
EFFECTS OF RICE-FISH COCULTURE ON RICE YIELD: A META-ANALYSIS
-
摘要: 定量分析世界范围内稻鱼共作对水稻的产量效应, 为稻鱼共作技术的大面积推广应用提供科学依据。研究搜集了公开发表的102组稻鱼共作与水稻单作处理的水稻产量数据(截至2021年12月31日)。运用Meta分析方法, 明确了稻鱼共作对水稻产量的综合效应, 进而量化分析了时间区域、田间条件、水稻品种、肥药管理、鱼苗投放和投喂管理等对稻鱼共作产量效应的影响。与水稻单作相比, 稻鱼共作显著提高了水稻产量, 平均增产率约为17.2%(95%CI: 9.4%—25.6%), 分析结果可靠。不同区域气候稻鱼共作的产量效应差异显著, 国外和热带气候区域较高。2011年以后年份、田沟布局为一侧、水稻品种为粳稻的试验田中, 稻鱼共作的水稻增产率相对较高; 不同测产方式的产量效应差异不显著。不同肥药管理稻鱼共作的产量效应差异显著, 施肥3次、单一追肥、施用有机肥与无机肥、施药的产量效应较高; 在保障增产效应的前提下可以适当减少肥料农药的施用量, 采用有机肥和不施农药方式以获取经济与环境的双重效益。投苗时间、养殖模式、投苗密度、投喂与否稻鱼共作的产量效应差异显著, 在水稻移栽后21—25d投苗单养, 规格、密度及生物量控制在40 g/尾、1 尾/m2和30 g/m2以内, 不投喂饲料更能发挥稻鱼共作的增产效应。稻鱼共作能够显著提高水稻产量, 是有效利用稻田资源的可行方式。Abstract: In order to provide a scientific basis for the large-scale application of rice-fish coculture technology, the study quantitatively analyzed the rice yield effects of rice-fish coculture in the world. 102 groups of rice yield data from published related literatures were sorted (up to December 31, 2021). Using the meta-analysis method, the comprehensive effect of rice-fish coculture on rice yield was clarified. The effects among different time, regions, field conditions, rice varieties, management methods of fertilizer, pesticide, fry stocking and feeding on rice yield were further quantified. Compared with rice monoculture, rice-fish coculture significantly increased rice yield, with an average increase rate of 17.2% (95% CI: 9.4%—25.6%). The analysis results were reliable. The results of subgroup analysis showed that there was significant difference in yield effect between different regions and climates, no significant difference between different yield estimation methods, and the yield effect was higher in foreign and tropical climate region. The yield increase rate of rice-fish coculture was relatively higher in the experimental fields where experiment was conducted after 2011, field ditch was arranged on one side (L1) and rice variety was Japonica. The yield effects of rice-fish coculture among different management methods of fertilizer and pesticide were significantly different, it was higher when fertilization times was three, fertilization pattern was single top fertilizer (TF), fertilization variety was organic and inorganic fertilizer (OIF) and pesticide was applied. On the premise of guaranteeing the yield increase effect, the application amount of fertilizer and pesticide could be appropriately reduced. Applying organic fertilizer (OF) and no pesticide could obtain dual benefits of economy and environment. The yield effects of rice-fish coculture were significantly different among fry stocking days, aquaculture models, fry densities and feeding strategies. When fish of single variety (MC) was stocked 21 to 25 days after rice transplanting without feed and the size, density and biomass were controlled within 40 g/tail, 1 tail/m2 and 30 g/m2, the rice-fish coculture had better yield increase effect. Rice-fish coculture can significantly increase rice yield, which is a feasible way to effectively utilize rice field resources.
-
Keywords:
- Rice-fish coculture /
- Rice /
- Yield effect /
- Meta analysis
-
鳗鲡(Anguilla), 俗称河鳗, 又称白鳝、青鳝等, 属鳗鱼目、鳗鲡科鱼类, 广泛分布于热带、亚热带和温带地区; 其肉质鲜美, 营养丰富, 具有一定药膳价值, 是公认的水产珍品, 一直享有“水中人参”的美誉[1]。自20世纪90年代中期以来, 鳗鲡已经成为我国重要的水产养殖种类, 连续多年在单一出口创汇水产品品种中排名第一。当前我国鳗鲡的养殖主要分布在南方地区, 养殖品种以日本鳗鲡和美洲鳗鲡为主。其中, 日本鳗鲡多在广东省主要采用土池模式养殖, 美洲鳗鲡主要在福建省及其他省份采用水泥池精养模式, 一般在投喂前将粉状配合饲料制作成面团状饲料的过程中添加鱼油。
鱼油是鱼粉加工过程中产出的副产品, 富含二十碳五烯酸(EPA)和二十二碳六烯酸(DHA)酸等n-3高不饱和脂肪酸(HUFA)。研究表明, 鱼类饲料中添加适量鱼油, 可以满足生长所需的能量和必需脂肪酸, 提高饲料利用率, 其在维持细胞膜的流动性、增强免疫、提高抗氧化能力及调节脂类代谢等方面发挥着重要的作用[2-5]。因此, 富含HUFA的鱼油一直被作为水产动物饲料优质脂肪源, 尤其是鳗鲡养殖中使用的最主要脂肪来源。由于鱼油中HUFA含量高, 极易氧化变质, 往往在鳗鲡养殖场将粉状配合饲料加水制作成面团状饲料时直接添加。目前, 已有日本鳗鲡(Anguilla japonica)[6]、欧洲鳗鲡(Anguilla anguilla)[7]、澳洲鳗鲡(Auguilla australis)[8]及花鳗鲡(Anguilla marmorata)[9]饲料中适宜脂肪水平的报道。但美洲鳗鲡(Anguilla rostrata)饲料中适宜的鱼油添加水平还未确定, 也未见饲料中适宜脂肪水平的报道。因此, 本试验以我国主要的鳗鲡养殖品种-美洲鳗鲡幼鱼为试验对象, 通过研究饲料中添加不同水平鱼油对其生长性能、体成分及肝脏脂肪代谢酶的影响, 确定适宜的鱼油添加水平, 为鱼油在美洲鳗鲡养殖中的科学应用提供参考。
1. 材料与方法
1.1 试验动物与试验设计
试验用鱼为美洲鳗鲡幼鱼。挑选800尾规格一致、体格健壮, 平均体重为(8.34±0.12) g的试验鱼, 随机分为5个处理组, 分别投喂基础饲料中添加0(对照组)、3%(FO3组)、6%(FO6组)、9%(FO9组)和12%(FO12组)鱼油的试验饲料, 每组4个重复, 每个重复40尾鱼。试验期56d。
1.2 试验饲料与饲养管理
基础饲料为福建正源饲料有限公司生产的黑仔鳗商品饲料, 主要营养成分为粗蛋白47.60%、粗脂肪5.60%、粗灰分12.90%和粗纤维0.40%。鱼油购于福州开发区高龙实业有限公司。试验在福建正源饲料有限公司养殖实验室进行, 1200尾试验鱼随机分到20个PVC养殖缸(注水量为250 L, 每缸60尾)中暂养2周后, 选择出规格接近的800尾用于养殖试验, 每缸放鱼40尾。期间将基础饲料按料水重量比1﹕1.2混合制作成团状饲料投喂。正式试验每次投喂时, 先将粉状商品饲料称重, 再分别与饲料重量0%、3%、6%、9%和12%的鱼油, 及饲料重量1.2倍左右的水一起混合均匀制成团状饲料投喂。每天饱食投喂2次(5: 00和18: 00), 根据鱼体重、摄食情况及时调整投喂量。投喂30min 后, 将残饵吸出, 收集并记录残饵重。在试验期间, 每日检测水质、水温和记录试验鱼的摄食情况等, 保持水温26—28℃, 溶解氧浓度>7.6 mg/L, pH为7.8—8.2, 氨氮浓度<0.25 mg/L。
1.3 样品采集与组织匀浆液的制备
在试验结束后, 美洲鳗鲡幼鱼禁食24h, 将每缸鱼捞出称总重并记录尾数, 用丁香酚和无水乙醇按比例1﹕4配置成50 mg/L的混合液麻醉5min左右, 每缸随机取6尾鱼, 用纱布擦拭鱼体表面水分。在冰浴上用无菌剪刀解剖分别取出肠道和肝脏, 用生理盐水漂洗装入灭菌的冻存管中, 液氮速冻, 保存于‒80℃冰箱。用于测定肠道消化酶和肝脏脂肪代谢酶指标分析。将保存于–80℃冰箱中美洲鳗鲡幼鱼肠道和肝脏取出, 在冰上解冻。按组织重(g)与0.86%生理盐水(mL)体积比1﹕9放入10 mL离心管中, 用组织研磨仪(Tissuelyser-24, 上海净信实业发展有限公司, 中国)在冰浴条件下匀浆(匀浆时间30s/次, 间隙20s, 连续3—5次), 将制备好的组织匀浆用4℃离心机离心(3000 r/min, 10min), 取上清液分装到离心管中备用。每缸另随机取5尾鱼(去除内脏)装入标记分组自封袋中, 置于‒20℃保存, 用于体成分的测定。
1.4 测定指标及方法
生长性能指标 生长性能指标计算公式如下:
增重率 (Weight gain rate, WGR, %)=100%×(Wt–W0)/W0
投饵率 (Feeding rate, FR, %)=100%×FC/[W0+Wt/2]/t
饲料效率 (Feed efficiency, FE, %)=100%×(Wt–W0)/FC
成活率 (Survival rate, SR, %)=100%×Nf/Ni
式中, W0 (g)为平均每尾鱼初始体重; Wt (g)为平均每尾鱼末体重; FC (g)为平均每尾鱼摄食饲料总量(风干样重); t (d)为饲喂天数; Ni为初始鱼尾数; Nf为试验终末鱼尾数。
肠道消化酶活性 肠道蛋白酶、淀粉酶和脂肪酶的活性均使用南京建成生物工程研究所生产的试剂盒进行测定, 操作步骤均按说明书进行。
全鱼体成分 试验鱼全鱼营养成分含量采用AOAC[10]的方法测定, 主要如下: 水分含量测定采用105℃烘干恒重法, 粗蛋白质含量的测定采用全自动定氮仪(Kjeltec8400, FOSS公司), 粗脂肪含量测定采用索氏提取法, 粗灰分含量测定采用550℃马弗炉灼烧法。
肝脏脂肪代谢酶水平或活性 脂肪酸合成酶(Fatty acid synthase, FAS)和乙酰辅酶A羧化酶(Acetyl CoA carboxylase, ACC)采用上海江莱生物科技有限公司生产的试剂盒测定; 脂蛋白酯酶(Lipoprotein lipase, LPL)和肝脏肝脂酶(Hepatic lipase, HL)活性采用南京建成生物工程研究所生产的试剂盒进行测定, 操作步骤均按说明书进行。
1.5 数据统计与分析
试验数据均以平均值±标准差(Mean±SD)表示, 用SPSS 22.0分析软件进行单因素方差分析(One-way ANOVA, LSD), 若存在显著差异, 则采用Duncan法进行多重比较, P<0.05表示差异显著。WGR、FE与饲料鱼油添加水平间的关系采用Linear Regression模块进行二次回归分析, 显著水平为P<0.05。
2. 结果
2.1 饲料中添加不同水平鱼油对美洲鳗鲡幼鱼生长性能的影响
由表 1可知, 饲料中添加鱼油显著影响美洲鳗鲡幼鱼WGR、FR和FE(P<0.05)。随着鱼油添加水平的增加, WGR、FR和FE均呈先上升后下降的趋势, FO6组上述指标数值均为最高。将WGR、FE与饲料中鱼油添加水平进行二次回归分析, 得出最佳WGR、FE时饲料中添加鱼油水平分别为6.78%和6.43%(图 1和图 2), 此时鳗鲡配合饲料中脂肪水平分别为11.59%和11.30%。试验期间, 每组鳗鲡的成活率为100%。
表 1 不同处理组美洲鳗鲡幼鱼生长性能Table 1. Growth performance of juvenile American eels in different treatment groups (mean±SD, n=4)项目Item 组别Group 对照组 FO3组 FO6组 FO9组 FO12组 初始体重IBW (g) 6.97±
0.656.67±
0.326.93±
0.157.03±
0.216.77±
0.40终末体重FBW (g) 10.36±
1.45a10.43±
0.76a15.43±
1.14c12.78±
1.11b11.18±
1.41ab增重率WGR (%) 48.36±
8.22a56.46±
10.14a122.36±
11.48c81.58±
12.35b64.78±
11.60ab投饵率FR (%) 0.77±
0.02a0.83±
0.06ab1.02±
0.11c0.94±
0.06bc0.94±
0.05bc饲料效率FE (%) 50.37±
5.86a53.04±
7.65a74.72±
5.16b61.36±
2.65a52.20±
9.36a成活率SR (%) 100 100 100 100 100 注: 同行数据肩标相同小写字母或无字母表示差异不显著(P>0.05), 不同小写字母表示差异显著(P<0.05)。下同Note: In the same row, vales with the same superscripts or no superscripts mean no significant difference (P>0.05), while vales with different superscripts mean significant difference (P<0.05). The same applies below ![]() 图 1 美洲鳗鲡幼鱼增重率与饲料中鱼油添加水平的关系Figure 1. Relationship between WGR and dietary fish oil supplementation level of juvenile American eels
图 1 美洲鳗鲡幼鱼增重率与饲料中鱼油添加水平的关系Figure 1. Relationship between WGR and dietary fish oil supplementation level of juvenile American eels![]() 图 2 美洲鳗鲡幼鱼饲料效率与饲料中鱼油添加水平的关系Figure 2. Relationship between FE and dietary fish oil supplementation level of juvenile American eels
图 2 美洲鳗鲡幼鱼饲料效率与饲料中鱼油添加水平的关系Figure 2. Relationship between FE and dietary fish oil supplementation level of juvenile American eels2.2 饲料中添加不同鱼油水平对美洲鳗鲡幼鱼肠道消化酶活性的影响
由表 2可知, 饲料中鱼油添加水平对美洲鳗鲡幼鱼肠道蛋白酶、脂肪酶和淀粉酶活性均有显著影响(P<0.05)。饲料中添加鱼油, 可显著降低肠道蛋白酶和淀粉酶活性(P<0.05), 不同鱼油添加组间两种酶活性无显著差异(P>0.05); 可显著提高肠道脂肪酶活性(P<0.05), FO6组、FO9组和FO12组间脂肪酶活性接近(P>0.05)。
表 2 不同处理组美洲鳗鲡幼鱼的肠道消化酶活性Table 2. Intestinal digestive enzyme activities of juvenile American eels in different treatment groups (mean±SD, n=4; U/mg prot)项目Item 组别Group 对照组 FO3组 FO6组 FO9组 FO12组 蛋白酶
Protease72.50±
9.42b48.28±
4.36a46.99±
3.77a41.55±
6.24a40.45±
5.60a脂肪酶
Lipase25.57±
3.80a35.70±
2.30b40.34±
3.21c41.61±
1.37c43.02±
2.33c淀粉酶
Amylase0.82±
0.07b0.56±
0.09a0.55±
0.19a0.40±
0.12a0.38±
0.08a2.3 饲料中添加鱼油水平对美洲鳗鲡幼鱼体成分的影响
由表 3可知, 饲料中添加鱼油显著影响美洲鳗鲡幼鱼全鱼粗蛋白质和粗脂肪含量(P<0.05)。随着饲料中鱼油添加水平的增加, 美洲鳗鲡全鱼粗蛋白质的含量呈逐渐下降趋势, FO12组数值上最低; 全鱼粗脂肪含量呈逐渐上升趋势, FO12组数值上最高。饲料中添加鱼油对全鱼水分和粗灰分的影响不显著(P>0.05)。
表 3 不同处理组美洲鳗鲡幼鱼的体成分Table 3. Body composition of juvenile American eels in different treatment groups (mean±SD, n=4; %)项目
Item组别 Group 对照组 FO3组 FO6组 FO9组 FO12组 水分Moisture 71.42±
0.1971.89±
0.3871.48±
0.3371.25±
0.2971.56±
0.37粗蛋白质Crude protein 17.63±
0.89b17.76±
0.26b16.59±
0.89ab16.53±
0.30ab15.86±
0.66a粗脂肪Crude lipid 6.61±
0.56a6.62±
0.32a8.42±
0.07b8.43±
0.18b8.52±
0.43b灰分Ash 2.78±
0.092.67±
0.102.57±
0.052.56±
0.042.65±
0.042.4 饲料中鱼油添加水平对美洲鳗鲡幼鱼肝脏脂肪代谢酶活性的影响
由表 4可知, 饲料中添加鱼油显著影响美洲鳗鲡幼鱼肝脏FAS、LPL和HL活性(P<0.05)。随着饲料中鱼油添加水平的增加美洲鳗鲡幼鱼肝脏FAS活性呈降低趋势, FO9组和FO12组FAS活性显著降低(P<0.05); LPL和HL活性呈升高趋势, 仅FO12组两种酶活性显著降低(P<0.05)。饲料中添加鱼油对ACC活性无显著影响(P>0.05)。
表 4 不同处理组美洲鳗鲡幼鱼的肝脏脂肪代谢酶水平或活性Table 4. Levels or activities of enzymes related to lipid metabolism in liver of juvenile American eels in different treatment groups (mean±SD, n=4)项目 Item 组别 Group 对照组 FO3组 FO6组 FO9组 FO12组 脂肪酸合成酶
FAS (ng/mg prot)39.42±
0.61b38.80±
0.78b39.77±
0.73b32.42±
0.22a32.26±
0.64a乙酰辅酶A羧化酶
ACC (ng/mg prot)0.27±
0.050.28±
0.020.29±
0.010.28±
0.010.26±
0.03脂蛋白酯酶
LPL (U/mg prot)0.67±
0.11a0.76±
0.10ab0.88±
0.19ab0.91±
0.15ab0.93±
0.18b肝脂酶
HL (U/mg prot)0.72±
0.15a0.74±
0.12a0.89±
0.08ab0.89±
0.09ab1.14±
0.29b3. 讨论
3.1 饲料中鱼油添加水平对美洲鳗鲡幼鱼生长性能的影响
目前还鲜见不同鱼油添加水平对鳗鲡生长性能影响的报道。在本试验中, 适量添加鱼油显著提高美洲鳗鲡幼鱼WGR、FR和FE, 且随饲料中鱼油添加水平增加, WGR、FR和FE呈先升高后下降的趋势。这种适量添加鱼油促进生长, 过量添加鱼油抑制生长的现象在斑石鲷(Oplegnathus punctatus)[11]、黄鳝(Monopterus albus)[12]、半滑舌鳎(Cynoglossus semilaevis)[13]、芙蓉鲤鲫(Furong crucian carp)[14]、斜带石斑鱼(Epinephelus coioides)[15]、异育银鲫(Carassius auratus gibelio)[16]、尼罗罗非鱼(Oreochromis niloticus)[17]和褐菖鲉(Sebastiscus marmoratus) [18]等研究中也有类似的报道。而在花鳗鲡[9]、三倍体虹鳟(Oncorhynchus mykiss)[19]、杂交黄颡鱼(Peltebagrus fulvidraco×P. vachelli)[20]、泰国草鱼(Puntius gonionotus)[21]、黄姑鱼(Nibea albiflora)[22]和大黄鱼(Larimichthys croceus)[23]等研究中, 随着饲料中鱼油添加水平升高, 鱼类生长未出现降低现象。导致这些差异的原因可能与试验鱼类品种、脂肪需要量、鱼油添加水平、鱼油组成、基础饲料脂肪水平、养殖环境等方面的差异有关, 具体情况还有待进一步查明。从上述不同鱼类添加鱼油的研究中可以看出, 饲料中适量添加鱼油大都促进鱼体生长, 可能与饲料中添加的鱼油为鱼类提供充足且高效的能量, 减少蛋白质供能的消耗, 发挥“脂肪节约蛋白质效应”有关[24-26]。
在本试验中, 美洲鳗鲡饲料中鱼油添加水平6.78%和6.43%时, WGR和FE达最高值。此时饲料中脂肪水平为11.59%和11.30%, 这与欧洲鳗鲡适宜脂肪水平为20%[7]、日本鳗鲡适宜脂肪水平为16%[6]和澳洲鳗鲡适宜脂肪水平为15%[8]相比, 美洲鳗鲡幼鱼适宜脂肪水平相对较低; 但高于花鳗鲡的8%适宜脂肪水平[27]。上述研究中不同品种鳗鲡饲料中适宜脂肪水平的变化情况与于海振[28]报道250 g、500 g两个规格不同品种鳗鲡肌肉中粗脂肪含量高低的顺序一致, 从高到低依次为欧洲鳗鲡>日本鳗鲡>美洲鳗鲡>花鳗鲡。可见, 鳗鲡饲料中适宜的脂肪水平与肌肉中粗脂肪含量密切相关。
此外, 与适宜鱼油添加水平组相比, 添加高水平的鱼油(9%和12%)显著抑制美洲鳗鲡生长性能。这种现象在斑石鲷[11]、黄鳝[12]、半滑舌鳎[13]、芙蓉鲤鲫[14]、斜带石斑鱼[15]、异育银鲫[16]、尼罗罗非鱼[17]和褐菖鲉[18]等饲料添加过量鱼油的研究中也有类似报道。研究表明, 鱼类与陆生动物一样为能而食, 其摄食量与饲料能量含量成反比; 当饲料脂肪水平升高时, 饲料的能量含量也相应增加, 鱼类的摄食量则减少, 降低了鱼类肠道对营养素的消化和吸收效率, 尤其是吸收过多的脂肪导致蛋白质利用率下降, 鱼类生长减缓[29]。这也说明鱼类对脂肪的利用有一定限度, 脂肪节约蛋白质的本质仅限于把蛋白质的分解供能降低到最低限度, 而对于蛋白质的其他功能则是脂肪无法替代[29, 30]。
3.2 饲料中鱼油添加水平对美洲鳗鲡幼鱼肠道消化酶活性的影响
在本试验中, 添加鱼油使美洲鳗鲡幼鱼肠道脂肪酶活性显著升高。这与斜带石斑鱼[15]、三倍体虹鳟[19]饲料中添加不同水平的鱼油研究结果一致。随着鱼油添加水平的上升美洲鳗鲡幼鱼肠道脂肪酶活性先升高后趋于相对稳定, 在斜带石斑鱼[15]、褐菖鲉[18]、巴丁鱼(Pangasianodon hypophthalmus)[31]和细鳞鲑 (Brachymystax lenok)[32]等饲料中添加不同水平鱼油后肠道脂肪酶活性也有类似的变化。而三倍体虹鳟随着鱼油添加水平增加肠道脂肪酶活性呈先升高后降低趋势[19]; 吉富罗非鱼饲料中添加过量鱼油水平可显著降低前、中肠脂肪酶活性[33]。本试验中添加较高鱼油水平未观察到美洲鳗鲡幼鱼脂肪酶活性受到明显抑制, 可能与鱼类对鱼油利用的能力、鱼油添加水平和脂肪需求量等方面的差异有关; 添加鱼油使美洲鳗鲡幼鱼肠道蛋白酶和淀粉酶活性降低, 与点篮子鱼(Siganus guttatus)饲料中添加鱼油后降低肠道蛋白酶和淀粉酶研究结果类似[34]; 在幼鱼和成鱼阶段石斑鱼添加9%和12%鱼油后降低肠道淀粉酶也有相同的结果[15]。然而不同鱼油添加水平间美洲鳗鲡幼鱼肠道蛋白酶和淀粉酶活性接近, 类似的结果在黄颡鱼[20]和丝尾鳠(Hemibagrus wyckioides)[35]饲料中添加鱼油研究中出现。这说明在一定鱼油水平基础上继续添加对鱼类肠道蛋白酶和淀粉酶活性影响不大。从添加鱼油对鱼类肠道消化酶影响的变化情况来看, 肉食性鱼类尤其是幼鱼阶段在脂肪源充足时, 优先利用脂肪作为能量来源, 通过提高脂肪酶的活力而增加对脂肪的消化吸收; 减少蛋白质氨基酸及碳水化合物作为能量的消耗, 相应降低肠道蛋白酶和淀粉酶的活性。
3.3 饲料中鱼油添加水平对美洲鳗鲡幼鱼体成分的影响
鱼体组成是反映鱼类生理状态和营养水平的重要指标。饲料中主要营养素水平的变化都会对鱼体组成产生影响。在本试验中, 添加6%、9%和12%鱼油显著提高美洲鳗鲡幼鱼全鱼粗脂肪含量, 而添加3%、6%和9%鱼油对全鱼粗蛋白质含量无显著影响。这些结果与尼罗罗非鱼[17]、泰国草鱼[21]、黄姑鱼[22]和大黄鱼[23]添加鱼油对全鱼体成分影响的报道相似, 说明在饲料中添加一定水平的鱼油可促进全鱼脂肪沉积[36], 而不影响鱼体中蛋白质的沉积。但本试验中仅添加12%的高水平鱼油显著降低美洲鳗鲡幼鱼全鱼粗蛋白含量, 与黑鲷(Acanthopagrus schlegelii)饲料中添加10%鱼油后全鱼粗蛋白含量变化相同[37], 说明添加过高鱼油添加可减少鱼体蛋白质的蓄积。
3.4 饲料中鱼油添加水平对美洲鳗鲡幼鱼肝脏脂肪代谢酶水平或活性的影响
FAS在鱼类肝脏合成脂肪酸的过程中起着重要作用[38], 其水平随着饲料脂肪水平的升高而降低[39]。在本试验中, 添加9%和12%水平的鱼油显著降低美洲鳗鲡幼鱼肝脏FAS水平, 与黑鲷添加10.0%鱼油[37]、道氏虹鳟添加10.95%和14.09%鱼油[40]和尼罗罗非鱼添加3.0%、6.0%、9.0%、12.0%和15.0%鱼油[17]后FAS水平降低的结果一致。研究表明, 鱼油富含EPA和DHA, 属于n-3系列不饱和脂肪酸,是FAS酶及相关基因的抑制剂[39, 41]。在道氏虹鳟添加7.86%、10.95%和14.09%鱼油[39], 斜带石斑鱼添加7.0%、10.5%和14.0%鱼油[42], 及大黄鱼添加12%鱼油也发现肝脏FAS基因表达丰度显著降低[43]。此外, 高水平n-3系列不饱和脂肪酸还可通过降低乙酰辅酶羟化酶和提高激素敏感脂肪酶的活性[44], 降低脂类合成相关基因和蛋白的表达水平[45], 从而抑制脂质合成。
LPL和HL是鱼类肝脏中参与脂肪分解代谢过程中的两个关键酶。LPL主要催化血浆中乳糜颗粒和极低密度酯蛋白, 将甘油三酯分解成脂肪酸和甘油[46]。HL主要参与高密度脂蛋白的重构和乳糜微粒残骸、低密度脂蛋白的代谢以及胆固醇的逆向转运[47]。在本试验中, 仅12%鱼油添加水平显著提高美洲鳗鲡幼鱼肝脏LPL和HL活性, 与道氏虹鳟添加10.95%和14.09%鱼油[40]、许氏平鲉(Sebastes schlegeli)添加9.0%和12.0%鱼油[48]、大菱鲆(Scophthalmus maximu)添加12.94%鱼油[49]、梭鱼(Chelon haematocheilus)添加12%和15%[26]和尼罗罗非鱼添加6.0%、9.0%、12.0%和15.0%[17]鱼油后, 肝脏LPL和HL活性与对照组相比的变化情况类似。而幼鱼和成鱼阶段石斑鱼添加高水平鱼油使这两种酶的活性显著降低[15]; 在尼罗罗非鱼鱼和多鳞鱚(Sillago sihama)等饲料中添加高水平鱼油也发现, LPL和HL活性与适宜鱼油添加水平组相比显著下降[17, 50]。这可能是不同品种鱼类在脂肪利用能力、脂肪需要量、鱼油添加水平和基础饲料脂肪水平等方面差异导致的。但总的来看, 添加一定水平的鱼油可提高肝脏脂肪分解酶的活性, 可能是饲料中添加鱼油使高水平的n-3多不饱和脂肪酸通过激活PPARɑ上调LPL靶基因表达所致[51]。鱼油调节鱼类肝脏HL活性的研究鲜见报道, 具体机制还有待进一步研究。
4. 结论
在本试验条件下, 饲料中适量添加鱼油可提高美洲鳗鲡幼鱼生长性能、肠道脂肪酶活性和全鱼粗脂肪含量, 调节肝脏脂肪代谢酶水平或活性; 美洲鳗鲡幼鱼饲料中适宜的鱼油添加水平推荐为6.43%—6.78%, 此时饲料中脂肪水平为11.30%—11.59%。
-
![]()
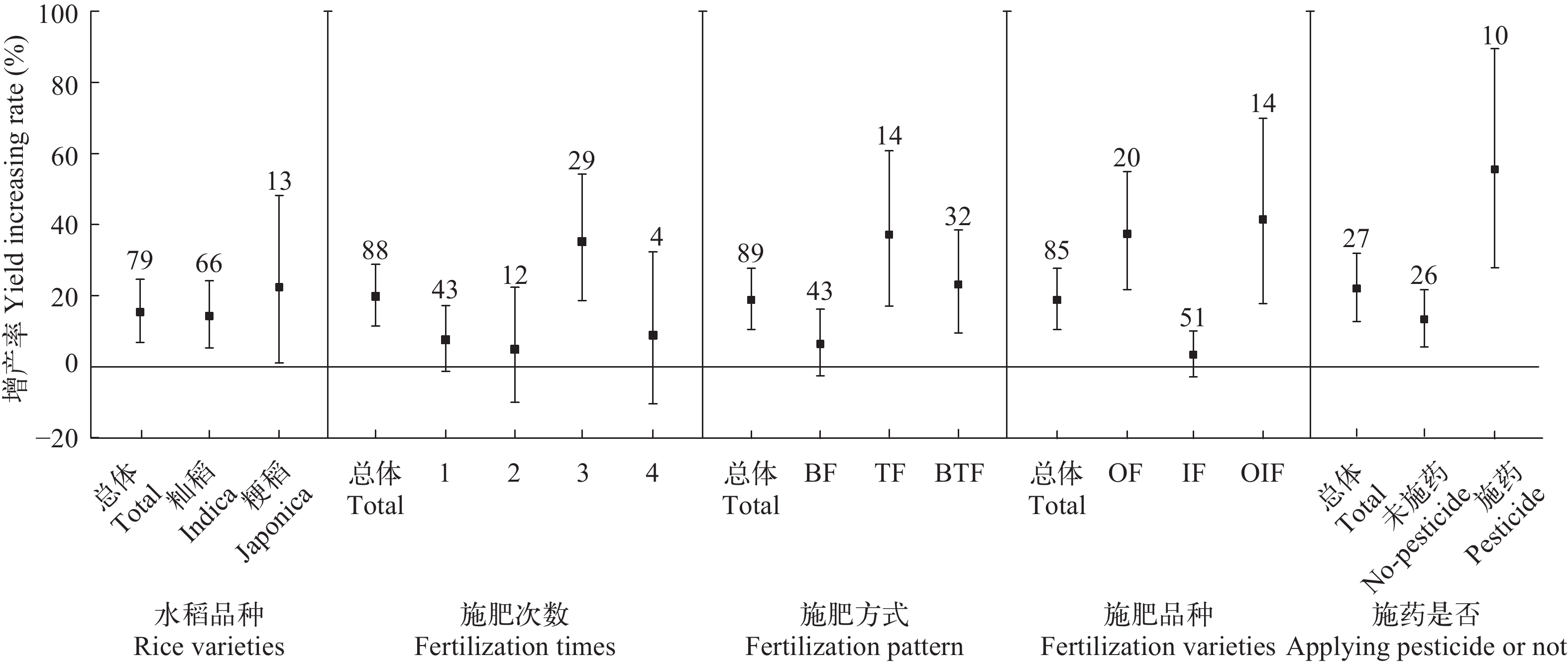
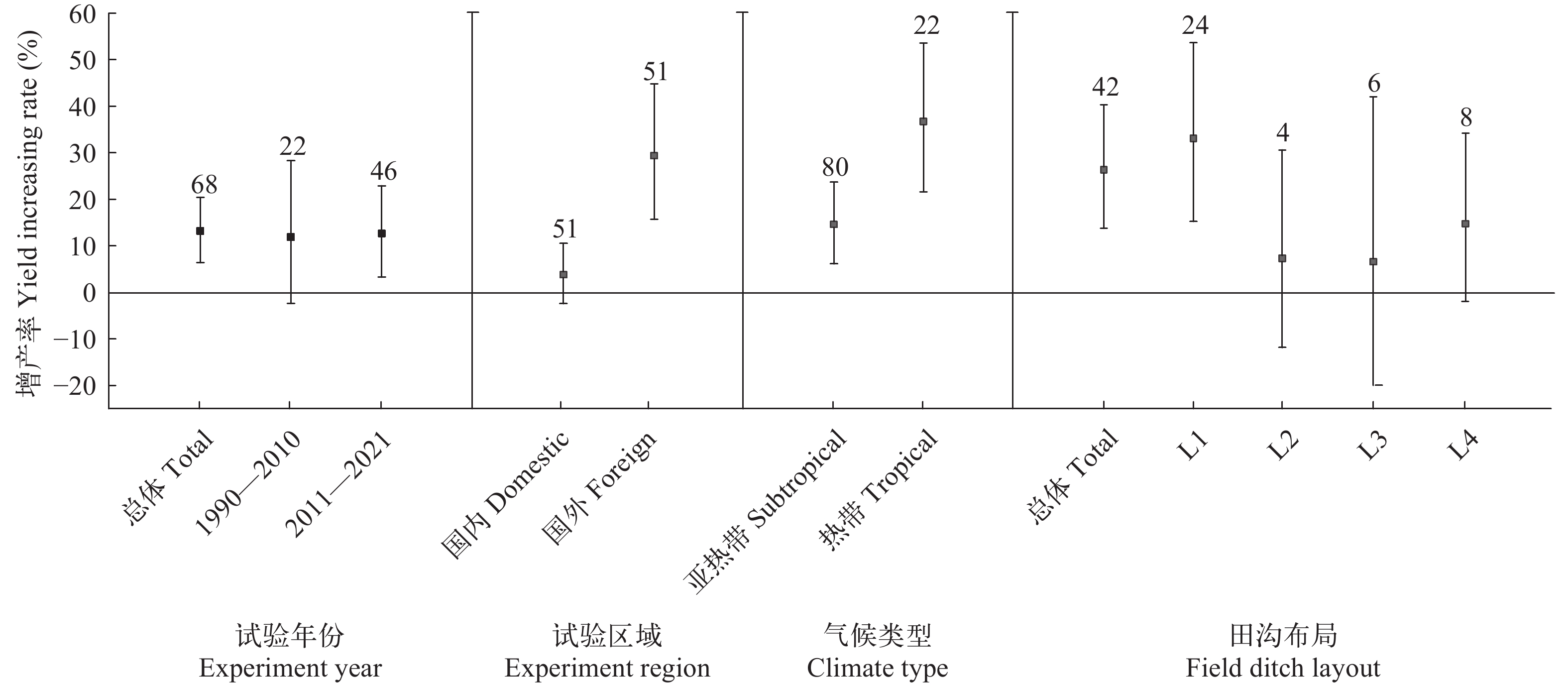
图 1 时间区域与田间条件对水稻产量的效应分析
Figure 1. Effect analysis of time, region and field conditions on rice yield
![]()
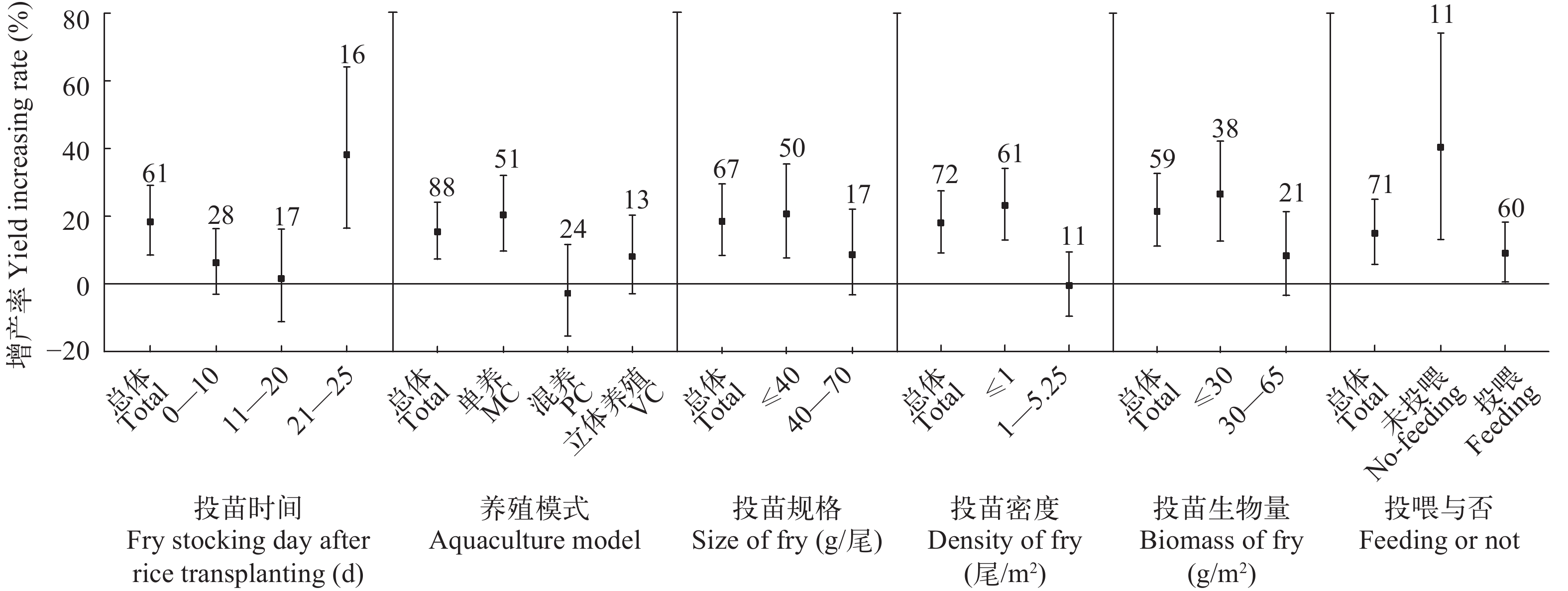
图 2 水稻品种与肥药管理对水稻产量的效应分析
Figure 2. Effect analysis of rice varieties, management of fertilizer and pesticide on rice yield
![]()
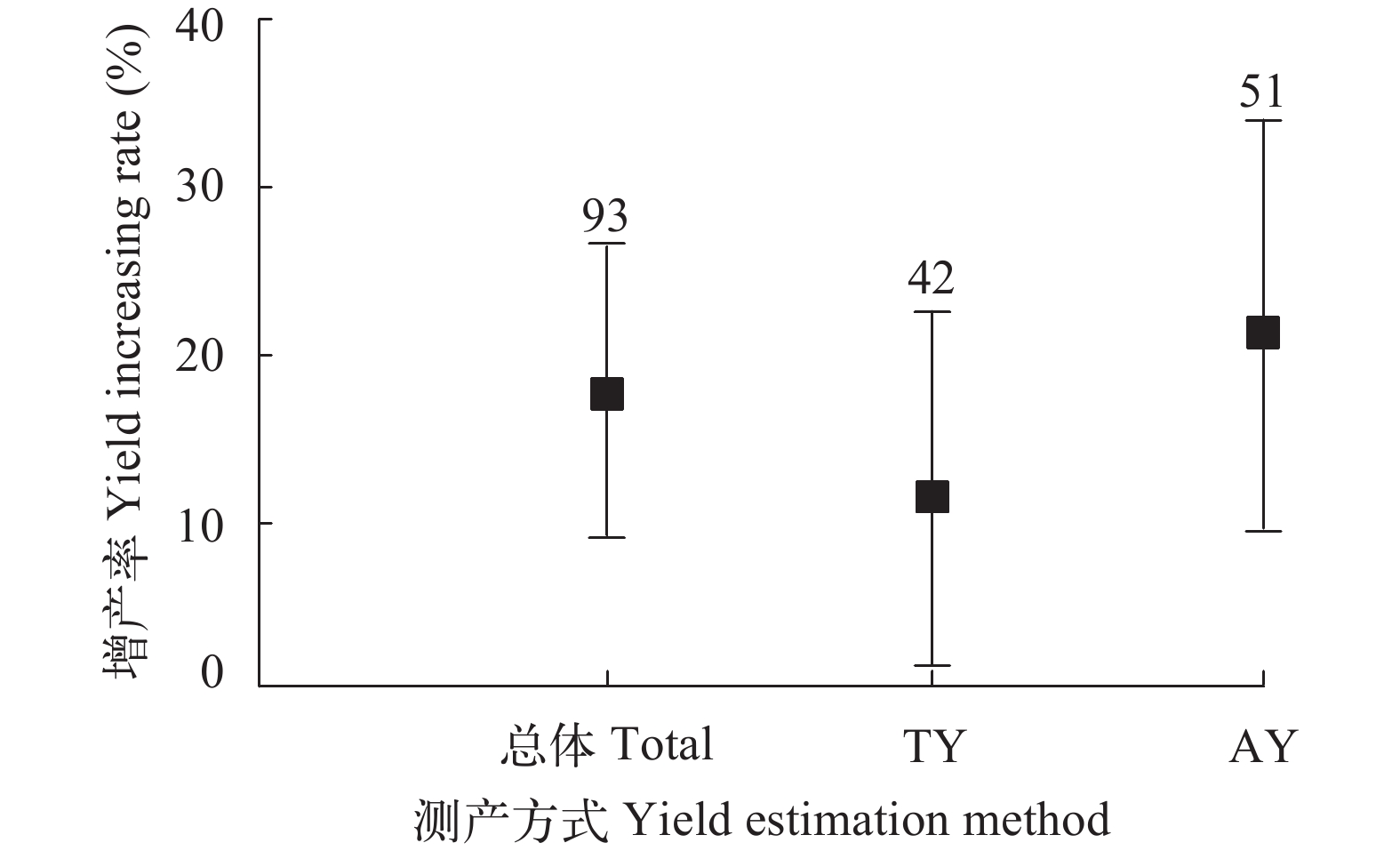
图 3 鱼苗投放与投喂管理对水稻产量的效应分析
Figure 3. Effect analysis of fry stocking and feeding management on rice yield
表 1 数据分组情况及Meta亚组分析结果
Table 1 Data grouping and results of sub-group meta-analysis
解释变量
Explaining variance分类亚组
Classification of sup-group异质性检验
Heterogeneity testQ P 时间区域与田间条件
Time, region and field conditions试验年份Experiment year 1990—2010、2011—2021 0.01 0.930 试验区域Experiment region 国内Domestic、国外Foreign 11.35 0.001* 气候类型Climate type 亚热带Subtropical、热带Tropical 6.15 0.013* 田沟布局Field ditch layout 一侧(L1)On one side、四周(L2)Around、内部(L3)Inside、四周与内部(L4)Around and inside 4.16 0.245 水稻品种与肥药管理Rice varieties, management of fertilizer and pesticide 水稻品种Rice varieties 籼稻Indica rice、粳稻Japonica rice 0.41 0.522 施肥次数Fertilization times 1、2、3、4 9.53 0.023* 施肥方式Fertilization pattern 单一基肥(BF)Base fertilizer、单一追肥(TF)Top fertilizer、基肥与追肥(BTF)Base and top fertilizer 9.02 0.011* 施肥品种Fertilization varieties 单一有机肥(OF)Organic fertilizer、单一无机肥(IF)Inorganic fertilizer、有机肥与无机肥(OIF)
Organic and inorganic fertilizer23.62 0.000* 施药与否Applying pesticide or not 未施药No-pesticide、施药Pesticide 9.20 0.002* 鱼苗投放与投喂管理Fry stocking and feeding management 投苗时间(移栽后天数)Fry stocking day after rice transplanting(d) 0—10、11—20、21—25 8.66 0.013* 养殖模式Aquaculture model 单养Monoculture(MC)、混养Polyculture(PC)、
立体养殖Vertical culture(VC)6.63 0.036* 投苗规格Size of fry (g/尾) ≤40、40—70 1.60 0.206 投苗密度Density of
fry (尾/m2)≤1、1—5.25 10.56 0.001* 投苗生物量Biomass of fry (g/m2) ≤30、30—65 3.52 0.060 投喂与否Feeding or not 未投喂No-feeding、投喂Feeding 4.61 0.032* 其他Other 测产方式Yield estimation method 理论产量(TY)Theoretical yield、实际产量
(AY)Actual yield1.40 0.237  下载: 导出CSV
下载: 导出CSV
表 2 稻鱼共作对水稻产量的综合效应值
Table 2 Comprehensive effect size of rice yield under rice-fish culture
模型
Model增产率
Rate (%)置信区间
CI (%)效应量检验
Effect size test异质性检验
Heterogeneity test发表偏倚检验
Publication bias test数据量
n下限LL 上限UL Z P Q P I2 (%) Nfs 剪补后After Trimfilling 增产率
Rate (%)置信区间CI (%) 下限LL 上限UL 随机效应模型REM 17.2 9.4 25.6 4.51 0.000 238.31 0.000 50.76 1243 30.2 22.1 38.8 102 Note: REM, CI, LL, UL represent Random effect model, confidence interval, lower limit and upper limit, respectively
下载: 导出CSV
-
[1] Khumairoh U, Lantinga E A, Handriyadi I, et al. Agro-ecological mechanisms for weed and pest suppression and nutrient recycling in high yielding complex rice systems [J]. Agriculture, Ecosystems & Environment, 2021(313): 107385.
[2] 朱昊俊, 强俊, 徐钢春, 等. 哈尼梯田稻-渔共作模式下杂交黄颡鱼肠道微生物研究 [J]. 水生生物学报, 2021, 45(6): 1232-1242. Zhu H Y, Qiang J, Xu G C, et al. Microbial community structure of hybrid yellow catfish in rice-fish co-culture system in Hani terrace [J]. Acta Hydrobiologica Sinica, 2021, 45(6): 1232-1242.
[3] 胡亮亮, 唐建军, 张剑, 等. 稻-鱼系统的发展与未来思考 [J]. 中国生态农业学报, 2015, 23(3): 268-275. Hu L L, Tang J J, Zhang J, et al. Development of rice-fish system: today and tomorrow [J]. Chinese Journal of Eco-Agriculture, 2015, 23(3): 268-275.
[4] Hu L L, Ren W Z, Tang J J, et al. The productivity of traditional rice-fish co-culture can be increased without increasing nitrogen loss to the environment [J]. Agriculture, Ecosystems & Environment, 2013(177): 28-34.
[5] 李伦, 罗强, 吴士龙, 等. 渔稻养作及其在涝渍地综合利用中的研究综述 [J]. 节水灌溉, 2016(5): 75-80. doi: 10.3969/j.issn.1007-4929.2016.05.017 Li L, Luo Q, Wu S L, et al. Review of integrated rice-fish farming and its comprehensive utilization in waterlogged land [J]. Water Saving Irrigation, 2016(5): 75-80. doi: 10.3969/j.issn.1007-4929.2016.05.017
[6] Gurung T, Wagle S. Revisiting underlying ecological principles of rice-fish integrated farming for environmental, economical and social benefits [J]. Our Nature, 2005, 3(1): 1-12.
[7] Tsuruta T, Yamaguchi M, Abe S I, et al. Effect of fish in rice-fish culture on the rice yield [J]. Fisheries Science, 2011, 77(1): 95-106. doi: 10.1007/s12562-010-0299-2
[8] Xie J, Hu L L, Tang J J, et al. Ecological mechanisms underlying the sustainability of the agricultural heritage rice-fish coculture system [J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(50): E1381-E1387.
[9] 梁玉刚, 李静怡, 王丹, 等. 垄作稻鱼鸡共生对水稻群体生长特性及产量形成的影响 [J]. 中国农业科技导报, 2020, 22(11): 165-175. Liang Y G, Li J Y, Wang D, et al. Impact of rice-fish-chicken ridge cultivation on rice population growth and yield [J]. Journal of Agricultural Science and Technology, 2020, 22(11): 165-175.
[10] 车阳, 程爽, 田晋钰, 等. 不同稻田综合种养模式下水稻产量形成特点及其稻米品质和经济效益差异 [J]. 作物学报, 2021, 47(10): 1953-1965. Che Y, Cheng S, Tian J Y, et al. Characteristics and differences of rice yield, quality, and economic benefits under different modes of comprehensive planting-breeding in paddy fields [J]. Acta Agronomica Sinica, 2021, 47(10): 1953-1965.
[11] 王旭东, 庄俊杰, 刘冰洋, 等. 秸秆还田条件下中国农田土壤有机碳含量变化及其影响因素的Meta分析 [J]. 中国农业大学学报, 2020, 25(8): 12-24. doi: 10.11841/j.issn.1007-4333.2020.08.02 Wang X D, Zhuang J J, Liu B Y, et al. Residue returning induced changes in soil organic carbon and the influential factors in China's croplands: a meta-analysis [J]. Journal of China Agricultural University, 2020, 25(8): 12-24. doi: 10.11841/j.issn.1007-4333.2020.08.02
[12] 姜玲玲, 刘静, 赵同科, 等. 有机无机配施对番茄产量和品质影响的Meta分析 [J]. 植物营养与肥料学报, 2019, 25(4): 601-610. doi: 10.11674/zwyf.18165 Jiang L L, Liu J, Zhao T K, et al. Meta-analysis of combinative application of organic and inorganic fertilizers on effect of yield and qualities of tomato [J]. Journal of Plant Nutrition and Fertilizers, 2019, 25(4): 601-610. doi: 10.11674/zwyf.18165
[13] 怀燕, 王岳钧, 陈叶平, 等. 稻田综合种养模式的化肥减量效应分析 [J]. 中国稻米, 2018, 24(5): 30-34. doi: 10.3969/j.issn.1006-8082.2018.05.006 Huai Y, Wang Y J, Chen Y P, et al. Chemical fertilizer reduction analysis of rice-based co-culture system [J]. China Rice, 2018, 24(5): 30-34. doi: 10.3969/j.issn.1006-8082.2018.05.006
[14] 周榆淇, 邹冬生, 王安岽, 等. 长期淹水条件下稻鱼复合种养对土壤养分和酶活性的影响 [J]. 农业现代化研究, 2022, 43(5): 1-10. doi: 10.13872/j.1000-0275.2022.0060 Zhou Y Q, Zou D S, Wang A D, et al. Effects of rice-fish integrated farming on soil nutrients and enzyme activities under long-term flooding [J]. Research of Agricultural Modernization, 2022, 43(5): 1-10. doi: 10.13872/j.1000-0275.2022.0060
[15] Vromant N, Chau N T, Ollevier F. The effect of rice seeding rate and fish stocking on the floodwater ecology of the rice field in direct-seeded, concurrent rice-fish systems [J]. Hydrobiologia, 2001, 445(1/2/3): 151-164.
[16] 曹志强, 梁知洁, 赵艺欣, 等. 北方稻田养鱼的共生效应研究 [J]. 应用生态学报, 2001, 12(3): 405-408. doi: 10.3321/j.issn:1001-9332.2001.03.021 Cao Z Q, Liang Z J, Zhao Y X, et al. Symbiotic effect of cultivating fish in rice field in North China [J]. Chinese Journal of Applied Ecology, 2001, 12(3): 405-408. doi: 10.3321/j.issn:1001-9332.2001.03.021
[17] 杨勇, 张洪程, 胡小军, 等. 稻渔共作水稻生育特点及产量形成研究 [J]. 中国农业科学, 2004, 37(10): 1451-1457. doi: 10.3321/j.issn:0578-1752.2004.10.008 Yang Y, Zhang H C, Hu X J, et al. Characteristics of growth and yield formation of rice in rice-fish farming system [J]. Scientia Agricultura Sinica, 2004, 37(10): 1451-1457. doi: 10.3321/j.issn:0578-1752.2004.10.008
[18] 袁伟玲, 曹凑贵, 李成芳, 等. 稻鸭、稻鱼共作生态系统CH4和N2O温室效应及经济效益评估 [J]. 中国农业科学, 2009, 42(6): 2052-2060. doi: 10.3864/j.issn.0578-1752.2009.06.022 Yuan W L, Cao C G, Li C F, et al. Methane and nitrous oxide emissions from rice-fish and rice-duck complex ecosystems and the evaluation of their economic significance [J]. Scientia Agricultura Sinica, 2009, 42(6): 2052-2060. doi: 10.3864/j.issn.0578-1752.2009.06.022
[19] Li T, Hasegawa T, Yin X, et al. Uncertainties in predicting rice yield by current crop models under a wide range of climatic conditions [J]. Global Change Biology, 2015, 21(3): 1328-1341. doi: 10.1111/gcb.12758
[20] Leigh C, Hiep L H, Stewart-Koster B, et al. Concurrent rice-shrimp-crab farming systems in the Mekong Delta: are conditions (sub) optimal for crop production and survival [J]? Aquaculture Research, 2017, 48(10): 5251-5262. doi: 10.1111/are.13338
[21] Zhao M, Xue K, Wang F, et al. Microbial mediation of biogeochemical cycles revealed by simulation of global changes with soil transplant and cropping [J]. The ISME Journal, 2014, 8(10): 2045-2055. doi: 10.1038/ismej.2014.46
[22] 吴雪, 谢坚, 陈欣, 等. 稻鱼系统中不同沟型边际弥补效果及经济效益分析 [J]. 中国生态农业学报, 2010, 18(5): 995-999. doi: 10.3724/SP.J.1011.2010.00995 Wu X, Xie J, Chen X, et al. Edge effect of trench-pond pattern on rice grain and economic benefit in rice-fish co-culture [J]. Chinese Journal of Eco-Agriculture, 2010, 18(5): 995-999. doi: 10.3724/SP.J.1011.2010.00995
[23] Halwart M, Borlinghaus M, Kaule G. Activity pattern of fish in rice fields [J]. Aquaculture, 1996, 145(1/2/3/4): 159-170.
[24] 龙瑞平, 杨兆春, 穆家伟, 等. 稻鱼共作下氮肥减量后移对水稻生长和氮肥利用效率的影响 [J]. 土壤, 2022, 54(4): 708-714. Long R P, Yang Z C, Mu J W, et al. Effects of postponing and decreasing nitrogen fertilizer on rice growth and nitrogen use efficiency under rice-fish coculture [J]. Soils, 2022, 54(4): 708-714.
[25] 成臣, 曾勇军, 王祺, 等. 氮肥运筹对南方双季晚粳稻产量及品质的影响 [J]. 植物营养与肥料学报, 2018, 24(5): 1386-1395. Cheng C, Zeng Y J, Wang Q, et al. Effects of nitrogen application regime on Japonica rice yield and quality of the late rice in the double rice system in Southern China [J]. Journal of Plant Nutrition and Fertilizers, 2018, 24(5): 1386-1395.
[26] 李文博. 稻田综合种养对水稻产量和品质的影响 [D]. 合肥: 安徽农业大学, 2021: 19. Li W B. Impacts of co-culture of rice and aquatic animals on rice yield and quality [D]. Hefei: Anhui Agricultural University, 2021.
[27] 倪达书, 汪建国. 论稻鱼共生生态系统的应用价值 [J]. 水产科技情报, 1983, 10(6): 1-4. Ni D S, Wang J G. Application value of rice-fish symbiosis ecosystem [J]. Fisheries Sicence & Technology Information, 1983, 10(6): 1-4.
[28] Wan N F, Li S X, Li T, et al. Ecological intensification of rice production through rice-fish co-culture [J]. Journal of Cleaner Production, 2019(234): 1002-1012. doi: 10.1016/j.jclepro.2019.06.238
[29] 马微微, 陈灿, 黄璜, 等. 垄作稻鱼鸡共生对稻田土壤养分含量及水稻产量的影响 [J]. 河南农业科学, 2021, 50(8): 9-17. Ma W W, Chen C, Huang H, et al. Effect of rice-fish-chicken symbiosis on paddy soil nutrient and rice yield under ridge cultivation [J]. Journal of Henan Agricultural Sciences, 2021, 50(8): 9-17.
[30] 韩枫, 常志强, 高勇, 等. 多营养层次生态养殖模式简析 [J]. 水产养殖, 2021(4): 24-30. Han F, Chang Z Q, Gao Y, et al. Analysis of the integrated multi-trophic aquaculture model [J]. Journal of Aquaculture, 2021(4): 24-30.
[31] 张剑, 胡亮亮, 任伟征, 等. 稻鱼系统中田鱼对资源的利用及对水稻生长的影响 [J]. 应用生态学报, 2017, 28(1): 299-307. Zhang J, Hu L L, Ren W Z, et al. Effects of fish on field resource utilization and rice growth in rice-fish coculture [J]. Chinese Journal of Applied Ecology, 2017, 28(1): 299-307.
[32] 丁伟华, 李娜娜, 任伟征, 等. 传统稻鱼系统生产力提升对稻田水体环境的影响 [J]. 中国生态农业学报, 2013, 21(3): 308-314. Ding W H, Li N N, Ren W Z, et al. Effects of improved traditional rice-fish system productivity on field water environment [J]. Chinese Journal of Eco-Agriculture, 2013, 21(3): 308-314.
-
期刊类型引用(2)
1. 王裕玉,贾晶,李媛媛,张新明. 淡水鳗鲡营养需求和配合饲料研究进展. 动物营养学报. 2024(05): 2728-2742 .  百度学术
百度学术
2. 徐心雨,武之绚,鲍梦圆,王海歌,牛化欣,常杰. 胆碱的生理功能及其对鱼脂肪代谢的调控机制. 饲料研究. 2023(06): 137-141 . 百度学术
其他类型引用(2)


计量
- 文章访问数: 859
- HTML全文浏览量: 366
- PDF下载量: 57
- 被引次数: 4