的肝胰腺混合成1个样, 每桶共采集3个肝胰腺混合样, –20℃保存, 用于肝胰腺消化酶测定。另取2尾虾的肝胰腺置于TRIzol (TaKaRa, 大连, 中国)中, –80°℃保存, 用于蛋白质和脂代谢相关基因测定。此外, 每桶随机取6尾虾, 混合成一个全虾样品, –20℃保存, 以备全虾体成分测定。随机取12尾虾的肌肉混合, –20℃保存, 以备肌肉氨基酸含量测定。
1.5 营养成分测定及肌肉氨基酸测定
根据AOAC[17]方法测定饲料和全虾样品的水份、粗蛋白质、粗脂肪和灰分含量。饲料或全虾在105℃烘箱中干燥至恒重后测得水份含量。取0.5 g左右饲料或者全虾的干物质样品, 利用全自动凯氏定氮仪(海能, 中国)通过凯氏定氮法测定粗蛋白。索氏提取法利用石油醚浸提出脂肪, 测定粗脂肪; 马弗炉550℃灼烧6.5h, 测定粗灰分。
肌肉氨基酸含量根据《饲料中氨基酸的测定》(GB/T 18246-2019)进行测定。肌肉冷冻干燥后, 用6 mol/L盐酸水解24h后, 氨基酸自动分析仪(sykam S-433D, 赛卡姆, 德国)进行测定及分析。
1.6 血淋巴生化指标及肝胰腺消化酶活性测定
血淋巴总蛋白(Total protein, TP)、总胆固醇(Total cholesterol, TC)、甘油三酯(Triglyceride, TG)、尿素(Urea)、葡萄糖(Glucose, GLU)均采用全自动生化分析仪(BS-400型, 迈瑞, 深圳, 中国)利用相应测定试剂盒(深圳迈瑞医药国际有限公司)分析测定。
取0.1 g肝胰腺样品, 加入0.9 mL无菌生理盐水, 冰浴匀浆, 4℃ 4000 r/min离心10min, 取上清液。胰蛋白酶(TRY)、脂肪酶(LPS)和淀粉酶(AMY)均按试剂盒(南京建成生物工程研究所)提供的方法进行测定。
1.7 蛋白质代谢和脂代谢相关基因表达
取TRIzol保存的肝胰腺样品, 用RNAiso plus (TaKaRa, 大连, 中国)提取总RNA, 琼脂糖凝胶电泳和Nanodrop 2000测定RNA浓度和纯度, 调节RNA浓度至500 ng/μL。反转录试剂盒PrimeScriptTM RT Master Kit (TaKaRa, 大连, 中国)将RNA反转录成cDNA, 反应体系为20 μL(5 × gDNA 2 μL, gDNA Eraser 1 μL, RNase free H2O 5 μL, Total RNA 2 μL, 42℃孵育2min; 加入5 × Primer buffer 4 μL, PrimeScriptTM RT Enzyme Mix 1 μL, Rt Primer Mix 1 μL, RNase free H2O 4 μL)。反转录程序为: 37℃ 15min, 85℃ 5s。合成的cDNA模板, 采用TB GreenTM Premix Ex TaqTM Kit(TaKaRa, 大连, 中国)在定量PCR仪(Bio-Rad, USA)进行荧光定量PCR反应。反应体系20 μL(SYBR Premix Ex Taq 10 μL, Primer 各0.8 μL, dd H2O 6.4 μL, cDNA 2 μL), 反应程序为: 95℃ 预变性30s; 95℃ 5s, 60℃ 30s, 72℃ 30s (循环30次); 溶解曲线温度范围为50—95℃。本试验所用基因的引物序列见表 2[8—12], 所有引物委托上海捷瑞生物工程有限公司合成。以β-actin为内参基因, 根据2−ΔΔCt法[13]计算基因相对表达量。
表 2 基因引物序列Table 2. RT-PCR primer sequence目的基因
Target gene引物序列
Primer sequence (5′—3′)序列登录号
GenBank
accession No.参考文献
Referencetor AACAAGTCTCGTCCGTGTCC — Zhou 等[8] TTGAGCAGCTTCACGGCTTA atf4 GGCGGTGCAGTTAAACACTC — Zhou等[8] GTCAGTTTCACCCATGTCGC s6k GGTCTTTGAGGGCTTTACG ON693253 郑金仙等[9] GTCCAGCAGAGTTTGGTGTAT acc CAAGGTCCACTACATGGTCT KP690138 丁志丽等[10] ACTCTTCCCAAACTCTCTCC fas CGGTCAGACAAACTACGGCT MK307767.1 Wang等[11] CACTGAATAGCCACCCCAGG elovl6 TGGTGCACAGTATCATGTAC KU953779 Luo等[12] TCATAGGAAACGTGACACTC hsl GAAGGCCAGCGCTAATTTCG MK633965.1 Wang等[11] TCGAACCACCCATGAGAAGC sr-bⅠ TTATCCCTGGTGTGAATGTG KP658863 丁志丽等[11] GAACTCTTCCCATTCCAACT β-actin GTGCCCATCTACGAGGGTTA FL589653.1 Luo等[12] CGTCAGGGAGCTCGTAAGAC 注: tor, target of rapamycin雷帕霉素靶蛋白; atf4, activating transcription factor 4转录激活因子4; s6k, ribosomal protein S6 kinase核糖体S6激酶; acc, acetyl-CoA carboxylase乙酰辅酶A羧化酶; fas, fatty acid synthase脂肪合成酶; hsl, hormone-sensitive lipase激素敏感脂肪酶; sr-bⅠ, scavenger receptor class B type Ⅰ, B类Ⅰ型清道夫受体; elovl6, elongase of very-long-chain fatty acids长链脂肪酸延长酶; β-actin, β-肌动蛋白 1.8 数据统计与分析
结果数据用平均值±标准误(mean±SE)表示。利用SPSS 20.0软件对数据进行单因素方差分析(One-way ANOVA), 使用Turkey进行多重比较, 当P<0.05表示差异显著。使用GraphPad Prism8.0进行作图。
2. 结果
2.1 乙醇梭菌蛋白替代鱼粉对日本沼虾生长、饲料利用率的影响
在CAP替代鱼粉饲喂日本沼虾8周后, 其生长和饲料利用率如表 3所示。CAP替代不同水平鱼粉对日本沼虾存活率没有显著影响。与对照组相比, D2和D3组日本沼虾的FBW、WGR和SGR差异不显著(P>0.05), 而D4、D5和D6组日本沼虾的FBW、WGR和SGR则显著降低(P<0.05)。此外, D5 和D6组FCR显著高于对照组(P<0.05), 而其他各组与对照组差异不显著(P>0.05), 而其他3组FCR则显著高于D1组。PER恰好相反, D5和D6组PER显著低于对照组(P<0.05)。
表 3 乙醇梭菌蛋白替代鱼粉后对生长和饲料利用的影响Table 3. Effects of CAP replacing fishmeal on growth performance and feed utilization指标Index 鱼粉替代水平 Fishmeal replacement (%) D1 (0) D2 (10%) D3 (20%) D4 (30%) D5 (40%) D6 (50%) 存活率SR (%) 74.67±1.33 88.50±2.99 86.00±8.52 85.33±3.53 80.50±4.57 90.50±2.50 末均重FBW (g) 0.76±0.01c 0.56±0.01b 0.57±0.02bc 0.46±0.02a 0.37±0.02a 0.39±0.06a 增重率WGR (%) 461.01±14.90c 392.07±18.74bc 380.87±36.65bc 288.17±10.19ab 199.56±10.89a 253.54±58.93a 特定生长率SGR (% /d) 2.45±0.05c 2.27±0.06bc 2.22±0.12bc 1.92±0.06ab 1.54±0.06a 1.73±022a 饵料系数FCR 1.66±0.04a 1.91±0.08ab 2.01±0.23ab 2.29±0.15ab 3.63±0.18c 3.02±0.53bc 蛋白质效率PER 145.03±3.69c 126.27±5.81bc 122.92±11.75bc 106.57±7.54abc 66.71±3.25a 89.66±19.81ab 注: n=4, 同一行右上角含有相同英文上标字母或者无上标表示差异不显著(P ≥ 0.05); 下同Note: n=4. Means in each line sharing the same letter or absence of superscripts are not significantly different determined by Tukey’s test (P ≥ 0.05); The same applies below 2.2 乙醇梭菌蛋白替代鱼粉对日本沼虾体成分的影响
饲料中CAP替代不同水平鱼粉对日本沼虾的全虾体成分包括水分、粗蛋白、粗脂肪和灰分等指标均没有产生显著的影响(P>0.05)。
表 4 乙醇梭菌蛋白替代鱼粉后对体成分的影响Table 4. Effects of CAP replacing fishmeal on shrimp composition指标Index 鱼粉替代水平 Fishmeal replacement (%) D1 (0) D2 (10%) D3 (20%) D4 (30%) D5 (40%) D6 (50%) 水份Moisture (%) 76.97±0.07 77.11±0.39 76.89±0.38 76.91±0.73 76.967±0.31 76.76±0.49 粗蛋白Protein (%) 14.99±0.15 15.82±0.30 15.26±0.27 15.70±0.38 15.86±0.27 15.91±0.28 粗脂肪Lipid (%) 3.04±0.24 2.85±0.32 3.38±0.21 3.63±0.29 2.98±0.32 3.32±0.22 灰分Ash (%) 5.13±0.43 5.82±0.16 5.33±0.40 6.22±0.20 5.85±0.52 6.17±0.26 2.3 乙醇梭菌蛋白替代鱼粉对日本沼虾肌肉中氨基酸的影响
CAP替代不同水平鱼粉对日本沼虾肌肉中必需氨基酸影响不显著(P>0.05), 但显著影响日本沼虾肌肉非必需氨基酸和总氨基酸含量(P<0.05)。除甘氨酸、精氨酸和苯丙氨酸以外, D2组和D3组日本沼虾肌肉氨基酸含量与对照组差异不显著(P>0.05)。 D6组日本沼虾肌肉的精氨酸含量显著低于对照组(P<0.05), 而其组氨酸、异亮氨酸和赖氨酸等必需氨基酸、以及天冬氨酸、甘氨酸、谷氨酸等非必需氨基酸均显著高于对照组(P<0.05)。
表 5 乙醇梭菌蛋白替代鱼粉对日本沼虾肌肉氨基酸含量的影响Table 5. Effect of CAP replacing FM on the amino acid profile of M. nipponense’s muscle氨基酸含量
Amino acid (%)鱼粉替代水平 Fishmeal replacement (%) D1 (0) D2 (10%) D3 (20%) D4 (30%) D5 (40%) D6 (50%) 亮氨酸Leu 1.52±0.01ab 1.48±0.00a 1.54±0.01ab 1.58±0.01ab 1.58±0.05ab 1.62±0.01b 组氨酸His 0.51±0.01a 0.50±0.00a 0.53±0.00a 0.53±0.01a 0.50±0.01a 0.61±0.03b 异亮氨酸Ile 0.90±0.01a 0.88±0.01a 0.94±0.01ab 0.95±0.00ab 0.92±0.04ab 1.00±0.01b 精氨酸Arg 1.67±0.03c 1.15±0.02b 1.58±0.04c 1.51±0.06bc 1.40±0.12bc 1.27±0.01ab 赖氨酸Lys 1.57±0.02a 1.56±0.00a 1.59±0.01a 1.67±0.00b 1.61±0.02ab 1.73±0.01c 蛋氨酸Met 0.59±0.01 0.59±0.01 0.61±0.01 0.59±0.00 0.61±0.00 0.62±0.01 苯丙氨酸Phe 0.83±0.01b 0.76±0.00a 0.80±0.00ab 0.85±0.01b 0.76±0.00a 0.86±0.01b 苏氨酸Thr 0.72±0.01 0.70±0.00 0.71±0.01 0.74±0.00 0.71±0.00 0.75±0.00 缬氨酸Val 0.88±0.01 0.93±0.03 0.94±0.00 0.90±0.02 0.91±0.01 0.95±0.02 必需氨基酸EAA 9.16±0.11 8.52±0.01 9.23±0.08 9.30±0.01 8.97±0.25 9.39±0.04 天冬氨酸Asp 2.04±0.03a 1.96±0.02a 2.06±0.02a 2.02±0.02a 1.96±0.04a 2.15±0.00b 丝氨酸Ser 0.73±0.01 0.70±0.00 0.71±0.01 0.73±0.01 0.68±0.02 0.71±0.01 甘氨酸Gly 1.30±0.00a 1.43±0.01b 1.54±0.03c 1.50±0.00bc 1.63±0.02d 1.60±0.00cd 丙氨酸Ala 1.32±0.01 1.34±0.02 1.31±0.00 1.39±0.03 1.36±0.02 1.32±0.02 半胱氨酸Cys 0.27±0.00 0.30±0.29 0.32±0.01 0.27±0.01 0.30±0.02 0.29±0.01 酪氨酸Tyr 0.70±0.01ab 0.63±0.00a 0.65±0.01ab 0.71±0.02ab 0.68±0.00ab 0.73±0.00b 谷氨酸Glu 2.99±0.05ab 2.86±0.01a 2.97±0.01ab 3.14±0.01b 3.00±0.01ab 3.27±0.04c 脯氨酸Pro 0.91±0.04b 0.90±0.01b 0.88±0.00ab 0.91±0.02b 0.72±0.08a 1.01±0.03b 非必需氨基NEAA 10.23±0.14ab 10.09±0.02a 10.43±0.08bc 10.66±0.04c 10.45±0.07bc 11.06±0.01d 总氨基酸TAA 19.39±0.25ab 18.61±0.01a 19.65±0.16bc 19.95±0.05bc 19.43±0.31ab 20.45±0.03c 2.4 乙醇梭菌蛋白替代鱼粉对日本沼虾血淋巴生化的影响
由表 6可知, D2组和D3组日本沼虾血淋巴总蛋白含量与对照组差异不显著(P>0.05), 但是D4、D5和D6组日本沼虾血淋巴总蛋白含量显著低于对照组和D2组(P<0.05)。D6组日本沼虾血淋巴尿素氮含量最低, 且显著低于对照组和D2组(P<0.05)。CAP替代高于10%鱼粉时, 日本沼虾血淋巴总胆固醇、甘油三酯显著低于对照组(P<0.05), 葡萄糖显著高于对照组(P<0.05)。
表 6 乙醇梭菌蛋白替代鱼粉后对血淋巴生化的影响Table 6. Effects of CAP replacing FM on hemolymph biochemical indexes指标Index 鱼粉替代水平 Fishmeal replacement (%) D1 (0) D2 (10%) D3 (20%) D4 (30%) D5 (40%) D6 (50%) 总蛋白TP (g/L) 43.14±1.56bc 45.92±2.61c 33.69±3.89ab 33.00±2.00a 31.99±2.09a 26.67±1.28a 总胆固醇TC (mmol/L) 0.78±0.12b 0.87±0.07b 0.32±0.03a 0.37±0.03a 0.26±0.02a 0.29±0.02a 甘油三酯TG (mmol/L) 18.90±1.25b 16.98±1.21b 8.08±0.57a 9.57±0.42a 8.63±0.48a 9.41±0.47a 葡萄糖GLU (mmol/L) 4.12±0.97a 5.16±0.88ab 9.32±1.75bc 8.86±0.81bc 9.82±0.78c 11.18±0.97c 尿素氮Urea (mmol/L) 1.66±0.06b 1.65±0.13b 1.41±0.15ab 1.33±0.05ab 1.24±0.12ab 1.00±0.05a 2.5 乙醇梭菌蛋白替代鱼粉对日本沼虾肝胰腺消化酶活性的影响
CAP替代鱼粉显著影响日本沼虾肝胰腺消化酶活性。在CAP替代10%鱼粉时, 日本沼虾肝胰腺脂肪酶活性与对照组相比, 显著升高(P<0.05)。在CAP替代高于10%鱼粉时, 日本沼虾肝胰腺的消化酶活性与对照组差异不显著(P>0.05)。此外, D4和D5组胰蛋白酶、脂肪酶活性及D6组淀粉酶活性显著低于D2组(P<0.05)。
![]() 图 1 乙醇梭菌蛋白替代鱼粉后对肝胰腺消化酶活性的影响Figure 1. Effects of CAP replacing FM on digestive enzyme activities
图 1 乙醇梭菌蛋白替代鱼粉后对肝胰腺消化酶活性的影响Figure 1. Effects of CAP replacing FM on digestive enzyme activities2.6 乙醇梭菌蛋白替代鱼粉对日本沼虾肝胰腺基因表达的影响
CAP替代不同水平鱼粉显著影响日本沼虾肝胰腺内蛋白质和脂肪代谢相关基因的表达。在CAP替代10%鱼粉时, 蛋白和脂肪代谢基因相对表达量降低, 但与对照组差异不显著(P>0.05)。在CAP替代鱼粉达到20%时, tor、s6k1和atf4基因相对表达量上升。D3组tor基因相对表达量显著高于对照组和D2组(P<0.05)。除D2组外, 其余各组s6k1基因相对表达量与对照组差异不显著(P>0.05), 但显著高于D2组(P<0.05)。D4和D5组atf4基因相对表达量显著高于D2组(P<0.05)。在脂质合成相关基因中, D3、D4和D5组acc基因相对表达量同样与对照组差异不显著(P>0.05), 但显著高于D2组(P<0.05)。D3、D5和D6组fas基因相对表达量显著高于D2组。D6组fas基因表达较对照组显著上调(P<0.05)。D3组和D5组elovl6基因相对表达量显著高于D2和D4组(P<0.05)。与对照组相比, 各试验组日本沼虾肝胰腺hsl和sr-bⅠ基因相对表达量差异不显著(P>0.05), 除D3组sr-bⅠ基因相对表达量显著高于对照组之外(P<0.05)。D2组hsl表达量显著低于D3、D5、D6组(P<0.05); D5组sr-bⅠ基因相对表达量显著低于D3和D6组(P<0.05)。
![]() 图 2 乙醇梭菌蛋白替代鱼粉后对肝胰腺基因表达的影响Figure 2. Effects of CAP replacing FM on hepatopancreas gene expression
图 2 乙醇梭菌蛋白替代鱼粉后对肝胰腺基因表达的影响Figure 2. Effects of CAP replacing FM on hepatopancreas gene expression3. 讨论
3.1 乙醇梭菌蛋白替代鱼粉对日本沼虾生长的影响
乙醇梭菌(Clostridium autoethanogenum)是一种天然的非致病性菌株, 是一种革兰氏阳性、成短杆状、能形成孢子的严格厌氧菌。CAP是由乙醇梭菌以炼钢废气中的一氧化碳为碳源、发酵生产工业乙醇, 经细菌分离、喷雾干燥后得到的副产物[14]。本研究结果显示在日本沼虾饲料中使用CAP作为蛋白源, 对日本沼虾的存活率无显著影响, 且CAP替代50%的鱼粉时, 日本沼虾存活率最高。据报道CAP作为单细胞蛋白, 在黑鲷(Acanthopagrus schlegelii)[14]和大口黑鲈[2]饲料中添加18%而不影响其生长; 在建鲤饲料中添加量可达20%[1], 在大口黑鲈饲料中可以替代15%的鱼粉[3]。此外, CAP在吉富罗非鱼中可以替代豆粕, 使用量达到20%时能显著提高其生长[4]。在甲壳类动物中, CAP最高可替代凡纳滨对虾饲料中30%鱼粉而不会对虾的生长表现产生不利影响[5]。克氏原螯虾的研究结果表明, CAP替代25%的鱼粉生长效果最佳, 替代全部鱼粉时也不影响克氏原螯虾的生长[6]。本研究发现日本沼虾饲料中CAP替代20%鱼粉对其生长无影响。此外, 日本沼虾中研究发现泰森鸡肉粉、鸡蛋粉、肠黏膜蛋白粉及水解羽毛粉等动物性蛋白也可以替代饲料中20%的鱼粉而对日本沼虾的生长不产生显著影响[15]。这表明CAP作为新的单细胞蛋白, 可以在水产动物饲料中进行应用, 但是在不同水产动物中差异较大。
CAP粗蛋白含量高, 可达85%以上, 且总氨基酸占蛋白质含量的85%, 富含赖氨酸, 其氨基酸构成与鱼粉相似, 含有多种微量元素, 且不含抗营养因子[13]。在本试验中, CAP替代30%以上的鱼粉时, 其FCR显著升高而PER显著降低, 这说明虽然CAP粗蛋白含量高于鱼粉, 但其蛋白质利用率显著低于鱼粉。这一结果与凡纳滨对虾类似, 当CAP替代超过30%鱼粉时, FCR显著降低[5]; 但是与克氏原螯虾的结果略有不同, CAP替代鱼粉对FCR无显著影响[6]。在黑鲷[13]、建鲤[1]和吉富罗非鱼[4]等鱼类中, 随着CAP添加量的增加, PER显著增加; 而在大口黑鲈中, CAP替代25%鱼粉时, FCR显著升高[3]。这说明不同物种对于CAP的消化吸收利用率存在一定的差异, 总体来看, 鱼类对CAP的利用率优于甲壳类。
3.2 乙醇梭菌蛋白替代鱼粉对日本沼虾肌肉品质的影响
本研究结果表明CAP对日本沼虾水分、粗蛋白、粗脂肪和灰分无影响, 表明饲料中CAP替代鱼粉不会影响日本沼虾养成分。这与建鲤和吉富罗非鱼的研究结果相似, 饲料中添加CAP不影响建鲤和吉富罗非鱼的体组成[1, 4]。此外, 大西洋鲑(Salmo salar)和庸鲽(Hippoglossus hippoglossus)也有相似结果, 饲料中添加细菌蛋白不影响大西洋鲑和庸鲽体成分[16—19]。与之相反, 在南美白对虾中, 对虾肌肉的粗蛋白会随着CAP添加量的上升而下降, 而肌肉水份则呈相反的趋势[5]; CAP会导致大口黑鲈肌肉粗脂肪含量下降[2]。产生这样差异的原因还需进一步研究。
肌肉是鱼最大的可食部分, 因此肌肉氨基酸含量不仅反应所摄食的饲料营养氨基酸平衡, 也代表水产品的营养品质的重要指标。在本研究中, CAP的应用对日本沼虾肌肉必需氨基酸影响不显著, 但是对非必需氨基酸和总氨基酸含量产生一定的影响。这可能是由于CAP与鱼粉的氨基酸组成差异导致的。日粮中CAP过量会显著提高日本沼虾肌肉中非必需氨基酸和总氨基酸含量。但在南美白对虾中表现出了与日本沼虾相反的趋势, 当45%以上的鱼粉被CAP替代时, 肌肉中必需氨基酸的含量下降[5, 20]。在本研究中发现, CAP过量使用不影响虾体肌肉必需氨基酸含量, 这可能与配方时, 调节了赖氨酸和蛋氨酸等限制性必需氨基酸的水平, 在一定程度上对饲料氨基酸的平衡进行了调节。由此可见, CAP的氨基酸组成较鱼粉还有所差距, 当利用CAP替代鱼粉时可能需要对必需氨基酸进行补充调节。
3.3 乙醇梭菌蛋白替代鱼粉对日本沼虾代谢的影响
日本沼虾饲料中CAP替代10%鱼粉对血淋巴总蛋白、尿素氮、总甘油三酯、总胆固醇和葡萄糖等含量影响不显著, 这可能与CAP替代10%鱼粉时日本沼虾肝胰腺的蛋白酶、脂肪酶和淀粉酶等消化酶活性在一定程度上升高有关。而当日本沼虾饲料中CAP替代20%以上鱼粉时, 血淋巴总蛋白和尿素氮水平显著降低, 这一现象显示出CAP对日本沼虾的蛋白质代谢产生了一些负面影响。此外, 在CAP替代20%鱼粉时, 显著降低血淋巴中胆固醇、甘油三酯的含量, 且显著提高葡萄糖含量, 这说明CAP可能会降低葡萄糖的糖异生作用, 降低日本沼虾体内的脂质合成。血淋巴总甘油三酯和总胆固醇减少的原因也可能是由于CAP存在的某些生物活性化合物具有促进脂质代谢的功能[21]。CAP提高血淋巴葡萄糖水平, 也可能表明CAP的添加显著增强了日本沼虾的能量调节, 这与建鲤的研究结果一致[1]。因此, 在使用CAP作为蛋白源时, 可能在饲料中需要提供更多的脂质以调节动物体内的糖脂代谢过程。
甲壳类动物肝胰腺消化酶活性可以体现机体营养代谢水平[8]。本研究结果表明在CAP替代10%鱼粉时, 日本沼虾肝胰腺的蛋白酶、脂肪酶和淀粉酶均达到峰值; 而在CAP替代20%鱼粉时, 日本沼虾肝胰腺的蛋白酶、脂肪酶和淀粉酶均较峰值下降。这说明在CAP替代水平较低时, 动物机体可以通过消化酶调节来加强机体对脂质的消化和吸收, 进而加强对脂质代谢的调控。而本研究中血淋巴总胆固醇和总甘油三酯的结果能说明这一问题, 当在CAP替代20%及以上鱼粉时, 血淋巴总胆固醇和总甘油三酯水平显著下降。但是, 各处理组日本沼虾肝胰腺的消化酶与对照组差异基本不显著, 这在一定程度上说明CAP替代鱼粉不会对日本沼虾肝胰腺中消化酶活性产生负面影响。当其他动物蛋白如鸡蛋粉、肠溶蛋白、羽毛粉等替代鱼粉时, 日本沼虾肝胰腺的消化酶也没有显著的负面影响[15]。
3.4 乙醇梭菌蛋白替代鱼粉对日本沼虾肝胰腺基因表达的影响
雷帕霉素靶蛋白(Target of rapamycin, TOR)是调节蛋白质合成、细胞骨架重塑及细胞内蛋白质通过自噬降解的细胞信号通路的一个重要基因。活化的TOR可以磷酸化下游靶蛋白——核糖体蛋白S6激酶-1 (Ribosomal protein S6 Kinase-1, S6K1), S6K1磷酸化的增加导致核糖体蛋白S6的过度磷酸化, 从而促进mRNA的翻译, 促进蛋白质合成[22]。本研究结果显示, 在日本沼虾饲料中CAP替代20%鱼粉会提高肝胰腺中tor和s6k1的表达水平, 但替代水平高于20%时, tor基因相对表达量会降低, 这可能是CAP替代20%鱼粉对日本沼虾生长无显著负面影响的主要原因。而在吉富罗非鱼中, CAP的添加不影响tor基因的表达水平, 但在50和100 g/kg CAP组中提高了s6k1基因的表达水平[4]。饲料中蛋白质缺乏或必需氨基酸不平衡激活氨基酸反应信号转导通路, 导致激活转录因子4 (Activating transcription factor 4, ATF4)高表达[23]。本研究发现, CAP替代超过30%鱼粉时, 日本沼虾肝胰腺中atf4表达显著升高, 这表明CAP可能会导致氨基酸不平衡, 影响日本沼虾的生长。此外, 研究表明氨基酸的不平衡会改变氨基酸以外的代谢, 如脂代谢等。氨基酸不平衡激活氨基酸反应信号通路后, 在小鼠和大鼠上会抑制胆固醇合成、脂肪酸合成, 进而导致血清胆固醇降低[24, 25]。本研究也发现高CAP水平会导致日本沼虾血淋巴胆固醇和甘油三酯水平降低, 这可能与氨基酸反应信号通路被激活相关。
新的脂肪酸合成反应由细胞质中的乙酰辅酶A羧化酶(ACC)和脂肪酸合成酶(FAS)可以在细胞质内催化新脂肪酸合成。ACC以ATP依赖的方式催化乙酰辅酶A羧化成丙二酰辅酶A, 然后FAS以乙酰辅酶A和丙二酰辅酶A为底物催化的七个酶促反应生成棕榈酸酯[24]。脂肪酸合成速率受到动物机体营养和激素状况的严格调控以满足身体对脂质和能量储存的需求。本研究结果显示, CAP替代超过20%鱼粉会导致日本沼虾肝胰腺acc和fas相对表达水平升高, 这可能与血淋巴内总甘油三酯和总胆固醇降低有关, 过低的血淋巴脂质水平会自动反馈给机体需要提高脂质合成能力。这与吉富罗非鱼的研究结果相一致。当饲料中CAP水平超过100 g/kg时, fas基因表达水平会上调, 吉富罗非鱼的脂质合成能力得到了提高[4]。而在大口黑鲈中则相反, CAP的添加则减少了脂肪的合成, 使脂肪合成和代谢达到了平衡水平, 减少内源性脂肪酸的合成来预防脂肪的过度积累, 从而达到抑制炎症的效果[2]。elovl6基因是超长链脂肪酸延伸酶 (ELOVLs)家族成员之一, 具有催化饱和和单不饱和脂肪酸的延伸, 它的表达主要受转录因子和一些激素的调控, 是动物机体内脂肪酸代谢的关键表达调控基因之一, 也是一种重要的长链脂肪酸延伸和合成反应的限速酶[25]。在本研究中, elovl6的表达量在CAP替代20%和40%鱼粉时升高, 但与对照组差异不显著。激素敏感性脂肪酶(HSL)是一种中性脂肪酶, 是脂肪分解的限速酶之一[26]。清道夫受体B类I型(SR-BI), 是CD36超家族的一员, 包括参与哺乳动物和鱼类脂质稳态调节的跨膜蛋白。SR-BI可以与低密度脂蛋白、极低密度脂蛋白高密度脂蛋白结合, 选择性摄取胆固醇酯, 进而调节细胞胆固醇稳态、双向胆固醇流动等功能[27]。日本沼虾肝胰腺sr-bI基因的表达与饲料脂肪源相关[27]。在本试验中, CAP替代20%鱼粉时, 日本沼虾肝胰腺sr-bI和hsl基因表达显著升高, 说明CAP替代鱼粉后对日本沼虾肝胰腺的脂代谢也产生了一定的影响, 但具体的影响机制仍需要进一步研究。
4. 结论
综上所述, CAP作为新型的单细胞蛋白源, 在日本沼虾饲料中替代20%鱼粉时, 对日本沼虾生长无显著负面影响, 同时降低日本沼虾血淋巴总胆固醇和甘油三酯, 提高血淋巴葡萄糖水平。CAP替代20%鱼粉对日本沼虾生长无显著负面影响的可能原因在于其提高了肝胰腺tor基因的表达量。总之, 日本沼虾可以利用CAP作为蛋白源, 适量使用对日本生长、蛋白及脂肪代谢无影响。
-
![]()
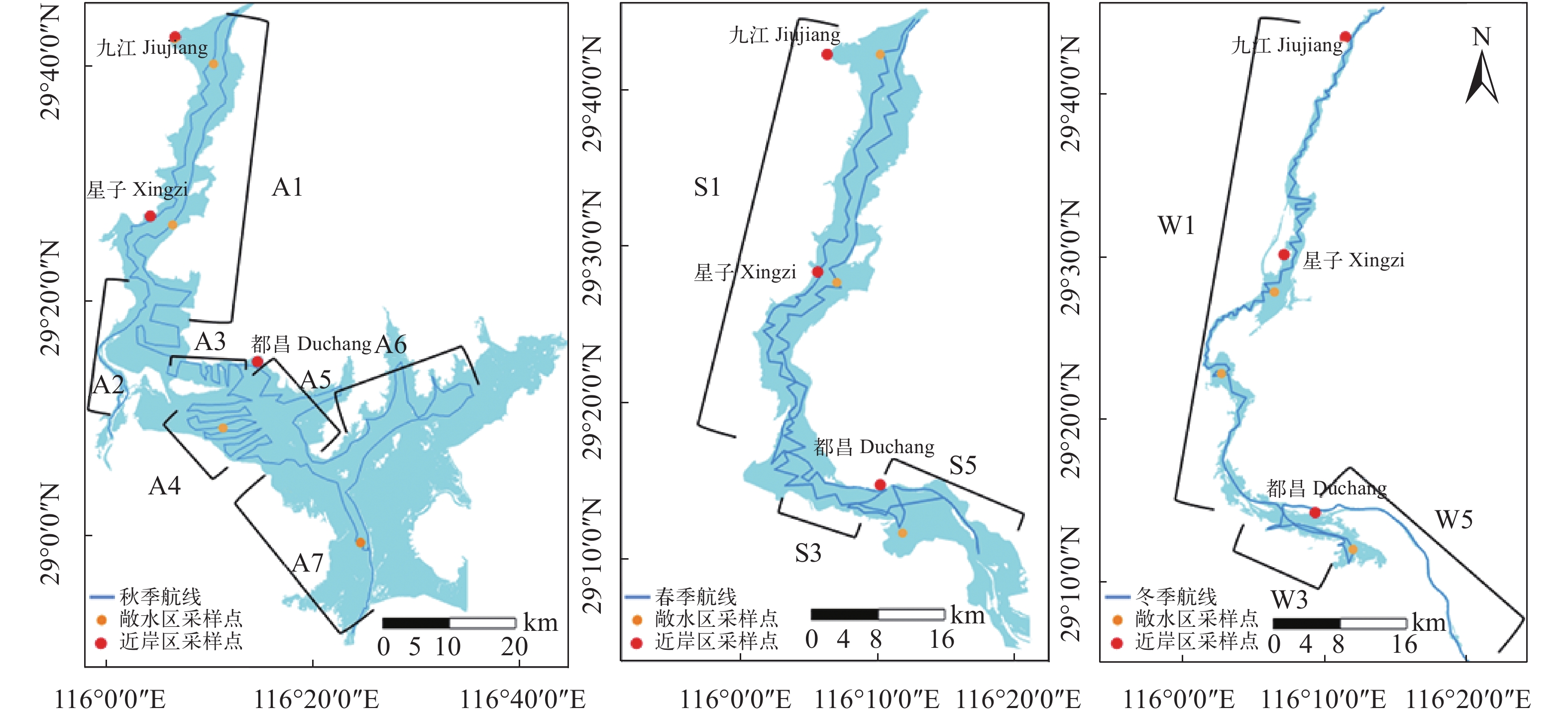
图 1 鄱阳湖水声学探测航线及渔获物采集点示意图
Figure 1. Routes of hydroacoustic survey and fish sampling sites in Poyang Lake
![]()
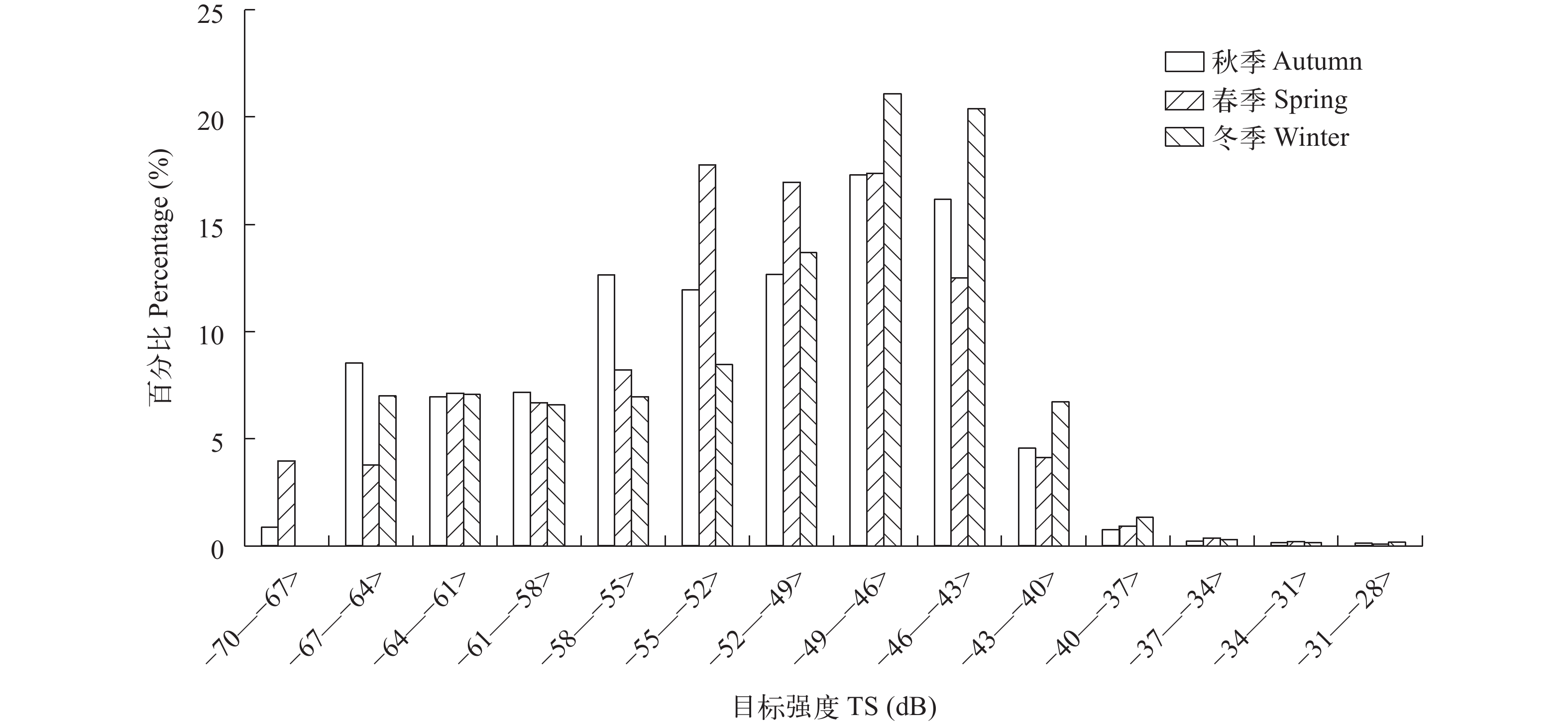
图 2 秋季、春季和冬季鱼类目标强度值分布
Figure 2. Distribution diagram of fish target strength among seasons
![]()
图 3 秋季、春季和冬季鱼类平均目标强度值比较
Figure 3. Distribution diagram of the mean fish target strength among seasons
![]()
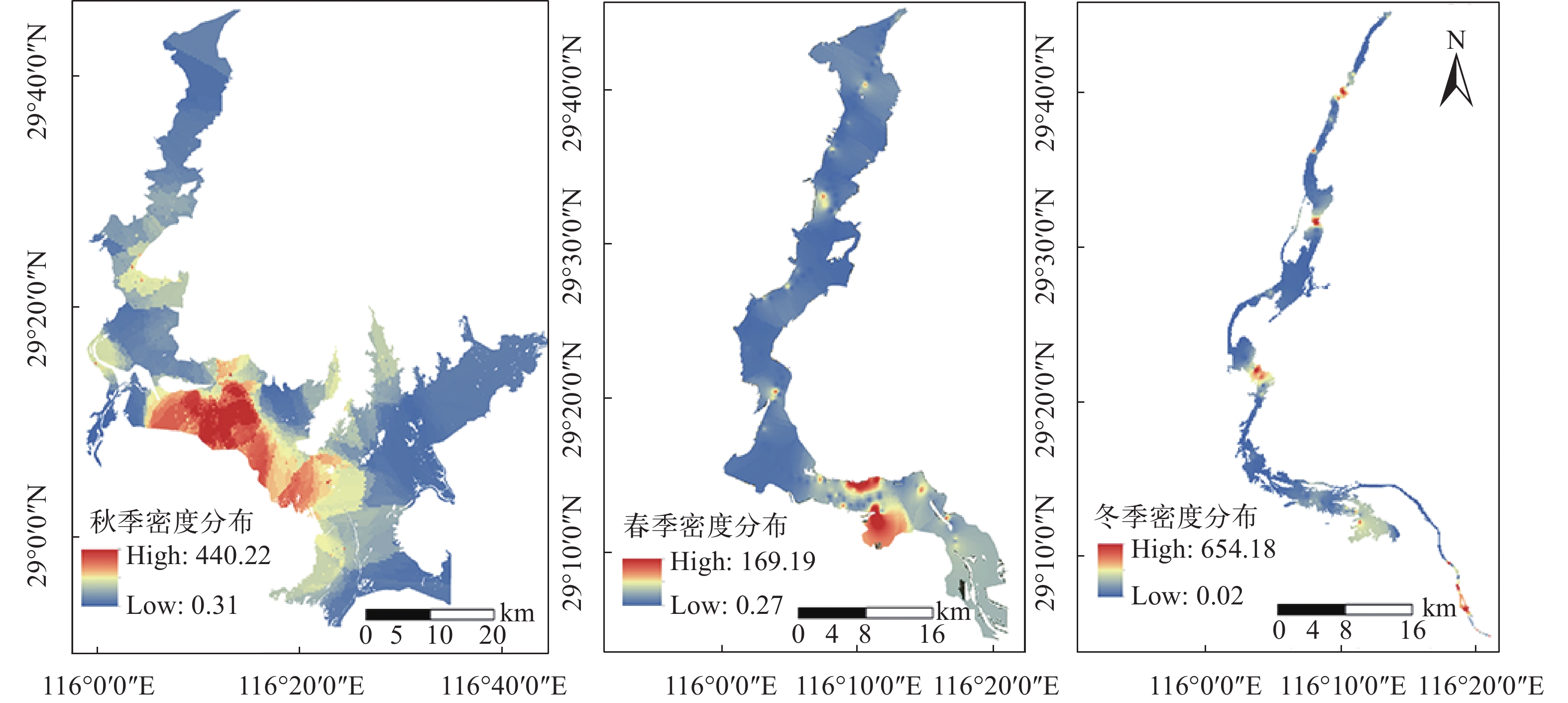
图 4 鄱阳湖三个季节鱼类密布水平分布
Figure 4. The horizontal distribution of fish density across Poyang Lake
![]()
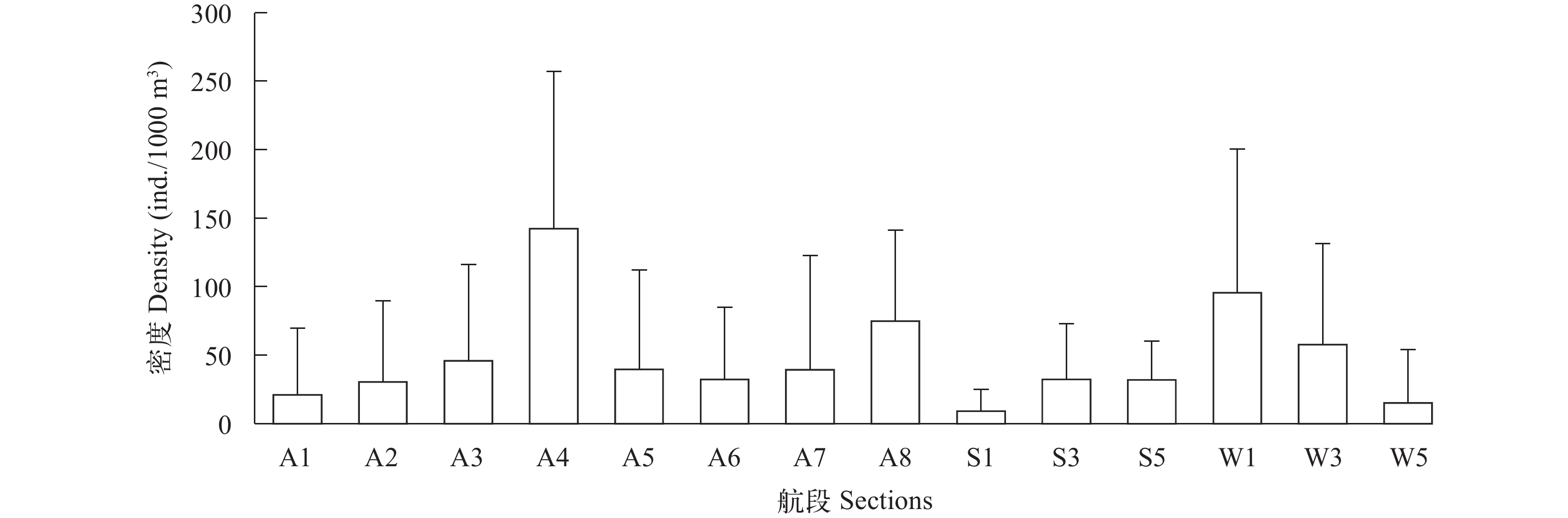
图 5 秋季、春季和冬季各航段鱼类密度分布
Figure 5. Horizontal density distribution of fish in sections of seasons
表 1 鄱阳湖水声学调查的基本信息
Table 1 Basic information of hydroacoustic survey in Poyang Lake
季节
Season调查时间
Date调查区域
Survey area航段
Sections航程
Survey distance
(km)平均水深
Average water
depth (m)温度
Temperature (℃)秋季Autumn 2020年
10月10日至10月27日通江水道 A1 98.31 12.40 19.94 赣江北支 A2 45.89 13.53 20.2 松门山北部 A3 58.41 11.06 19.85 松门山南部 A4 90.37 10.62 19.3 和合乡-周溪镇 A5 41.62 9.39 19.97 周溪镇-万户镇 A6 95.61 8.74 20.1 三江口-周溪镇 A7 40.12 9.20 19.25 三江口-南矶山 A8 19.21 9.16 19.67 冬季Winter 2021年
1月15日至1月30日通江水道 W1 127.95 8.61 7.76 松门山北部 W3 87.91 7.92 7.22 和合乡-周溪镇 W5 49.62 7.16 7.57 春季Spring 2021年
4月15日至4月27日通江水道 S1 123.93 10.22 18.54 松门山北部 S3 55.20 9.63 19.01 和合乡-三江口 S5 26.62 8.50 18.95  下载: 导出CSV
下载: 导出CSV
附表 1 调查期间湖口水位
Appendix 1 The water level during field survey time
日期Date 湖口水位Water level (m) 日期Date 湖口水位Water level (m) 2020年10月10号 17.30 2021年01月20号 8.98 2020年10月11号 17.25 2021年01月21号 8.9 2020年10月12号 17.14 2021年01月22号 8.82 2020年10月13号 17.01 2021年01月23号 8.74 2020年10月14号 16.94 2021年01月24号 8.62 2020年10月15号 16.83 2021年01月25号 8.53 2020年10月16号 16.75 2021年01月26号 8.42 2020年10月17号 16.69 2021年01月27号 8.30 2020年10月18号 16.69 2021年01月28号 8.22 2020年10月19号 16.65 2021年01月29号 8.15 2020年10月20号 16.59 2021年01月30号 8.10 2020年10月21号 16.50 2021年04月15号 12.29 2020年10月22号 16.44 2021年04月16号 12.46 2020年10月23号 16.32 2021年04月17号 12.62 2020年10月24号 16.22 2021年04月18号 12.72 2020年10月25号 16.11 2021年04月19号 12.85 2020年10月26号 15.98 2021年04月20号 12.84 2020年10月27号 15.85 2021年04月21号 12.77 2021年01月15号 9.23 2021年04月22号 12.60 2021年01月16号 9.27 2021年04月23号 12.77 2021年01月17号 9.32 2021年04月24号 12.82 2021年01月18号 9.21 2021年04月25号 12.86 2021年01月19号 9.08 2021年04月26号 13.08 2021年01月20号 8.98 2021年04月27号 13.31
下载: 导出CSV
附表 2 鄱阳湖渔获物鱼类种类组成
Appendix 2 The species composition and ecotypes in Poyang Lake
目Order 科Family 属Genus 种Species 空间
生态位
Space niche春
Spring秋
Autumn冬
Winter鲤形目Cypriniformes 鲿科Bagridae 黄颡鱼属Pelteobagrus 光泽黄颡鱼Pelteobagrus nitidus 下层 √ √ √ 黄颡鱼Pelteobagrus fulvidraco 下层 √ √ √ 瓦氏黄颡鱼Pelteobagrus vachelli 下层 √ √ √ 花鳅科Cobitidae 副沙鳅属Parabotia 花斑副沙鳅Parabotia fasciata 下层 — √ — 泥鳅属Misgurnus 泥鳅Misgurnus anguillicaud-atus 下层 √ √ √ 鲤科Cyprinidae 鳊属Parabramis 鳊Parabramis pekinensis 中下层 √ √ √ 鲌属Chanodichthys 蒙古鲌Culter mongolicusBasilewsky 上层 √ √ √ 䱗属Hemiculter 贝氏䱗Hemiculter bleekeri 下层 √ √ √ 䱗Hemiculter leucisculus 中上层 √ √ √ 草鱼属Ctenopharyngodon 草鱼Ctenopharyngodon idellus 中上层 √ √ √ 赤眼鳟属Squaliobarbus 赤眼鳟Squaliobarbus curriculus 中下层 √ √ √ 鲂属Megalobrama 鲂Megalobrama skolkovii 中上层 √ √ √ 鳡属Elopichthys 鳡Elopichthys bambusa 中下层 √ √ √ 鲴属Xenocypris 黄尾鲴Xenocypris davidi 中下层 √ √ √ 红鳍鲌属Chanodichthys 达氏鲌Culter dabryi 上层 √ √ √ 䱻属Hemibarbus 花䱻Hemibarbus maculatus 中下层 √ √ √ 鲤属Cyprinus 鲫Carassius auratus 中下层 √ √ √ 鲤Cyprinus carpio 中下层 √ √ √ 鲢属Hypophthal-michthys 鲢Hypophthalmichthys molitrix 中上层 √ √ √ 麦穗鱼属Pseudorasbora 麦穗鱼Pseudorasbora parva 中上层 √ √ √ 鳑鲏属Rhodeus 高体鳑鲏Rhodeus ocellatus 中上层 √ √ √ 中华鳑鲏Rhodeus sinensis 中下层 √ √ √ 飘鱼属Pseudolaubuca 寡鳞飘鱼Pseudolaubuca engraulis 上层 √ √ √ 翘嘴鲌属Culterilishaeformis 翘嘴鲌Culter alburnus 中上层 √ √ √ 青鱼属Mylopharyngodon 青鱼Mylopharyngodon piceus 中层 √ √ √ 蛇属SaurogobioBleeker 蛇Saurogobio dabryi 中下层 √ √ √ 鳙属Aristichthys 鳙Aristichthys nobilis 上层 √ √ √ 鱊属Acheilognathus 大鳍鱊Acheilognathus macropterus 中上层 √ √ √ 原鲌属Cultrichthys 红鳍原鲌Chanodichthys erythropterus 中下层 √ √ √ 鳅科Cobitidae 沙鳅属Botia 中华沙鳅Botia superciliaris 下层 √ √ √ 鲈形目Perciformes 鳢科Channidae 鳢属Channa 乌鳢Channa argus 下层 √ √ √ 鮨鲈科Percichthyidae 鳜属Siniperca 斑鳜Siniperca scherzeri 下层 √ √ — 大眼鳜Siniperca kneri 下层 √ √ √ 鳜Siniperca chuatsi 下层 √ √ √ 塘鳢科Eleotridae 沙塘鳢属Odontobutis 沙塘鳢Odontobutis obscura 下层 — √ — 虾虎鱼科Cobitidae 吻虾虎鱼属Rhinogobius 子陵吻虾虎鱼Ctenogobius giurinus 下层 √ √ — 鲇形目Siluriformes 鲇科Siluridae 鲇属Silurus 南方鲇Silurus meridionalis 下层 √ √ √ 鲇Silurus asotus 下层 √ √ √ 颌针鱼目Beloniformes 青鳉科Oryziatidae 青鳉属Oryzias 青鳉Oryzias latipes 中上层 — √ √ 鱵科Hemirhamphidae 下鱵属GenusHyporham-phus 间下鱵Hyporhamphus intermedius 中上层 — √ √ 鲱形目Clupeiformes 鳀科Engraulidae 鲚属Coilia 短颌鲚Coilia brachygnathus 中上层 √ √ √ 合鳃鱼目Synbranchiformes 刺鳅科Mastacembelidae 中华刺鳅属Sinobdella 中华刺鳅Sinobdella sinensis 下层 √ √ √
下载: 导出CSV
表 2 秋季、春季和冬季之间各水层鱼类密度差异性比较
Table 2 Comparison of fish density differences among different water layers in seasons
季节
Season水层
Layer平均密度
Average density最大密度
Maximum density最小密度
Minimum densityP值
P value密度比
Ratio (%)2020年秋
Autumn 2020表层Surface 18.63 289.64 0 <0.01 0.46 中层 Middle 40.29 1137.85 0 表层 Surface 18.63 289.64 0 <0.01 0.31 底层 Bottom 59.72 1166.29 0 中层 Middle 40.29 1137.85 0 <0.01 0.67 底层 Bottom 59.72 1166.29 0 2020年冬
Winter 2020表层 Surface 5.03 465.83 0 0.644 0.32 中层 Middle 15.76 502.53 0 表层 Surface 5.03 465.83 0 <0.01 0.2 底层 Bottom 24.59 496.32 0 中层 Middle 15.76 502.53 0 <0.01 0.64 底层 Bottom 24.59 496.32 0 2021年春
Spring 2021表层 Surface 18.08 226.78 0 <0.01 0.94 中层 Middle 19.33 268.67 0 表层 Surface 18.08 226.78 0 <0.01 3.02 底层 Bottom 5.99 107.35 0 中层 Middle 19.33 268.67 0 <0.01 3.23 底层 Bottom 5.99 107.35 0
下载: 导出CSV
-
[1] 张堂林, 李钟杰. 鄱阳湖鱼类资源及渔业利用 [J]. 湖泊科学, 2007, 19(4): 434-444. doi: 10.3321/j.issn:1003-5427.2007.04.012 Zhang T L, Li Z J. Fish resources and fishery utilization of Lake Poyang [J]. Journal of Lake Sciences, 2007, 19(4): 434-444. doi: 10.3321/j.issn:1003-5427.2007.04.012
[2] 杨少荣, 黎明政, 朱其广, 等. 鄱阳湖鱼类群落结构及其时空动态 [J]. 长江流域资源与环境, 2015, 24(1): 54-64. doi: 10.11870/cjlyzyyhj201501008 Yang S R, Li M Z, Zhu Q G, et al. Spatial and temporal variations of fish assemblages in Poyanghu Lake [J]. Resources and Environment in the Yangtze Basin, 2015, 24(1): 54-64. doi: 10.11870/cjlyzyyhj201501008
[3] 陈文静, 贺刚, 吴斌, 等. 鄱阳湖通江水道鱼类空间分布特征及资源量评估 [J]. 湖泊科学, 2017, 29(4): 923-931. Chen W J, He G, Wu B, et al. Spatial distribution and biomass assessment of fish in the channel connecting the Lake Poyang and the Yangtze River [J]. Journal of Lake Sciences, 2017, 29(4): 923-931.
[4] 连玉喜, 黄耿, Godlewska M, 等. 基于水声学探测的香溪河鱼类资源时空分布特征评估 [J]. 水生生物学报, 2015, 39(5): 920-929. doi: 10.7541/2015.121 Lian Y X, Huang G, Godlewska M, et al. Hydroacoustic assessment of spatio-temporal distribution and abundance of fish resources in the Xiangxi River [J]. Acta Hydrobiologica Sinica, 2015, 39(5): 920-929. doi: 10.7541/2015.121
[5] Appenzeller A R. Seasonal variability patterns of acoustic estimates of fish biomass and fish size in a lake dominated by coregonids (Lake Constance) [J]. Ergebnise Der Limno-logie, 1998(50): 227-236.
[6] 郭治之, 刘瑞兰. 江西鱼类研究 [J]. 南昌大学学报(理科版), 1995, 19(3): 222-232. Guo Z Z, Liu R L. The fishes of Jiangxi Province [J]. Journal of Nanchang University, 1995, 19(3): 222-232.
[7] 郭治之. 鄱阳湖鱼类调查报告 [J]. 江西大学学报, 1964(2): 121-130. Guo Z Z. Investigation report on fish in Poyang Lake [J]. Journal of Jiangxi University, 1964(2): 121-130.
[8] 伍汉霖, 邵广昭, 赖春福. 拉汉世界鱼类名典 [M]. 水产出版社, 1999: 1-1028. Wu H L, Shao G Z, Lai C F, et al. Latin-Chinese Dictionary of Fishes Names [M]. Taiwan: Fisheries Press, 1999: 1-1028.
[9] 褚新洛, 郑葆珊, 戴定远, 等. 中国动物志–硬骨鱼纲鲇形目. 北京: 科学出版社, 1999: 1-230. Chu X L, Zheng B S, Dai D Y, et al. Fauna Sinica Osteichthyes Siluriformes [M]. Beijing: Science Press, 1999: 1-230.
[10] 陈宜瑜等. 中国动物志-硬骨鱼纲. 鲤形目(中) [M]. 北京: 科学出版社, 1998: 1-531. Chen Y Y, et al. Fauna Sinica Osteichthyes Cypriniformes Ⅱ [M]. Beijing: Science Press, 1998: 1-531
[11] 乐佩奇等. 中国动物志-硬骨鱼纲. 鲤形目(下) [M]. 北京: 科学出版社, 2000: 1-661. Yue P Q, et al. Fauna Sinica Osteichthyes Cypriniformes Ⅲ [M]. Beijing: Science Press, 2000: 1-661.
[12] M]. Nanjing: Jiangsu Science Press, 1995: 1-549. [朱松泉. 中国淡水鱼类检索 [M]. 南京: 江苏科学技术出版社, 1995: 1-549. Zhu S Q. The Synopsis of Freshwater Fishes of China
[13] 陶江平, 龚昱田, 谭细畅, 等. 长江葛洲坝坝下江段鱼类群落变化的时空特征 [J]. 中国科学: 生命科学, 2012, 42(8): 677-688. Tao J P, Gong Y T, Tan X C, et al. Temporal and spatial characteristics of fish community changes in the lower reaches of Gezhouba Dam of the Yangtze River [J]. Scientia Sinica (Vitae)
, 2012, 42(8): 677-688. [14] Tan X, Kang M, Tao J, et al. Hydroacoustic survey of fish density, spatial distribution, and behavior upstream and downstream of the Changzhou Dam on the Pearl River, China [J]. Fisheries Science, 2011, 77(6): 891-901. doi: 10.1007/s12562-011-0400-5
[15] Aglen A. Random errors of acoustic fish abundance estimates in relation to the survey grid density applied [J]. Fao Fisheries Report, 1983.
[16] Godlewska M, Długoszewski B, Doroszczyk L, et al. The relationship between sampling intensity and sampling error-empirical results from acoustic surveys in Polish vendace lakes [J]. Fisheries Research, 2009, 96(1): 17-22. doi: 10.1016/j.fishres.2008.09.014
[17] Foote K G. Fish target strengths for use in echo integrator surveys [J]. The Journal of the Acoustical Society of America, 1987, 82(3): 981-987. doi: 10.1121/1.395298
[18] 陶江平, 艾为明, 龚昱田, 等. 采用渔业声学方法和GIS模型对楠溪江鱼类资源量及空间分布的评估 [J]. 生态学报, 2010, 30(11): 2992-3000. Tao J P, Ai W M, Gong Y T, et al. Assessment of fish abundance and distribution using fisheries acoustics and GIS modeling in the Nanxi River of Wenzhou City [J]. Acta Ecologica Sinica, 2010, 30(11): 2992-3000.
[19] 段辛斌, 谢意军, 郭杰等. 长江中游洪湖至宜昌江段鱼类空间分布特征的水声学研究 [J]. 长江流域资源与环境, 2016, 25(12): 1842-1849. doi: 10.11870/cjlyzyyhj201612007 Duan X B, Xie Y J, Guo J, et al. Hydroacoustic surveys on temporal and spatial distribution of fishes in the section from Honghu to Yichang of the Yangtze River middle reaches [J]. Resources and Environment in the Yangtze Basin, 2016, 25(12): 1842-1849. doi: 10.11870/cjlyzyyhj201612007
[20] 张俊, 陈丕茂, 陈国宝等. 基于Echoview声学数据后处理系统的背景噪声扣除方法 [J]. 渔业科学进展, 2014, 35(1): 9-17. doi: 10.3969/j.issn.1000-7075.2014.01.002 Zhang J, Chen P M, Chen G B, et al. Study on background noise removal based on Echoview acoustic data post-processing system [J]. Progress in Fishery Sciences, 2014, 35(1): 9-17. doi: 10.3969/j.issn.1000-7075.2014.01.002
[21] Szczucka J, Hoppe Ł, Schmidt B, et al. Acoustical estimation of fish distribution and abundance in two Spitsbergen fjords [J]. Oceanologia, 2017, 59(4): 585-591. doi: 10.1016/j.oceano.2017.04.007
[22] 蔡晓斌. 主被动遥感辅助下的鄱阳湖水位时空动态及洲滩变化研究 [D]. 武汉: 武汉大学, 2010: 90-114. Cai X B. The change analysis of water stage and beach wetland in Poyang Lake with the aid of multi-temporal active and passive remote sensing data [D]. Wuhan: Wuhan University, 2010: 90-114.
[23] Feng L, Hu C M, Chen X L, et al. MODIS observations of the bottom topography and its inter-annual variability of Poyang Lake [J]. Remote Sensing of Environment, 2011, 115(10): 2729-2741. doi: 10.1016/j.rse.2011.06.013
[24] 钱新娥, 黄春根, 王亚民, 等. 鄱阳湖渔业资源现状及其环境监测 [J]. 水生生物学报, 2002, 26(6): 612-617. doi: 10.3321/j.issn:1000-3207.2002.06.006 Qian X E, Huang C G, Wang Y M, et al. The status quo of fishery resources of Poyang Lake and its environmental monitoring [J]. Acta Hydrobiologica Sinica, 2002, 26(6): 612-617. doi: 10.3321/j.issn:1000-3207.2002.06.006
[25] 蒋祥龙, 黎明政, 杨少荣, 等. 鄱阳湖鱼类集合群落结构特征及其时间变化研究 [J]. 长江流域资源与环境, 2022, 31(3): 588-601. Jiang X L, Li M Z, Yang S R, et al. Temporal variation of fish metacommunity structure in Poyang Lake [J]. Resources and Environment in the Yangtze Basin, 2022, 31(3): 588-601.
[26] 陈文静, 张燕萍, 赵春来, 等. 近年长江湖口江段鱼类群落组成及多样性 [J]. 长江流域资源与环境, 2012, 21(6): 684-691. Chen W J, Zhang Y P, Zhao C L, et al. Species composition and biodiversity of fish community in Hukou section of the Yangtze River [J]. Resources and Environment in the Yangtze Basin, 2012, 21(6): 684-691.
[27] Holles S, Simpson S D, Radford A N, et al. Boat noise disrupts orientation behaviour in a coral reef fish [J]. Marine Ecology Progress Series, 2013(485): 295-300.
[28] 陈新军. 渔业资源与渔场学 [M]. 北京: 海洋出版社, 2004: 56-62. Chen X J. Fishery Resources and Fishery Science [M]. Beijing: Ocean Press, 2004: 56-62.
[29] 杜浩, 班璇, 张辉, 等. 天然河道中鱼类对水深、流速选择特性的初步观测——以长江江口至涴市段为例 [J]. 长江科学院院报, 2010, 27(10): 70-74. doi: 10.3969/j.issn.1001-5485.2010.10.015 Du H, Ban X, Zhang H, et al. Preliminary observation on preference of fish in natural channel to water velocity and depth: case study in reach of Yangtze River from Jiangkou town to Yuanshi town [J]. Journal of Yangtze River Scientific Research Institute, 2010, 27(10): 70-74. doi: 10.3969/j.issn.1001-5485.2010.10.015
[30] 罗卫. 鄱阳湖水沙时空变化遥感监测研究 [D]. 赣州: 江西理工大学, 2015: 60-64. Luo W. Poyang lake water-sediment change of time and space remote sensing monitoring research [D]. Ganzhou: Jiangxi university of science and technology, 2015: 60-64.
[31] Dronova I, Beissinger S, Burnham J et al. Landscape-level associations of wintering waterbird diversity and abundance from remotely sensed wetland characteristics of Poyang Lake [J]. Remote Sensing, 2016, 8(6): 462. doi: 10.3390/rs8060462
[32] 姜涛. 基于耳石形态和微化学特征的我国鲚属鱼类洄游生态学研究 [D]. 南京: 南京农业大学, 2014: 55-69. Jiang T. Study on migratory ecology of Coilia fish in Chinese waters inferred from otolith morphometric and microchemical characteristics [D]. Nanjing: Nanjing Agricultural University, 2014: 55-69.
[33] Chu D Z. Technology evolution and advances in fisheries acoustics [J]. Journal of Marine Science and Technology, 2011, 19(3): 245-252.
[34] Ye S W, Li Z J, Feng G P, et al. Length-weight relationships for thirty fish species in lake Niushan, a shallow macrophytic Yangtze lake in China [J]. Asian Fisheries Science, 2007(20): 217-226.
[35] Power M E. Predator Avoidance by Grazing Stream Fishes in Temperate and Tropical Streams: Importance of Stream Depth and Prey Size. Predation: Direct and Indirect Impacts in Aquatic Communities [M]. Hanover: University Press of New England, 1987: 333-351.
[36] 章守宇, 汪振华. 鱼类关键生境研究进展 [J]. 渔业现代化, 2011, 38(5): 58-65. doi: 10.3969/j.issn.1007-9580.2011.05.014 Zhang S Y, Wang Z H. Progress and current status of studies on essential fish habitat [J]. Fishery Modernization, 2011, 38(5): 58-65. doi: 10.3969/j.issn.1007-9580.2011.05.014
[37] 刘丹, 王烜, 李春晖, 等. 水文连通性对湖泊生态环境影响的研究进展 [J]. 长江流域资源与环境, 2019, 28(7): 1702-1715. Liu D, Wang X, Li C H, et al. Eco-environmental effects of hydrological connectivity on lakes: a review [J]. Resources and Environment in the Yangtze Basin, 2019, 28(7): 1702-1715.
[38] 王珂, 段辛斌, 刘绍平, 等. 三峡库区大宁河鱼类的时空分布特征 [J]. 水生生物学报, 2009, 33(3): 516-521. Wang K, Duan X B, Liu S P, et al. Survey on temporal and spatial distribution of fish in the Daning River [J]. Acta Hydrobiologica Sinica, 2009, 33(3): 516-521.
[39] 何学福, 邓其祥. 嘉陵江主要经济鱼类越冬场、产卵场、幼鱼索饵场调查及保护利用 [J]. 西南师范学院学报(自然科学版), 1979, 4(2): 27-41. He X F, Deng Q X. Investigation, protection and utilization of overwintering grounds, spawning grounds and juvenile feeding grounds of main economic fish in Jialing River [J]. Journal of Southwest China Normal University (
Natural Science Edition ) , 1979, 4(2): 27-41.


计量
- 文章访问数: 3641
- HTML全文浏览量: 475
- PDF下载量: 108