
AQUATIC FOOD WEB OF THE HUN-TAI RIVER BASIN IN LIAONING BASED ON STABLE ISOTOPE ANALYSIS
-
摘要:
为探究辽宁省浑太河流域水生生物营养结构特征及其变化, 分别于2020年秋季(10月)和2021年春季(5月)对该流域开展渔业资源调查, 依据主要消费者及饵料生物样品的碳、氮稳定同位素值(δ13C和δ15N), 利用SIBER和MixSIAR模型分析渔获物群落营养结构的时空差异, 并初步构建该流域的食物网。结果表明, 主要渔获物的δ13C和δ15N值分别为–37.18‰— –19.28‰和7.98‰—16.51‰, 且季节性差异不显著(P>0.05), 但δ13C值空间差异极显著(P<0.01)。浑太河流域渔获物的营养级为1.71—4.39, 同种鱼类营养级具有极显著的时空差异(P<0.01)。与春季相比, 鱼类在秋季摄食的食物资源更丰富、所占的生态位更宽, 同时太子河的各项群落营养结构指标均优于浑河。基础食源分析结果表明水生植物与陆生植物分别为浑太河两个季度的主要碳源, 陆生植物和POM分别为浑河和太子河中鱼类的主要碳源。研究填补了对浑太河流域水生生物食物网及群落营养结构研究的空缺, 为该流域后续的保护、修复及进一步开发提供参考依据。
Abstract:In order to investigate the trophic structure characteristics and spatiotemporal variations of aquatic communities in the Hun-Tai River, an important branch of the Liaohe River, the fishery resources surveys were conducted in autumn (October) 2020 and spring (May) 2021. Based on stable isotope analysis, the δ13C and δ15N values of dominant consumers and basal food sources in the river were determined. Then the spatiotemporal variations of trophic structures and the contributions of the basal carbon sources to the aquatic food web were analyzed by SIBER and MixSIAR package. In our study, a total of 23 species of catches, belonging to 3 orders, 6 families, and 20 genera were collected. All the δ13C and δ15N values of the main catches ranged widely from –37.18‰ to –19.28‰, 7.98‰ to 16.51‰, respectively. There were no significant seasonal differences in δ13C and δ15N values (P>0.05). However, the δ13C values variations of spatiality were significant (P<0.01), and the carnivorous δ15N values were higher than omnivorous (P<0.05). The trophic level of catches, ranging from 1.71 to 4.39 in the Hun-Tai River, had extremely significant temporal and spatial differences (P<0.01). The comparison of the ecosystem trophic structure revealed that the food resources and ecological niche occupied by fish in autumn were higher and broader than that in spring, and all trophic structure indicators (NR/CR/TA/CD/NND/SDNND) of the Taizi River higher than those of the Hun River might be brought out by the higher degree of exploitation in the Hun River. In addition, the basal food sources analysis results supposed that aquatic plants and terrestrial plants were major carbon sources in autumn and spring, respectively. Meanwhile the terrestrial plants and POM were major carbon sources in the Hun River and Taizi River, respectively. This study of enriching the contents about food web in the Hun-Tai River provided references for the subsequent protection, restoration and further development, and accumulated basic data for the reconstruction of the river ecosystem in Northeast China.
-
Keywords:
- Aquatic Food Webs /
- Stable Isotopes /
- Trophic Structure /
- Hun-Tai River
-
在水域生态系统中, 种内和种间关系本质上表现为营养关系, 这种关系使水生生物构成了一个相互依存并相互制约的有机整体, 从而实现物质和能量的循环和流动[1, 2]。栖息于水域生态系统中各个物种间的营养关系并不是简单的食物链关系, 而是由各种食物链彼此交错形成的一种网状关系, 即水生食物网[1—5]。水域生态系统正是通过食物网进行着物质和能量的流动, 从而维持自身的动态平衡状态[1]。传统的食物网测定方法是肠含物分析法, 因具有工作量大、鉴别困难的缺点[6], 限制其在水生生物营养结构方面的应用, 相反稳定同位素法(Stable isotope analysis, SIA)能快速准确地评估生态系统长期的营养结构[7—10]。目前, SIA已广泛运用于池塘、河流、湖泊、海洋和湿地等水域的食物网测定。如叶学瑶等[11]通过测定阳澄湖鱼类稳定同位素值提出应当关注δ13C—δ15N双坐标图中的边缘鱼类如翘嘴鲌(Culter alburnus)、达氏鲌(Culter dabryi)的变化; 赵冬福等[12]比较增殖区与非增殖区的食物网推断放养鲢鳙会改变群落食物网结构; 李学梅等[13]研究了长湖拆围后鱼类群落营养结构的变化特征, 为该湖渔业资源修复提供依据; 霍堂斌等[14]通过对黑龙江中游主要鱼类食物网的构建得出了该生态系统营养结构图; 温周瑞等[15]研究发现浮游植物和附着藻类为贡湖流域内鱼类的主要碳源; 而国外常用δ13C、δ15N和δ18O来区分野生与养殖品种[16], 同时结合食物网分析重金属(Hg等)在食物网中的生物富集作用[17]。
浑太河流经人口稠密, 工农业发达的东北老工业基地[18—20], 近几十年来, 采砂、农田耕作、城镇化、水利工程和酷渔滥捕等人类活动已经引起流域内水生生物栖息地日渐恶化[21—23]、水生生物资源不断减少[24—26]、物种濒危程度不断加剧及生物多样性日益降低等一系列问题, 河流健康状况不佳及生态服务功能的减弱严重制约了流域内社会经济绿色高质量发展[19, 23, 24]。因此, 深入了解该生态系统的结构和功能, 颁布详细的保护政策迫在眉睫。目前, 关于浑太河流域生态系统的研究主要集中在鱼类群落结构[18—20]、水质状况[23—25]和底栖生物多样性[26, 27]等方面, 而通过SIA测定群落营养结构的相关研究还未见报道。本文针对浑太河流域面临的水生态环境恶化与渔业资源衰退等问题, 利用碳、氮稳定同位素技术分析流域内水生生物的稳定同位素值并以此建立营养级图谱, 揭示该流域水生生物营养结构, 进而构建浑太河流域食物网, 为浑太河开展生态修复和保护工作提供参考。
1. 材料与方法
1.1 研究区域概况
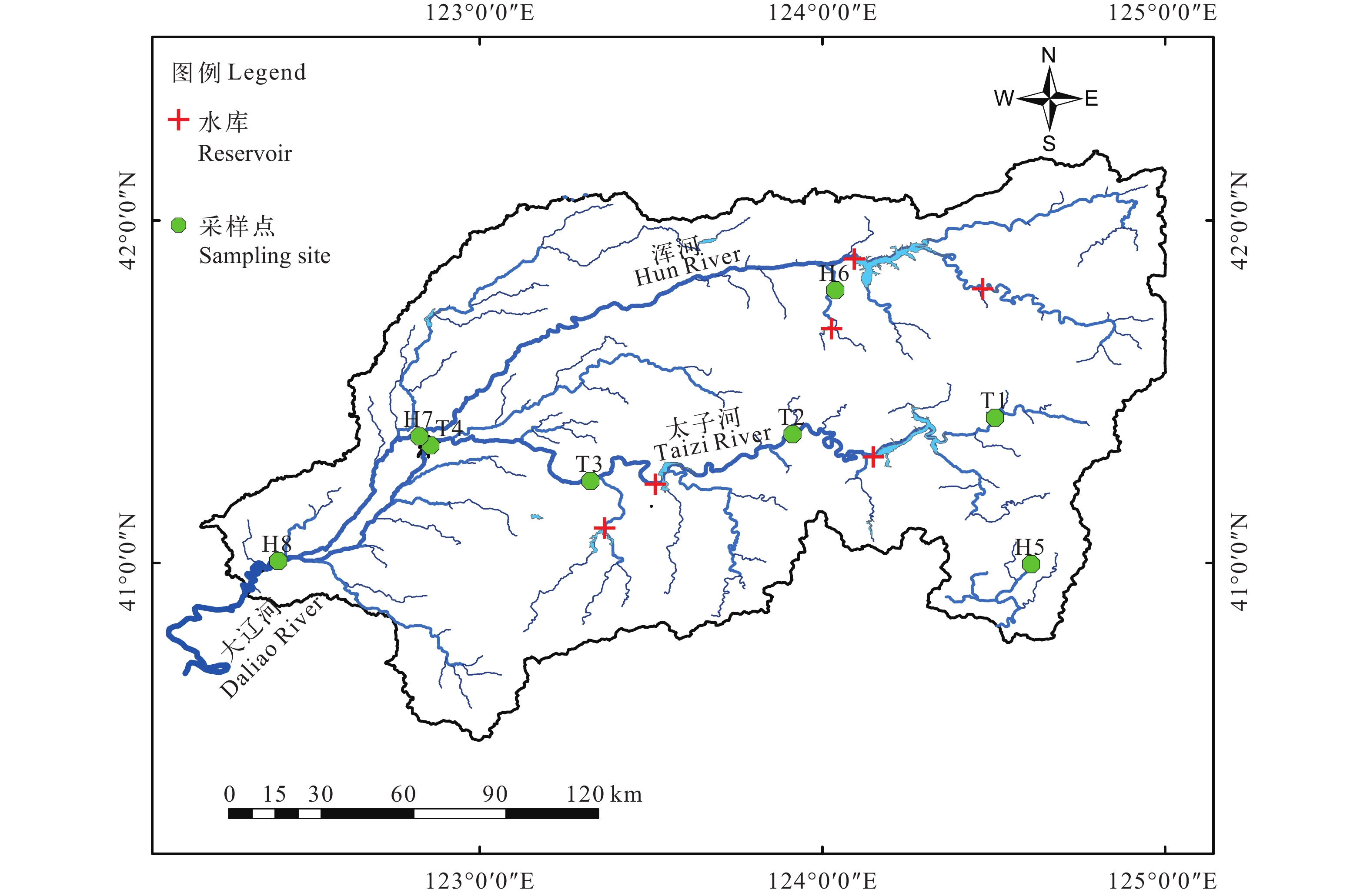
浑太河(122°05′—125°17′E, 40°40′—42°10′N)位于我国东三省之一的辽宁, 是我国辽河流域的重要分支, 主要由浑河与太子河组成, 流域面积达27300 km2, 全长近900 km[28], 流经辽宁省一些重要的农业、工业及城市区域[29]。为调查浑太河流域水生生物食物网结构, 分别于2020年秋季(10月)和2021年春季(5月)对浑太河流域的8个站位开展2次渔业资源调查工作(图 1)。
![]() 图 1 辽宁省浑太河采样站点分布图Figure 1. Distribution of sampling sites in the Hun-Tai River, Liaoning Province
图 1 辽宁省浑太河采样站点分布图Figure 1. Distribution of sampling sites in the Hun-Tai River, Liaoning Province1.2 样品采集与处理
采用刺网、电捕及地笼对鱼类样品进行采集, 现场对新鲜的样本进行种类鉴定, 并测量体长和体重, 样本测量完后转入冰柜中速冻, 运回实验室取其背部白肌。鱼类鉴别及食性判断等主要参考《东北地区淡水鱼类》[20]。植物类样本在现场采集后使用蒸馏水清洗干净后, 60℃烘干、研磨。浮游生物使用WhatmanGF/F滤膜抽滤后, 用1 mol/L盐酸酸化后, 60℃烘干、研磨。所有样本的采集和前处理方式均参照徐军等[30]。
1.3 稳定同位素分析
所有样品的同位素比值均采用稳定同位素比率质谱仪(Isoprime 100, 英国)测定, 每测定12个样品后加测1个标准样, 保证数据的准确性。碳、氮稳定同位素标准物质为咖啡因(Caffeine, IAEA-600)与大气氮, 测定结果以δ13C、δ15N表示, 其公式为:
$$ \delta{X}(\text{‰})\text=\left({{R}}_{\text{sample}}-{{R}}_{\text{Standard}}\right)/{{R}}_{\text{Standard}}\text{×1000} $$ 式中,
$\delta{X}$ 为所测样品的δ13C或δ15N,$ {{R}}_{\text{sample}} $ 为样本的同位素比值(13C/12C或15N/14N),$ {{R}}_{\text{Standard}} $ 为参考标准物质的同位素比值。营养级(Trophic level, TL)计算公式为:
$$ \text{TL} = \left({{\delta}^{\text{15}}\text{N}}_{\text{consumer}}-{{\delta}^{\text{15}}\text{N}}_{\text{baseline}}\right)/{\delta}^{\text{15}}\text{N}+\lambda $$ 式中,
${{\delta}^{\text{15}}\text{N}}_{\text{consumer}}$ 为待测样品氮同位素值,${{\delta}^{\text{15}}\text{N}}_{\text{baseline}}$ 为基准生物的氮同位素值,${\delta}^{\text{15}}\text{N}$ 为相邻营养级的氮富集度, 本研究取3.4‰[31]。λ为基准生物的营养级, 选取初级消费者浮游动物为基准(Baseline), λ=2。1.4 数据分析
使用Excel 2019进行碳、氮稳定同位素值基础统计。使用SPSS 26进行单因素方差分析(ANOVA)检验比较渔获物δ13C、δ15N值及营养结构的时空差异。通过ArcGIS 10.2、Origin 2021软件制作站点分布图及营养级图谱。分别利用基于R 4.2.1的SIBER和MixSIAR包对整个浑太河流域渔获物的δ13C和δ15N值进行分析, 计算氮同位素比值范围(δ15N range, NR)、碳同位素比值范围(δ13C range, CR)、生态位总面积(Total area, TA)、平均离心距离(Mean distance to centroid, CD)、最邻近距离平均值(Mean nearest neighbor distance, NND)和最邻近距离标准差(Standard deviation of nearest neighbor distance, SDNND)六个群落营养结构指标[32]与基础碳源对渔获物的贡献率。
2. 结果
2.1 稳定同位素δ13C、δ15N值特征
本实验两个季度共采集渔获物4276尾, 隶属于3目6科20属23种, 其中共测定同位素样品343个(表 1)。秋季主要渔获物的δ13C值为–37.18‰[黑鳍鳈(Sarcocheilichthys nigripinnis)]— –19.28‰[兴凯银(Squalidus chankaensis)], 差值达17.90‰, δ15N值为7.98‰[中华绒螯蟹(Eriocheir sinensis)]—14.15‰(黑鳍鳈), 相差6.04‰; 春季主要渔获物的δ13C值为–27.40‰[方氏鳑鲏(Rhodeus fangi)]— –20.53‰[银(Squalidus argentatus)], 相差6.87‰, δ15N值为8.72‰[抚顺(Gobio fushunensis)]—16.51‰[小黄䱂(Micropercops swinhonis)], 相差7.79‰。比较不同食性渔获物的δ13C和δ15N值发现, 肉食性[δ13C=(–24.08±3.31)‰, δ15N=(12.83±2.69)‰]的δ13C值与植食性[δ13C=(–23.89±2.84)‰, δ15N=(11.95±1.89)‰]和杂食性[δ13C=(–24.37±3.22)‰, δ15N=(11.77±2.12)‰]相比无显著差异(P>0.05), 但肉食性的δ15N值显著高于杂食性(P<0.05)。
表 1 浑太河生态系统碳、氮稳定同位素值Table 1. δ13C and δ15N values in the Hun-Tai River ecosystem (n=343, x±SD)种类Species 食性
Feeding habit秋季Autumn 春季Spring 样本量n δ13C (‰) δ15N (‰) 样本量n δ13C (‰) δ15N (‰) 䱗Hemiculter leucisculus Om 8 –24.33±0.85 12.94±0.71 10 –26.52±2.17 12.74±1.28 彩副鱊Paracheilognathus imberbis Om 5 –26.04±1.01 12.10±0.85 — — — 方氏鳑鲏Rhodeus fangi Om — — — 8 –27.40±2.59 11.76±2.52 高体鳑鲏Rhodeus lighti Om 13 –26.18±4.67 11.75±2.34 10 –25.65±2.06 12.22±1.37 大鳍鱊Acheilognathus macropterus Om 16 –28.44±6.20 12.87±3.13 13 –25.04±1.57 12.37±1.58 麦穗鱼Pseudorasbora parva Om 12 –26.67±9.36 12.67±1.28 13 –23.27±1.97 13.06±1.1 黑鳍鳈Sarcocheilichthys nigripinnis Om 3 –37.18±10.76 14.15±0.62 — — — 银Squalidus argentatus Om — — — 5 –20.53±0.54 10.87±0.47 亮银Squalidus nitens Om — — — 3 –24.52±0.29 11.48±0.37 清徐胡Huigobio chinssuensis Om 18 –22.16±4.12 11.82±1.75 7 –25.95±1.24 13.42±2.54 棒花鱼Abbottina rivularis Om — — — 7 –24.26±1.22 13.77±1.83 鲫Carassius auratus Om 36 –26.03±6.99 11.71±1.58 9 –25.11±1.82 11.06±1.84 北方须鳅Barbatula barbatula nuda Om 10 –23.95±0.53 9.00±1.02 9 –25.29±3.99 10.16±1.36 泥鳅Misgurnus anguillicaudatus Om 7 –26.85±8.77 8.11±1.95 3 –24.27±1.25 9.80±1.65 日本沼虾Macrobrachium nipponense Om 7 –22.62±1.63 12.03±1.89 6 –24.61±1.73 12.29±2.77 中华绒螯蟹Eriocheir sinensis Om 5 –25.52±0.41 7.98±2.02 — — — 宽鳍鱲Zacco platypus He 22 –23.66±2.48 12.18±1.76 19 –24.21±3.24 10.55±2.39 鲢Hypophthalmichthys molitrix He 4 –30.10±7.01 10.57±1.22 — — — 拉氏鱥Rhynchocypris lagowskii Ca 17 –22.67±3.18 10.59±3.65 18 –22.89±3.24 10.55±2.39 抚顺Gobio fushunensis Ca — — — 3 –25.95±0.73 8.72±0.74 兴凯银Squalidus chankaensis Ca 5 –19.28±0.27 11.96±0.16 — — — 小黄䱂Micropercops swinhonis Ca 6 –34.66±10.89 13.02±0.97 3 –25.55±0.24 16.51±1.49 纹缟虾虎鱼Tridentiger trigonocephalus Ca — — — 3 –26.06±0.43 14.64±0.71 浮游植物Phytoplankton 8 –26.75±1.89 4.12±0.76 8 –25.49±1.55 2.34±0.83 浮游动物Zooplankton 8 –26.38±1.80 5.19±1.71 8 –26.12±2.52 9.25±0.73 颗粒有机物Particulate organic matter 8 –26.28±1.76 4.97±2.45 8 –26.52±1.55 2.97±0.62 陆生植物Terrestrial plant 9 –30.57±4.56 7.06±2.86 4 –28.37±0.95 4.45±1.39 水生植物Aquatic plant 15 –25.55±3.63 6.90±1.61 18 –29.33±2.27 5.02±2.30 着生藻类Periphytic algae 5 –21.89±3.33 1.65±0.84 6 –24.09±3.09 5.18±1.43 注: Om、He和Ca 分别表示杂食性, 植食性和肉食性; “—”表示无相关数据Note: Om, He and Ca indicate omnivore, herbivore and carnivore, respectively. “—” indicates no data 除此之外, 选取同种类不同季节和同种类不同支流的渔获物δ13C、δ15N值进行时空差异比较。结果表明, 在整个浑太河流域, 春季时渔获物δ13C、δ15N值相较于秋季小幅度上升(图 2a; P>0.05); 而太子河渔获物的δ13C值显著高于浑河(图 2b; P<0.01), 但δ15N值无显著差异(P>0.05)。
![]() 图 2 浑太河流域同种渔获物δ13C和δ15N值的时空差异a为浑太河同种渔获物不同季节的δ13C和δ15N值; b为浑太河同种渔获物不同支流的δ13C和δ15N值; **表示差异极显著(P<0.01)Figure 2. The spatiotemporal differences of δ13C and δ15N values of the same selected species in the Hun-Tai Rivera. δ13C and δ15N values of the same selected species in different seasons in the Hun-Tai River; b. δ13C and δ15N values of the same selected species in different tributaries in the Hun-Tai River; **indicates highly significant difference (P<0.01)
图 2 浑太河流域同种渔获物δ13C和δ15N值的时空差异a为浑太河同种渔获物不同季节的δ13C和δ15N值; b为浑太河同种渔获物不同支流的δ13C和δ15N值; **表示差异极显著(P<0.01)Figure 2. The spatiotemporal differences of δ13C and δ15N values of the same selected species in the Hun-Tai Rivera. δ13C and δ15N values of the same selected species in different seasons in the Hun-Tai River; b. δ13C and δ15N values of the same selected species in different tributaries in the Hun-Tai River; **indicates highly significant difference (P<0.01)2.2 营养级
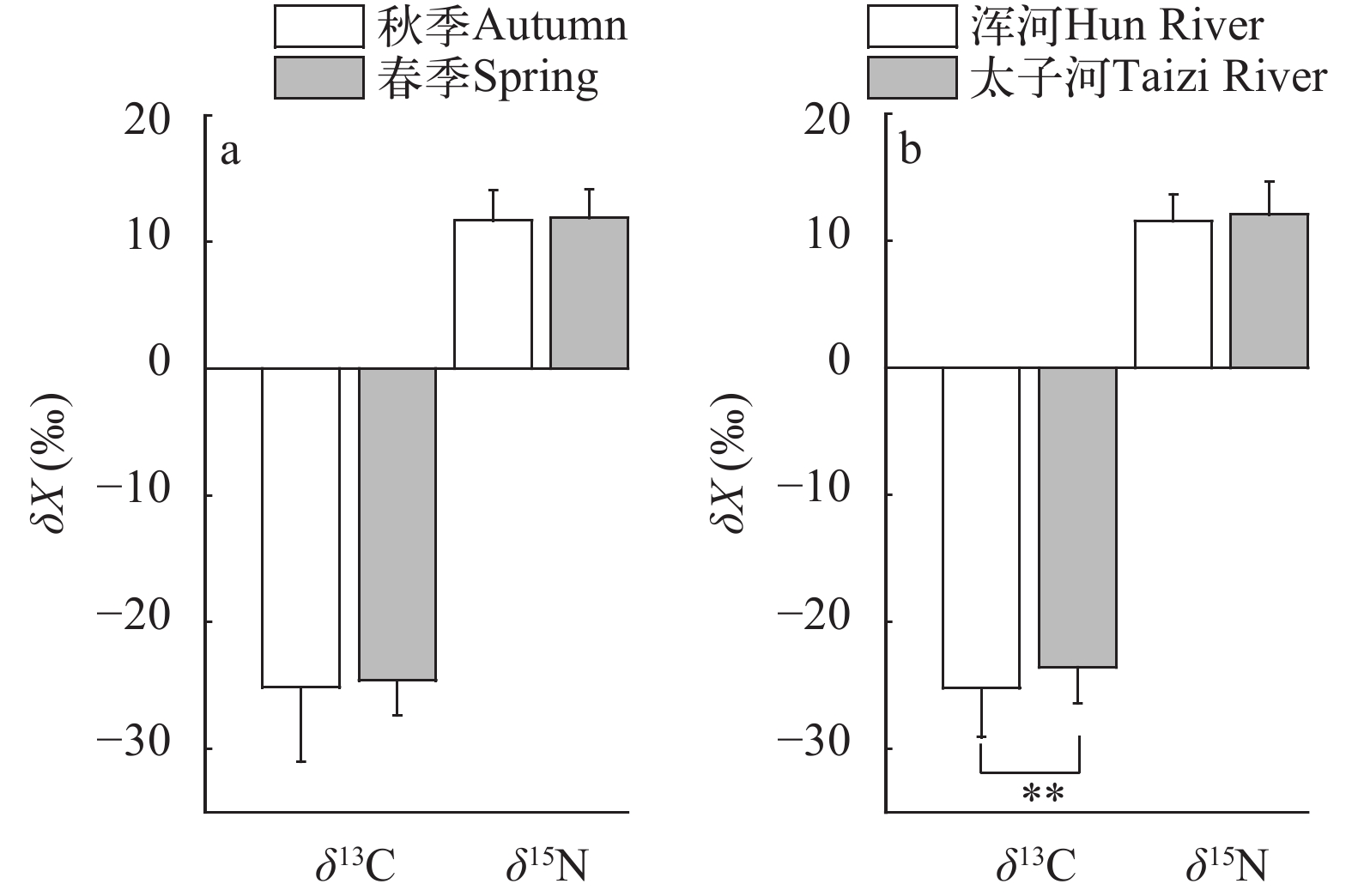
通过δ15N值计算营养级(图 3), 浑太河流域秋季主要渔获物营养级为2.57(中华绒螯蟹)—4.39(黑鳍鳈), 均值为3.64±0.69, 跨度为1.82, 属于3个营养级; 春季营养级为1.71(抚顺)—4.00(小黄䱂), 均值为2.64±0.66, 跨度为2.29, 属于4个营养级, 秋季营养级跨度小于春季。不同食性渔获物营养级之间差异较大, 杂食性的营养级为2.03—4.03, 植食性的营养级为2.49—3.81, 肉食性的营养级为1.71—4.05。
![]() 图 3 浑太河流域秋季和春季渔获物营养级图谱Figure 3. Continuous trophic spectrum of catches in autumn and spring in the Hun-Tai River
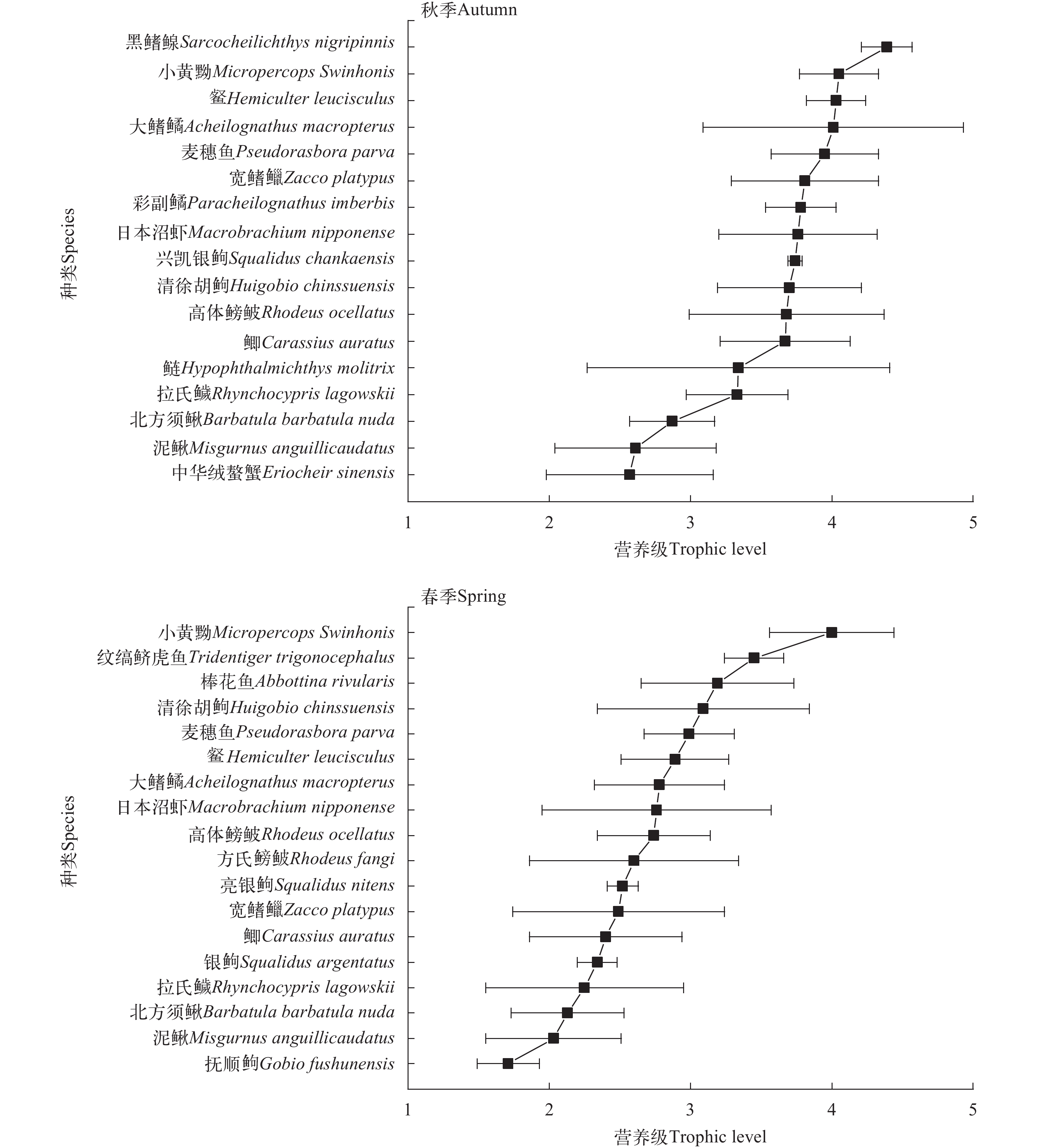
图 3 浑太河流域秋季和春季渔获物营养级图谱Figure 3. Continuous trophic spectrum of catches in autumn and spring in the Hun-Tai River选取同种类不同季节和同种类不同支流的渔获物营养级进行时空差异比较。由图 4可见, 秋季浑太河渔获物的营养级显著高于春季(秋季: 3.65±0.70; 春季: 2.64±0.66; P<0.01), 其中宽鳍鱲(Zacco platypus)、䱗(Hemiculter leucisculus)、大鳍鱊(Acheilognathus macropterus)、鲫(Carassius auratus)、拉氏鱥(Rhynchocypris lagowskii)和日本沼虾(Macrobrachium nipponense)的营养级降幅均大于1(图 3), 而降幅最小的为小黄䱂和泥鳅(Misgurnus anguillicaudatus); 不同支流的营养级差异也极显著(图 4; 浑河: 3.38±0.61; 太子河: 3.77±0.83; P<0.01), 特别是棒花鱼(Abbottina rivularis)、方氏鳑鲏、高体鳑鲏(Rhodeus lighti)、小黄䱂、麦穗鱼(Pseudorasbora parva)。
![]() 图 4 浑太河流域同种渔获物营养级的时空差异比较**表示差异极显著(P<0.01)Figure 4. The spatiotemporal differences of trophic level of the same selected species in the Hun-Tai River** indicates highly significant difference (P<0.01)
图 4 浑太河流域同种渔获物营养级的时空差异比较**表示差异极显著(P<0.01)Figure 4. The spatiotemporal differences of trophic level of the same selected species in the Hun-Tai River** indicates highly significant difference (P<0.01)2.3 群落营养结构
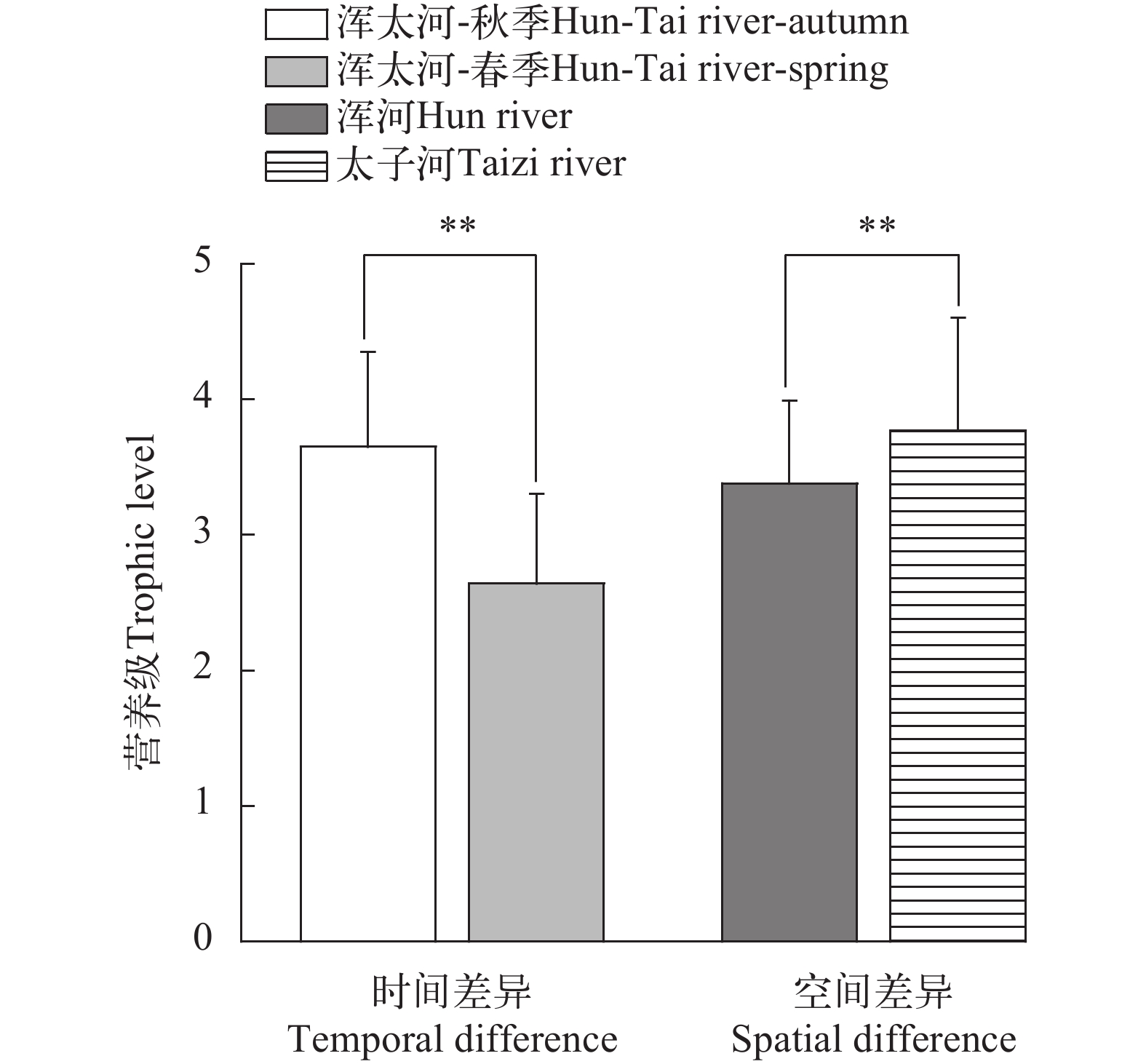
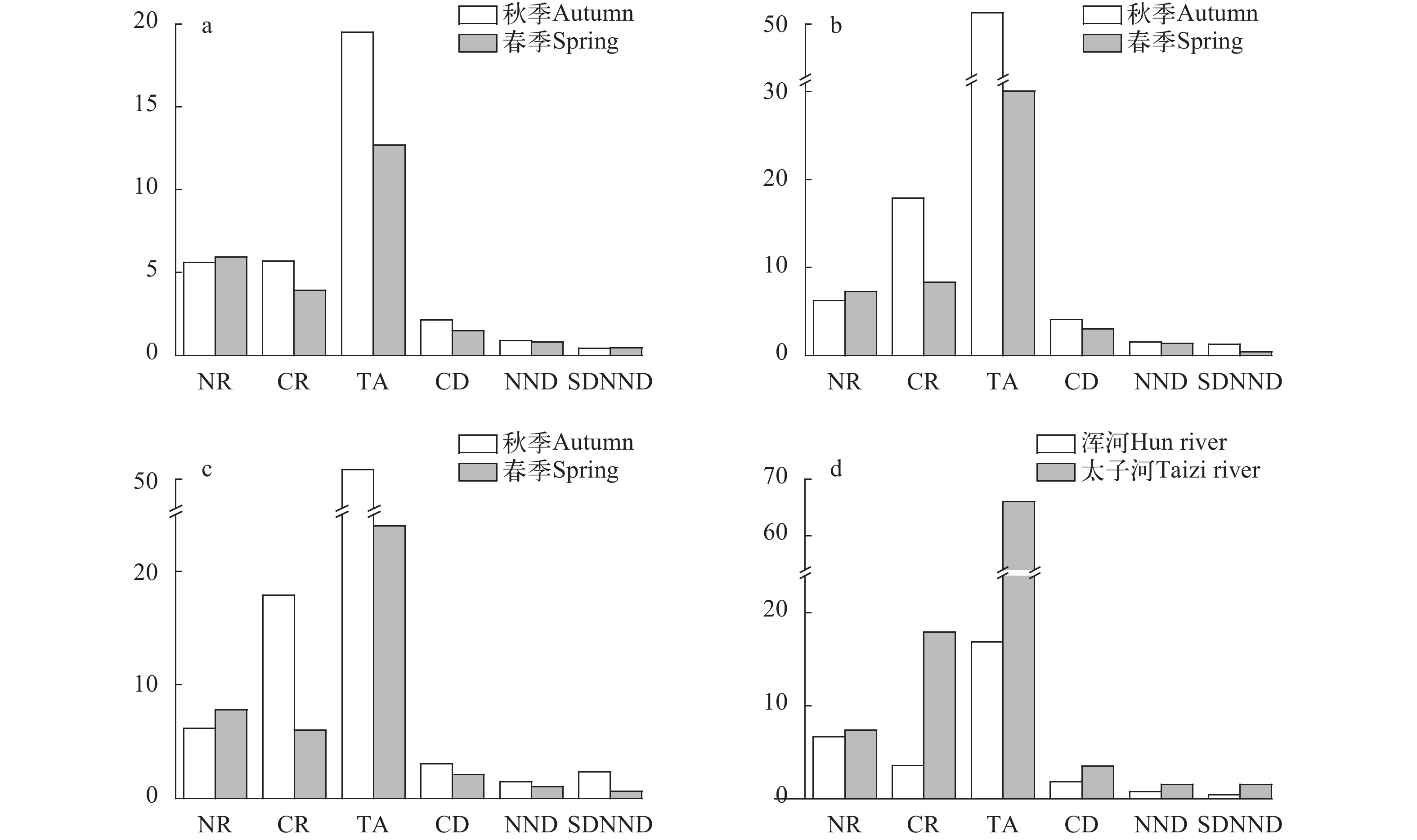
根据稳定同位素值计算生物群落指标, 在浑河流域(图 5a), 秋季的CR值(基础食物源)、TA值(生态位空间)、CD值(营养多样性)、NND值(营养位置)均高于春季, 只有NR值(食物网长度)、SDNND值(营养冗余程度)略低于春季。而在太子河流域(图 5b), 秋季的CR值、TA值、CD值、NND值、SDNND值明显高于春季, 但NR值较低。在整个浑太河流域(图 5c), CR、TA、CD、NND、SDNND值随时间呈递减趋势, 只有NR值有所上升。对比两个支流的各项指标(图 5d), 太子河的六项群落营养结构指标值均大于浑河。
![]() 图 5 浑太河流域生物群落指标a, b, c分别为浑河、太子河、浑太河不同季节鱼类生物群落指标; d为浑太河不同支流鱼类生物群落指标Figure 5. The community-wide metrics in the Hun-Tai Rivera, b and c represent the community-wide metrics in different seasons in the Hun, Taizi and Hun-Tai River respectively; d represents the community-wide metrics in different tributaries in the Hun-Tai River
图 5 浑太河流域生物群落指标a, b, c分别为浑河、太子河、浑太河不同季节鱼类生物群落指标; d为浑太河不同支流鱼类生物群落指标Figure 5. The community-wide metrics in the Hun-Tai Rivera, b and c represent the community-wide metrics in different seasons in the Hun, Taizi and Hun-Tai River respectively; d represents the community-wide metrics in different tributaries in the Hun-Tai River2.4 潜在碳源贡献率
本研究调查到的水生植物有蒌蒿(Artemisia selengensis)、水烛(Typha angustifolia)、荻(Miscanthus sacchariflorus)、芦苇(Phragmites australis)、水蓼(Polygonum hydropiper)、香蒲(Typha orientalis)、虉草(Phalaris arundinacea)、具芒碎米莎草(Cyperus microiria)和穗状狐尾藻(Myriophyllum spicatum); 陆生植物有披碱草(Elymus dahuricus)、旱柳(Salix matsudana)、春榆(Ulmus davidiana var. japonica)、汉白杨(Populus ningshanica)和酸模(Rumex acetosa)。
不同季节浑太河流域5种潜在碳源[浮游植物、陆生植物、颗粒有机物(POM)、水生植物、着生藻类]对不同鱼类食源的贡献率如表 2所示。秋季时各潜在碳源贡献率顺序为水生植物(36.15%)>陆生植物(24.24%)>着生藻类(17.10%)>浮游植物(13.21%)>POM(9.30%), 而春季时为陆生植物(27.09%)>着生藻类(22.70%)>水生植物(18.66%)>浮游植物(18.51%)>POM(13.04%), 其中水生植物两季的贡献率波动最大。在两条支流中, 浑河潜在碳源贡献率顺序为陆生植物(36.60%)>着生藻类(22.36%)>浮游植物(17.92%)>POM(12.92%)>水生植物(10.20%), 而太子河的顺序为POM(29.01%)>陆生植物(26.52%)>着生藻类(23.91%)>水生植物(10.83%)>浮游植物(9.73%), 其中陆生植物和POM的贡献率在空间分布上存在较大波动。
表 2 浑太河流域不同季度鱼类基础食物碳源贡献率[均值(5%—97.5%置信区间)]Table 2. The contribution rate of basic food carbon sources of fish in the Hun-Tai River in different seasons (mean proportion with 5% and 97.5% confidence intervals)种类
Species秋季Autumn (%) 春季Spring (%) 浮游植物
Phytoplankton陆生植物
Terrestrial plantPOM 水生植物
Aquatic plant着生藻类
Periphytic algae浮游植物
Phytoplankton陆生植物
Terrestrial plantPOM 水生植物
Aquatic plant着生藻类
Periphytic algae䱗Hemiculter leucisculus 15.4
(0.3—66.5)29.3
(5.1—72)11.9
(1—59.1)26.6
(3.1—78.4)16.7
(1.1—66.4)16.3
(0.5—66.8)41.5
(10.1—85.1)11.5
(0.4—55.3)17.1
(0.3—67.9)13.6
(0.6—52.8)彩副鱊Paracheilognathus imberbis 16.5
(0.4—64.6)24.7
(3.5—66.2)12.3
(1.3—55.7)29.9
(3.4—79.9)16.6
(0.9—71.4)方氏鳑鲏Rhodeus fangi 17
(0.5—69.2)42.6
(9.7—88.3)12.5
(0.3—57.4)18.4
(0.3—72)9.6
(0.5—37.8)高体鳑鲏Rhodeus lighti 8.4
(0.2—52.8)13.2
(0.6—57.6)5.3
(0.3—35.2)61.9
(12.4—96.7)11.1
(0.3—57.2)18
(0.4—71.5)32.2
(8.1—78.5)13.2
(0.3—59.4)18.4
(0.3—72.4)18.2
(1.1—51.8)大鳍鱊Acheilognathus macropterus 12.9
(0.4—67)41.7
(8.6—86)9
(0.5—49.7)21.7
(1.8—78)14.6
(0.8—62.6)17.7
(0.4—69.1)31.6
(7.8—70.9)14
(0.4—56.6)17.6
(0.3—67.8)19.1
(1.4—56.1)麦穗鱼Pseudorasbora parva 11.7
(0.4—58.7)25.2
(3—72.2)6.6
(0.4—39.9)40.7
(4.3—90.2)15.8
(0.7—69.9)15.6
(0.3—65.4)25.4
(6.3—63.4)10.6
(0.4—45.2)16.4
(0.3—66.7)32
(2.5—71.4)黑鳍鳈Sarcocheilichthys nigripinnis 4.9
(0.1—34.4)2.8
(0.1—14.6)2.4
(0.1—14.6)82.9
(47.2—98.9)7
(0.2—44.7)银Squalidus argentatus 19.1
(0.6—71.4)12.1
(2.1—40.7)13.1
(0.3—57.9)18.3
(0.3—72.6)37.4
(3.2—86.1)亮银Squalidus nitens 19.5
(0.8—72.1)25.5
(4.3—71.4)15.3
(0.5—67.3)19.9
(0.3—74.2)19.8
(1—61.5)清徐胡Huigobio chinssuensis 14.3
(0.5—69.7)23.2
(2.9—65.8)9.8
(0.8—57.2)33.5
(3.5—86.2)19.1
(1.1—73.3)16.3
(0.3—63.7)44.7
(11.8—85)11.8
(0.3—57.8)16.6
(0.3—65.8)10.6
(0.5—37.8)棒花鱼Abbottina rivularis 16.8
(0.4—63.3)34.9
(10.1—77.6)11.3
(0.3—48.6)16.2
(0.3—64.5)20.8
(1.1—59.7)鲫Carassius auratus 14.8
(0.5—66.1)19.7
(2.6—59.6)8.7
(0.9—45.5)39.7
(6.7—85.9)17.1
(0.9—74.7)20.6
(0.5—76)22.8
(3.9—64)15.4
(0.4—67.4)21.4
(0.3—74.1)19.8
(1.4—65.9)北方须鳅Carassius auratus 17
(1—49.9)14.1
(3.1—42.5)12.5
(3.1—35)36.2
(12.1—74.3)20.1
(2.4—64.6)18
(0.7—51.5)23.7
(11.6—55.1)12.6
(0.9—39.5)18.9
(0.7—53.1)26.8
(2.5—64.9)泥鳅Misgurnus anguillicaudatus 24.2
(0.5—77)7.5
(0.6—32.1)22.3
(1.3—70.1)23.1
(2.1—73)23
(0.9—86.2)22.4
(0.6—82.3)17.7
(2.3—60.5)16.7
(0.3—73.4)19.7
(0.3—74.6)23.5
(1.1—73)宽鳍鱲Zacco platypus 12.1
(0.4—50.2)27.4
(4.2—70.4)9.5
(0.7—52.4)33.3
(5—84.1)17.7
(1—74.2)15.6
(0.4—61.2)17.6
(4.1—50.8)10.8
(0.5—46.5)18.3
(0.3—77.2)37.6
(3.1—77.8)鲢Hypophthalmichthys molitrix 18.7
(0.5—71.5)15.9
(1.1—56.8)13.3
(1.1—54.6)35.1
(3.3—85.3)17
(0.8—74.9)拉氏鱥Rhynchocypris lagowskii 12.1
(0.4—59.4)20.5
(2.4—63.4)9.5
(0.9—53.1)37.7
(4.4—86.4)20.3
(1—75.1)20.2
(0.4—80.2)11.2
(1.9—41)11.3
(0.4—47)18.9
(0.3—73.5)38.4
(2.6—83.9)抚顺Gobio fushunensis 21.8
(0.5—80)20.6
(1.6—72.1)18.1
(0.3—84.9)22.5
(0.3—81.1)17
(0.6—71.6)兴凯银Squalidus chankaensis 9.6
(0.4—59.4)14.3
(0.8—58.1)6.7
(0.5—39)52.6
(5.5—94.9)16.7
(0.6—70)小黄䱂Micropercops swinhonis 14.3
(0.4—67.5)28.3
(2.9—79.2)9.6
(0.7—53.7)32
(2.7—89.4)15.7
(0.9—69.1)14.8
(0.4—62.1)50.8
(16.6—87.5)10.1
(0.3—47.9)14.5
(0.3—63.2)9.8
(0.5—36.3)纹缟虾虎鱼Tridentiger trigonocephalus 16.7
(0.5—66.5)44.7
(11.1—86.5)11.1
(0.3—50.9)16.5
(0.3—68.9)11.1
(0.5—40.8)3. 讨论
3.1 稳定同位素δ13C、δ15N值特征
浑太河流域(表 1)的δ13C值为–37.18‰— –19.28‰, 相差17.9‰; δ15N值为7.98‰—16.51‰, 相差8.53‰, 与黑龙江中游[14](δ13C: –32.00‰— –17.32‰; δ15N: 7.58‰—14.78‰)、长江中游[33](δ13C: –33.83‰— –17.36‰; δ15N: 4.83‰—15.13‰)、大宁河[34](δ13C: –29.74‰— –16.61‰; δ15N: 4.54‰—17.51‰)、香溪河[35](δ13C: –28.30‰— –13.90‰; δ15N: 4.80‰—14.40‰)等江河水域相似, 渔获物稳定同位素比值(δ13C和δ15N)范围较广, 远高于阳澄湖[11](δ13C: –28.98‰— –20.65‰, δ15N: 11.92‰—17.85‰)、长湖[13](δ13C: –27.70‰— –24.80‰, δ15N: 11.40‰—16.60‰)、贡湖[15](δ13C: –23.10‰— –28.10‰, δ15N: 14.50‰—19.10‰)等湖泊的稳定同位素比值范围, 这可能由于江河相比于湖泊流域面积大, 水体流动性强, 基础外源性食物源输入较多(δ13C值范围大)[14, 33], 其鱼类营养级位置较为分散(δ15N值范围大), 食物竞争压力较小。另外稳定同位素比值与海拔和经纬度相关[36], 浑太河流域海拔差在1500 m左右[28], 远高于上述湖泊。
生物组织的δ13C值可以确认食物来源, 而δ15N值可以计算营养级[37]。本研究肉食性、植食性和杂食性鱼类三者的δ13C值无显著差异(表 1; P>0.05), 据此推测三者的食物来源较接近。而植食性鱼类的δ13C值略高于其余两种鱼类, 与着生藻类的δ13C值较高有一定联系[11]。肉食性鱼类的δ15N值最高, 也符合其处在群落中较高的营养位置。植食性和杂食性鱼类δ15N值无差异(P>0.05)可能由于本流域中水生植物占了杂食性鱼类基础食源的很大部分[11]。不同季节浑太河流域同种类渔获物的δ13C值与δ15N值变化不显著(图 2a; P>0.05), 但随时间有所上升, 可能由于浑太河春季日照时间长, 风力大等气候条件[28], 导致渔获物减少低δ13C、δ15N值水生植物的摄食比例, 增加高δ13C、δ15N值着生藻类的摄食比例。浑河同种类渔获物的δ13C、δ15N值均小于太子河(图 2b), 且δ13C值差异极显著(P<0.01), 除浑河流域面积较小外[28], 还可能由于浑河流域内农业用地较多[38], 而太子河流域周围以林地为主[39], 外源性碳源较为丰富。
3.2 营养级
浑太河秋季主要渔获物营养级范围跨度小于春季(图 3), 主要原因可能为秋季没有采集到抚顺等低营养级消费者, 春季没有黑鳍鳈等高营养级消费者所导致的物种组成差异所致。浑太河流域的营养级为1.71—4.39(图 3), 黑龙江中游[14]的营养级为1.06—3.98, 跨度均超过了2.5个营养级, 而太湖[15](3.30—4.40)与阳澄湖[11](2.05—3.80)营养级范围较小。同时比较浑太河与其他水域相同鱼类的营养级发现, 浑太河鱼类营养级偏高[9, 11, 13, 15], 推测这些现象是由于δ15N基准物的选择不同[40], 本研究选取的基准物为浮游动物, 而其余湖泊选取了底栖无脊椎动物蚌类和螺类[9, 11, 13, 15], 二者的δ15N值均存在一定的种内、种间差异[40]。为了减少由种类组成造成浮游动物δ15N值差异, 本研究使用浮游生物网在采样站点100 m范围内进行3—5次收集并混合, 并将非浮游动物物质分离。同时大型河流与湖泊的营养程度不同, 而栖息环境的营养状态会影响食物组成及营养级[40, 41], 且本研究采集的渔获物体型偏小, 不同规格鱼类具有显著的稳定同位素差异[7, 41], 这些因素都可能造成营养级差异。河流生态系统中水生生物营养级的高低也与其渔业资源开发情况有关[11], 本研究中低营养级渔获物占比偏多(图 3), 由此可知浑太河流域水生生物被过度开发。
秋季的营养级远高于春季(图 4; P<0.01), 推测与春季浮游植物生物量高(未发表数据)有关, 消费者以初级生产者浮游植物为主要饵料导致营养级降低[11]。同时营养级范围波动大表明其食物来源广, 可能与本研究渔获物以杂食性为主有关。小黄䱂和泥鳅的营养级随季节波动较小, 可能由于二者均是栖息在底层水体且是以动物食性为主的杂食性鱼类[20], 而浑太河流域底栖动物群落相对稳定[29]。两条支流的营养级差异也极显著(图 4; P<0.01), 造成这一现象的原因除了食物组成不同外, 基础食物源的稳定同位素值也存在空间差异。
3.3 群落营养结构分析
鱼类群落营养结构是评估生态系统健康状况的一个重要方法。浑太河秋季的NR值较春季的小(图 5c), CR值大, 表明其食物网长度较小, 但其基础食物源更丰富, 可能是由于: (1)两季渔获物种类组成差异; (2)浮游生物量较低时, 部分鱼类必须广泛摄食以获取生长所需能量[11, 15]; (3)陆生植物的δ13C值较高, 通过同位素分馏效应升高了以此为食的渔获物的δ13C值, 如黑鳍鳈; (4)浑太河流域广, 人为因素, 如城市生活用水、工业废水等流入生态系统中[19, 21, 22], 从而增加了食物多样性(CR值)。秋季的TA值(δ13C—δ15N面积)大于春季(图 5c), 说明秋季鱼类的生态位总空间大于春季。秋季的鱼类多样性程度较高(CD值), 鱼类营养位置差异大(NND值), 营养生态位分布不均匀(SDNND值)。浑河(图 5a)、太子河(图 5b)两季度各项指标的整体趋势与浑太河流域(图 5c)大致相同, 只有浑河的SDNND值趋势与之相反, 表明秋季浑河营养冗余度较高, 因为秋季时水生食物网中消费者的营养级较为集中。对比两季度浑河与太子河的各项生物群落指标(图 5d), 太子河的多项指标都优于浑河, 表现在其食物多样性水平高、营养层级高、生态位空间大、群落营养冗余程度低, 这一点也与太子河F-IBI、IEI等评价优于浑河相互印证[24]。
本研究结果显示浑太河流域鱼类群落营养结构具有一定的时空特征。春季基础食物源较少, 可以通过人工种植水生植物, 培养绿藻等浮游植物补充基础饵料生物[37]; 秋季鱼类多样性较低, 可以以增殖放流土著鱼类、人工建设产卵场等来改善[19]; 相较于太子河流域, 浑河流域更加需要开展一系列保护措施来恢复其生态系统的健康。
3.4 潜在碳源贡献率分析
潜在碳源是维持河流生态系统稳定的基石[42, 43], 根据其来源不同分为内源性和外源性碳源[37]。浑太河流域广, 既有水生植物、着生藻类和浮游植物等内源性碳源, 也有部分比例的陆生植物等外源性碳源, 且种类与张远等[44]对浑太河流域的调查结果基本一致。
河流周围生境不同, 食物源组成也不同[8, 42, 43]。在浑河流域, 陆生植物贡献率最大; 而太子河的主要食源为POM, 导致这一现象的原因可能除了鱼类种类组成不同和两条支流周围土地利用率不同有关以外[8], 浑河还有着较明显的季节性水量变化[45], 水量增加至淹没河岸导致陆生植物输入水体。这种基础碳源的空间差异在珠江等流域都存在[42, 43]。而水生植物与陆生植物贡献率的季节性变化可能与其生物量有关[42, 43], 辽宁省内河岸周围的陆生植物自8月开始衰退[46], 而春季(5月)恰是陆生植物的快速生长阶段, 而此时浑太河的水温等环境因子不适宜水生植物的生长[44], 导致秋季水生植物为主要食物源, 春季陆生植物为主要食物源。由于浑太河流经重工业区等造成了一定程度的重金属污染[26], 本研究也通过潜在碳源贡献率分析为该流域未来开展重金属沿食物链的生物富集效应及其治理等研究提供了基础数据。
4. 结论
本研究通过SIA初步构建了浑太河生态系统食物网, 分析了其流域内主要渔获物的营养结构, 应用SIBER模型分析该流域营养结构的时空差异, 同时使用MixSIAR模型揭示潜在碳源对水生生物的贡献率, 为东北地区河流生态系统食物网的构建及未来河流的管理、规划等提供理论依据。但由于在采样期间, 上游多数水库因泄洪及农业灌溉等需求放水导致水位上涨, 严重影响底栖生物的采集, 因此本研究未涉及底栖生物, 并使用浮游动物为基准; 部分浑太河特有和名贵珍稀鱼类, 如雷氏七鳃鳗(Lampetra reissneri)等并未采集到, 且部分种类仅出现1次, 比较群体营养结构的时空差异时无法排除鱼类物种差异。在以后的研究中, 应持续检测该流域内水生生物的营养结构, 以此来分析该地区人为活动对河流生态系统的影响, 为改善河流生态环境提出建议。
-
![]()
图 1 辽宁省浑太河采样站点分布图
Figure 1. Distribution of sampling sites in the Hun-Tai River, Liaoning Province
![]()
图 2 浑太河流域同种渔获物δ13C和δ15N值的时空差异
a为浑太河同种渔获物不同季节的δ13C和δ15N值; b为浑太河同种渔获物不同支流的δ13C和δ15N值; **表示差异极显著(P<0.01)
Figure 2. The spatiotemporal differences of δ13C and δ15N values of the same selected species in the Hun-Tai River
a. δ13C and δ15N values of the same selected species in different seasons in the Hun-Tai River; b. δ13C and δ15N values of the same selected species in different tributaries in the Hun-Tai River; **indicates highly significant difference (P<0.01)
![]()
图 3 浑太河流域秋季和春季渔获物营养级图谱
Figure 3. Continuous trophic spectrum of catches in autumn and spring in the Hun-Tai River
![]()
图 4 浑太河流域同种渔获物营养级的时空差异比较
**表示差异极显著(P<0.01)
Figure 4. The spatiotemporal differences of trophic level of the same selected species in the Hun-Tai River
** indicates highly significant difference (P<0.01)
![]()
图 5 浑太河流域生物群落指标
a, b, c分别为浑河、太子河、浑太河不同季节鱼类生物群落指标; d为浑太河不同支流鱼类生物群落指标
Figure 5. The community-wide metrics in the Hun-Tai River
a, b and c represent the community-wide metrics in different seasons in the Hun, Taizi and Hun-Tai River respectively; d represents the community-wide metrics in different tributaries in the Hun-Tai River
表 1 浑太河生态系统碳、氮稳定同位素值
Table 1 δ13C and δ15N values in the Hun-Tai River ecosystem (n=343, x±SD)
种类Species 食性
Feeding habit秋季Autumn 春季Spring 样本量n δ13C (‰) δ15N (‰) 样本量n δ13C (‰) δ15N (‰) 䱗Hemiculter leucisculus Om 8 –24.33±0.85 12.94±0.71 10 –26.52±2.17 12.74±1.28 彩副鱊Paracheilognathus imberbis Om 5 –26.04±1.01 12.10±0.85 — — — 方氏鳑鲏Rhodeus fangi Om — — — 8 –27.40±2.59 11.76±2.52 高体鳑鲏Rhodeus lighti Om 13 –26.18±4.67 11.75±2.34 10 –25.65±2.06 12.22±1.37 大鳍鱊Acheilognathus macropterus Om 16 –28.44±6.20 12.87±3.13 13 –25.04±1.57 12.37±1.58 麦穗鱼Pseudorasbora parva Om 12 –26.67±9.36 12.67±1.28 13 –23.27±1.97 13.06±1.1 黑鳍鳈Sarcocheilichthys nigripinnis Om 3 –37.18±10.76 14.15±0.62 — — — 银Squalidus argentatus Om — — — 5 –20.53±0.54 10.87±0.47 亮银Squalidus nitens Om — — — 3 –24.52±0.29 11.48±0.37 清徐胡Huigobio chinssuensis Om 18 –22.16±4.12 11.82±1.75 7 –25.95±1.24 13.42±2.54 棒花鱼Abbottina rivularis Om — — — 7 –24.26±1.22 13.77±1.83 鲫Carassius auratus Om 36 –26.03±6.99 11.71±1.58 9 –25.11±1.82 11.06±1.84 北方须鳅Barbatula barbatula nuda Om 10 –23.95±0.53 9.00±1.02 9 –25.29±3.99 10.16±1.36 泥鳅Misgurnus anguillicaudatus Om 7 –26.85±8.77 8.11±1.95 3 –24.27±1.25 9.80±1.65 日本沼虾Macrobrachium nipponense Om 7 –22.62±1.63 12.03±1.89 6 –24.61±1.73 12.29±2.77 中华绒螯蟹Eriocheir sinensis Om 5 –25.52±0.41 7.98±2.02 — — — 宽鳍鱲Zacco platypus He 22 –23.66±2.48 12.18±1.76 19 –24.21±3.24 10.55±2.39 鲢Hypophthalmichthys molitrix He 4 –30.10±7.01 10.57±1.22 — — — 拉氏鱥Rhynchocypris lagowskii Ca 17 –22.67±3.18 10.59±3.65 18 –22.89±3.24 10.55±2.39 抚顺Gobio fushunensis Ca — — — 3 –25.95±0.73 8.72±0.74 兴凯银Squalidus chankaensis Ca 5 –19.28±0.27 11.96±0.16 — — — 小黄䱂Micropercops swinhonis Ca 6 –34.66±10.89 13.02±0.97 3 –25.55±0.24 16.51±1.49 纹缟虾虎鱼Tridentiger trigonocephalus Ca — — — 3 –26.06±0.43 14.64±0.71 浮游植物Phytoplankton 8 –26.75±1.89 4.12±0.76 8 –25.49±1.55 2.34±0.83 浮游动物Zooplankton 8 –26.38±1.80 5.19±1.71 8 –26.12±2.52 9.25±0.73 颗粒有机物Particulate organic matter 8 –26.28±1.76 4.97±2.45 8 –26.52±1.55 2.97±0.62 陆生植物Terrestrial plant 9 –30.57±4.56 7.06±2.86 4 –28.37±0.95 4.45±1.39 水生植物Aquatic plant 15 –25.55±3.63 6.90±1.61 18 –29.33±2.27 5.02±2.30 着生藻类Periphytic algae 5 –21.89±3.33 1.65±0.84 6 –24.09±3.09 5.18±1.43 注: Om、He和Ca 分别表示杂食性, 植食性和肉食性; “—”表示无相关数据Note: Om, He and Ca indicate omnivore, herbivore and carnivore, respectively. “—” indicates no data  下载: 导出CSV
下载: 导出CSV
表 2 浑太河流域不同季度鱼类基础食物碳源贡献率[均值(5%—97.5%置信区间)]
Table 2 The contribution rate of basic food carbon sources of fish in the Hun-Tai River in different seasons (mean proportion with 5% and 97.5% confidence intervals)
种类
Species秋季Autumn (%) 春季Spring (%) 浮游植物
Phytoplankton陆生植物
Terrestrial plantPOM 水生植物
Aquatic plant着生藻类
Periphytic algae浮游植物
Phytoplankton陆生植物
Terrestrial plantPOM 水生植物
Aquatic plant着生藻类
Periphytic algae䱗Hemiculter leucisculus 15.4
(0.3—66.5)29.3
(5.1—72)11.9
(1—59.1)26.6
(3.1—78.4)16.7
(1.1—66.4)16.3
(0.5—66.8)41.5
(10.1—85.1)11.5
(0.4—55.3)17.1
(0.3—67.9)13.6
(0.6—52.8)彩副鱊Paracheilognathus imberbis 16.5
(0.4—64.6)24.7
(3.5—66.2)12.3
(1.3—55.7)29.9
(3.4—79.9)16.6
(0.9—71.4)方氏鳑鲏Rhodeus fangi 17
(0.5—69.2)42.6
(9.7—88.3)12.5
(0.3—57.4)18.4
(0.3—72)9.6
(0.5—37.8)高体鳑鲏Rhodeus lighti 8.4
(0.2—52.8)13.2
(0.6—57.6)5.3
(0.3—35.2)61.9
(12.4—96.7)11.1
(0.3—57.2)18
(0.4—71.5)32.2
(8.1—78.5)13.2
(0.3—59.4)18.4
(0.3—72.4)18.2
(1.1—51.8)大鳍鱊Acheilognathus macropterus 12.9
(0.4—67)41.7
(8.6—86)9
(0.5—49.7)21.7
(1.8—78)14.6
(0.8—62.6)17.7
(0.4—69.1)31.6
(7.8—70.9)14
(0.4—56.6)17.6
(0.3—67.8)19.1
(1.4—56.1)麦穗鱼Pseudorasbora parva 11.7
(0.4—58.7)25.2
(3—72.2)6.6
(0.4—39.9)40.7
(4.3—90.2)15.8
(0.7—69.9)15.6
(0.3—65.4)25.4
(6.3—63.4)10.6
(0.4—45.2)16.4
(0.3—66.7)32
(2.5—71.4)黑鳍鳈Sarcocheilichthys nigripinnis 4.9
(0.1—34.4)2.8
(0.1—14.6)2.4
(0.1—14.6)82.9
(47.2—98.9)7
(0.2—44.7)银Squalidus argentatus 19.1
(0.6—71.4)12.1
(2.1—40.7)13.1
(0.3—57.9)18.3
(0.3—72.6)37.4
(3.2—86.1)亮银Squalidus nitens 19.5
(0.8—72.1)25.5
(4.3—71.4)15.3
(0.5—67.3)19.9
(0.3—74.2)19.8
(1—61.5)清徐胡Huigobio chinssuensis 14.3
(0.5—69.7)23.2
(2.9—65.8)9.8
(0.8—57.2)33.5
(3.5—86.2)19.1
(1.1—73.3)16.3
(0.3—63.7)44.7
(11.8—85)11.8
(0.3—57.8)16.6
(0.3—65.8)10.6
(0.5—37.8)棒花鱼Abbottina rivularis 16.8
(0.4—63.3)34.9
(10.1—77.6)11.3
(0.3—48.6)16.2
(0.3—64.5)20.8
(1.1—59.7)鲫Carassius auratus 14.8
(0.5—66.1)19.7
(2.6—59.6)8.7
(0.9—45.5)39.7
(6.7—85.9)17.1
(0.9—74.7)20.6
(0.5—76)22.8
(3.9—64)15.4
(0.4—67.4)21.4
(0.3—74.1)19.8
(1.4—65.9)北方须鳅Carassius auratus 17
(1—49.9)14.1
(3.1—42.5)12.5
(3.1—35)36.2
(12.1—74.3)20.1
(2.4—64.6)18
(0.7—51.5)23.7
(11.6—55.1)12.6
(0.9—39.5)18.9
(0.7—53.1)26.8
(2.5—64.9)泥鳅Misgurnus anguillicaudatus 24.2
(0.5—77)7.5
(0.6—32.1)22.3
(1.3—70.1)23.1
(2.1—73)23
(0.9—86.2)22.4
(0.6—82.3)17.7
(2.3—60.5)16.7
(0.3—73.4)19.7
(0.3—74.6)23.5
(1.1—73)宽鳍鱲Zacco platypus 12.1
(0.4—50.2)27.4
(4.2—70.4)9.5
(0.7—52.4)33.3
(5—84.1)17.7
(1—74.2)15.6
(0.4—61.2)17.6
(4.1—50.8)10.8
(0.5—46.5)18.3
(0.3—77.2)37.6
(3.1—77.8)鲢Hypophthalmichthys molitrix 18.7
(0.5—71.5)15.9
(1.1—56.8)13.3
(1.1—54.6)35.1
(3.3—85.3)17
(0.8—74.9)拉氏鱥Rhynchocypris lagowskii 12.1
(0.4—59.4)20.5
(2.4—63.4)9.5
(0.9—53.1)37.7
(4.4—86.4)20.3
(1—75.1)20.2
(0.4—80.2)11.2
(1.9—41)11.3
(0.4—47)18.9
(0.3—73.5)38.4
(2.6—83.9)抚顺Gobio fushunensis 21.8
(0.5—80)20.6
(1.6—72.1)18.1
(0.3—84.9)22.5
(0.3—81.1)17
(0.6—71.6)兴凯银Squalidus chankaensis 9.6
(0.4—59.4)14.3
(0.8—58.1)6.7
(0.5—39)52.6
(5.5—94.9)16.7
(0.6—70)小黄䱂Micropercops swinhonis 14.3
(0.4—67.5)28.3
(2.9—79.2)9.6
(0.7—53.7)32
(2.7—89.4)15.7
(0.9—69.1)14.8
(0.4—62.1)50.8
(16.6—87.5)10.1
(0.3—47.9)14.5
(0.3—63.2)9.8
(0.5—36.3)纹缟虾虎鱼Tridentiger trigonocephalus 16.7
(0.5—66.5)44.7
(11.1—86.5)11.1
(0.3—50.9)16.5
(0.3—68.9)11.1
(0.5—40.8)
下载: 导出CSV
-
[1] 殷名称. 鱼类生态学 [M]. 北京: 中国农业出版社, 1995: 231-236. Yin M C. Fish Ecology [M]. Beijing: China Agriculture Press, 1995: 231-236.
[2] Lindeman R L. The trophic-dynamic aspect of ecology [J]. Ecology, 1942, 23(4): 399-417. doi: 10.2307/1930126
[3] Pimm S L. Food Webs [M]. London: Chapman and Hall, 1982: 1-11.
[4] Belgrano A, Scharler U M, Dunne J, et al. Aquatic Food Webs: An Ecosystem Approach [M]. Oxford: Oxford University Press, 2004: 1-66.
[5] Kwak I S, Park Y S. Food chains and food webs in aquatic ecosystems [J]. Applied Sciences, 2020, 10(14): 5012. doi: 10.3390/app10145012
[6] 高小迪, 陈新军, 李云凯. 水生食物网研究方法的发展和应用 [J]. 中国水产科学, 2018, 25(6): 1347-1360. doi: 10.3724/SP.J.1118.2018.17334 Gao X D, Chen X J, Li Y K. A review on the methods used in aquatic food web research: development and applications [J]. Journal of Fishery Sciences of China, 2018, 25(6): 1347-1360. doi: 10.3724/SP.J.1118.2018.17334
[7] 杨蕊, 韩东燕, 高春霞, 等. 浙江南部近海前肛鳗营养生态位变化研究——基于稳定同位素技术 [J]. 生态学报, 2022, 42(23): 9796-9807. Yang R, Han D Y, Gao C X, et al. Chang of trophic niche of Dysomma anguillare in the offshore waters of southern Zhejiang by stable isotope analysis [J]. Acta Ecologica Sinica, 2022, 42(23): 9796-9807.
[8] 彭彪彪, 王思凯, 赵峰, 等. 长江口盐沼湿地3种亚生境中国花鲈的碳、氮稳定同位素特征及基础食源差异 [J]. 中国水产科学, 2022, 29(2): 295-303. Peng B B, Wang S K, Zhao F, et al. Carbon and nitrogen isotopic characteristics and food source differences of Lateolabrax maculatus in three sub habitats of salt marsh wetland in the Yangtze River Estuary [J]. Journal of Fishery Sciences of China, 2022, 29(2): 295-303.
[9] 郭雯, 黄林培, 王明果, 等. 不同组织碳、氮元素含量和同位素分馏特征研究——以抚仙湖草鱼、鱇浪白鱼为例 [J]. 中国环境科学, 2022, 42(1): 345-355. doi: 10.19674/j.cnki.issn1000-6923.2022.0013 Guo W, Huang L P, Wang M G, et al. Carbon and nitrogen contents and isotopic fractionation in different tissues of Ctenopharyngodon idellus and Anabarilius grahami in Fuxian Lake [J]. China Environmental Science, 2022, 42(1): 345-355. doi: 10.19674/j.cnki.issn1000-6923.2022.0013
[10] 王晨. 水文变化对巴松错食物源及食物网特征的影响 [D]. 武汉: 华中农业大学, 2020: 1-14. Wang C. Effects of hydrological variation on food organisms and food webs in lake Basomtso [D]. Wuhan: Huazhong Agriculture University, 2020: 1-14.
[11] 叶学瑶, 任泷, 匡箴, 等. 基于稳定同位素技术的阳澄湖鱼类群落营养结构研究 [J]. 中国水产科学, 2021, 28(6): 703-714. Ye X Y, Ren L, Kuang Z, et al. Analysis of the trophic structure of fish populations in Yangcheng Lake based on stable isotope technology [J]. Journal of Fishery Sciences of China, 2021, 28(6): 703-714.
[12] 赵冬福, 任泷, 任可成, 等. 梅梁湾鲢鳙生态增殖区内外食物网结构差异 [J]. 中国水产科学, 2022, 29(11): 1601-1611. Zhao D F, Ren L, Ren K C, et al. Differences in the structure of food webs inside and outside the ecological stocking area of silver carp and bighead carp in the Meiliang Bay [J]. Journal of Fishery Sciences of China, 2022, 29(11): 1601-1611.
[13] 李学梅, 朱挺兵, 王旭歌, 等. 基于稳定同位素技术的长湖鱼类营养结构研究 [J]. 生物资源, 2021, 43(6): 545-551. Li X M, Zhu T B, Wang X G, et al. Study on trophic structure of fish communities in Changhu Lake based on stable carbon and nitrogen isotopes [J]. Biotic Resources, 2021, 43(6): 545-551.
[14] 霍堂斌, 姜作发, 马波, 等. 应用同位素分析黑龙江中游主要鱼类营养层次 [J]. 淡水渔业, 2013, 43(2): 3-8. Huo T B, Jiang Z F, Ma B, et al. Study on trophic level of main fishes in the middle reach of Heilongjiang River by stable carbon and nitrogen isotopes analysis [J]. Freshwater Fisheries, 2013, 43(2): 3-8.
[15] 温周瑞, 熊鹰, 徐军, 等. 太湖贡湖湾食物网特征研究 [J]. 水生生物学报, 2016, 40(1): 131-138. Wen Z R, Xiong Y, Xu J, et al. The studies on the structures of the food web and the trophic relationships in the Gonghu Bay of the Taihu Lake [J]. Acta Hydrobiologica Sinica, 2016, 40(1): 131-138.
[16] Arechavala-Lopez P, Fernandez-Jover D, Black K D, et al. Differentiating the wild or farmed origin of Mediterranean fish: a review of tools for sea bream and sea bass [J]. Reviews in Aquaculture, 2013, 5(3): 137-157. doi: 10.1111/raq.12006
[17] Hilgendag I R, Swanson H K, Lewis C W, et al. Mercury biomagnification in benthic, pelagic, and benthopelagic food webs in an Arctic marine ecosystem [J]. Science of the Total Environment, 2022(841): 156424.
[18] Wang X N, Ding H Y, He X G, et al. Assessing fish species tolerance in the Huntai River Basin, China: biological traits versus weighted averaging approaches [J]. Water, 2018, 10(12): 1843. doi: 10.3390/w10121843
[19] 刘猛, 渠晓东, 彭文启, 等. 浑太河流域鱼类生物完整性指数构建与应用 [J]. 环境科学研究, 2016, 29(3): 343-352. Liu M, Qu X D, Peng W Q, et al. Development and application of a fish-based index of biological integrity for the Hun-Tai River Basin [J]. Research of Environmental Sciences, 2016, 29(3): 343-352.
[20] 解玉浩. 东北地区淡水鱼类 [M]. 沈阳: 辽宁科学技术出版社, 2007: 1-483. Xie Y H. Freshwater Fishes in Northeast Region of China [M]. Shenyang: Liaoning Science and Technology Publishing House, 2007: 1-483.
[21] 魏锦宏, 谭春阳, 宋开山, 等. 2000—2010年浑太河流域土地利用与景观格局变化研究 [J]. 林业资源管理, 2014(1): 98-103. Wei J H, Tan C Y, Song K S, et al. Study on land use and landscape shifting in Huntai River Basin from 2000 to 2010 [J]. Forest Resources Management, 2014(1): 98-103.
[22] 黄荣. 辽河流域河流生态环境特征及其分类研究 [D]. 沈阳: 辽宁大学, 2015: 4-64. Huang R, Research on the river ecology environment and river classification in Liaohe River Basin [D]. Shenyang: Liaoning University, 2015: 4-64.
[23] 李尧, 刘建卫, 秦国帅, 等. 浑太流域水质演变特征及污染源解析 [J]. 中国农村水利水电, 2021(8): 14-17,22. Li Y, Liu J W, Qin G S, et al. Water quality evolution characteristics and pollution source analysis in Huntai River Basin [J]. China Rural Water and Hydropower, 2021(8): 14-17,22.
[24] 张杰, 苏航, 盛楚涵, 等. 浑太河河流生态系统完整性评价体系的构建 [J]. 环境科学研究, 2020, 33(2): 363-374. Zhang J, Su H, Sheng C H, et al. Construction of an evaluation system to assess the ecosystem integrity of the Hun-Tai River [J]. Research of Environmental Sciences, 2020, 33(2): 363-374.
[25] 李柳阳, 邵田田, 张鑫, 等. 伊洛河和浑太河春季水体光学吸收特征的对比 [J]. 环境科学, 2018, 39(9): 4122-4131. Li L Y, Shao T T, Zhang X, et al. Comparative characteristics of optical absorption in waters from Yiluo River and Huntai River in spring [J]. Environmental Science, 2018, 39(9): 4122-4131.
[26] Qu X D, Ren Z, Zhang M, et al. Sediment heavy metals and benthic diversities in Hun-Tai River, northeast of China [J]. Environmental Science and Pollution Research, 2017, 24(11): 10662-10673. doi: 10.1007/s11356-017-8642-0
[27] 张晏溧. 浑太河沉积物中重金属分布特征及其对大型底栖动物的影响 [D]. 重庆: 西南大学, 2012: 1-52. Zhang Y L. Distribution of heavy metals in sediments of Hun-Tai River and the impact on macroinvertebrate [D]. Chongqing: Southwest University, 2012: 1-52.
[28] 徐彩彩. 辽河流域河流分类与生境评价研究 [D]. 太原: 山西大学, 2015: 1-38. Xu C C. The study of river classification and river habitat assessment in Liaohe River Basin [D]. Taiyuan: Shanxi University, 2015: 1-38.
[29] 李艳利, 李艳粉, 李科. 不同尺度下人类活动对浑太河流域鱼类和大型底栖动物群落特征的影响 [J]. 环境科学研究, 2016, 29(8): 1145-1153. Li Y L, Li Y F, Li K. Relative influence of anthropogenic stressors on fish and macroinvertebrate communities at different scales in Huntai Rivers [J]. Research of Environmental Sciences, 2016, 29(8): 1145-1153.
[30] 徐军, 王玉玉, 王康, 等. 水域生态学中生物稳定同位素样品采集、处理与保存 [J]. 水生生物学报, 2020, 44(5): 989-997. Xu J, Wang Y Y, Wang K, et al. Protocols for sample collection, pretreatment and preservation of aquatic organisms in stable isotope ecology [J]. Acta Hydrobiologica Sinica, 2020, 44(5): 989-997.
[31] Post D M, Pace M L, Hairston N G. Ecosystem size determines food-chain length in lakes [J]. Nature, 2000, 405(6790): 1047-1049. doi: 10.1038/35016565
[32] Layman C A, Arrington D A, Montaña C G, et al. Can stable isotope ratios provide for community-wide measures of trophic structure [J]? Ecology, 2007, 88(1): 42-48. doi: 10.1890/0012-9658(2007)88[42:CSIRPF]2.0.CO;2
[33] 巴家文, 邓华堂, 段辛斌, 等. 应用稳定性同位素(δ13C、δ15N)技术研究长江中游干流主要鱼类的营养级 [J]. 动物学杂志, 2015, 50(4): 537-546. Ba J W, Deng H T, Duan X B, et al. Trophic level analysis on main fish species in the middle reaches of Yangtze River by δ13C and δ15N analysis [J]. Chinese Journal of Zoology, 2015, 50(4): 537-546.
[34] 邓华堂, 段辛斌, 刘绍平, 等. 大宁河下游主要鱼类营养结构的时空变化 [J]. 生态学报, 2014, 34(23): 7110-7118. Deng H T, Duan X B, Liu S P, et al. Temporal and spatial variations in the trophic structure of key species in downstream of the Daning River [J]. Acta Ecologica Sinica, 2014, 34(23): 7110-7118.
[35] 周正, 黄宇波, 王斌梁, 等. 运用稳定同位素技术分析三峡坝前水域的食物网结构 [J]. 生态科学, 2020, 39(5): 82-90. Zhou Z, Huang Y B, Wang B L, et al. The analysis of food web structure in the area in front of the Three Gorges Dam using the stable isotope technology [J]. Ecological Science, 2020, 39(5): 82-90.
[36] Reckerth A, Stichler W, Schmidt A, et al. Long-term data set analysis of stable isotopic composition in German rivers [J]. Journal of Hydrology, 2017(552): 718-731.
[37] 李斌, 徐丹丹, 吴迪, 等. 基于C、N同位素技术的河流食物网基础碳源研究进展 [J]. 核农学报, 2017, 31(5): 1029-1035. Li B, Xu D D, Wu D, et al. Research on carbon sources of river food webs based on the carbon and nitrogen stable isotopic technique [J]. Journal of Nuclear Agricultural Sciences, 2017, 31(5): 1029-1035.
[38] Xie J, Wang C Y, Liu L, et al. Assessment of aquatic ecological health based on the characteristics of the fish community structures of the Hun River Basin, Northeastern China [J]. Water, 2023, 15(3): 501. doi: 10.3390/w15030501
[39] Wang C Y, Shao J, Ma B S, et al. Longitudinal patterns in fish assemblages after long-term ecological rehabilitation in the Taizi River, Northeastern China [J]. Sustainability, 2022, 14(22): 14973. doi: 10.3390/su142214973
[40] 徐军, 张敏, 谢平. 氮稳定同位素基准的可变性及对营养级评价的影响 [J]. 湖泊科学, 2010, 22(1): 8-20. doi: 10.18307/2010.0102 Xu J, Zhang M, Xie P. Variability of stable nitrogen isotopic baselines and its consequence for trophic modeling [J]. Journal of Lake Sciences, 2010, 22(1): 8-20. doi: 10.18307/2010.0102
[41] Sha Y C, Su G H, Zhang P Y, et al. Diverse dietary strategy of Lake Anchovy Coilia ectenes taihuensis in lakes with different trophic status [J]. Journal of Ichthyology, 2015, 55(6): 866-873. doi: 10.1134/S0032945215060193
[42] 曾艳艺, 杨婉玲, 李海燕, 等. 珠江河网典型段基础碳源对水生动物的贡献 [J]. 生态学杂志, 2018, 37(8): 2403-2410. Zeng Y Y, Yang W L, Li H Y, et al. Contribution of basal carbon sources to aquatic animals in typical sections of the Pearl River [J]. Chinese Journal of Ecology, 2018, 37(8): 2403-2410.
[43] 尹洪洋, 朱文涛, 马文刚, 等. 三亚蜈支洲岛海洋牧场区域夏季食物网研究 [J]. 生态学报, 2022, 42(8): 3241-3253. Yin H Y, Zhu W T, Ma W G, et al. The summer food web in the marine ranch area of Wuzhizhou Island in Sanya, Hainan [J]. Acta Ecologica Sinica, 2022, 42(8): 3241-3253.
[44] 张远, 陈立斌, 渠晓东, 等. 辽宁太子河大型水生植物的群落特征及其与环境的关系 [J]. 植物科学学报, 2011, 29(5): 552-560. Zhang Y, Chen L B, Qu X D, et al. Environmental factors and community characteristics of aquatic macrophytes in Taizi River Tributaries of Liaoning Province [J]. Plant Science Journal, 2011, 29(5): 552-560.
[45] 刘佳. 浑河中上游近60年水文气象要素变化特征及周期演变分析 [J]. 水利技术监督, 2018(4): 242-247. Liu J. Study on changes of hydrological and meteorological factors in the upper and middle reaches of Hunhe river in recent 60 years [J]. Technical Supervision in Water Resources, 2018(4): 242-247.
[46] 曹永强, 张亮亮, 袁立婷. 辽宁省植被生长季NDVI对气候因子的响应 [J]. 植物学报, 2018, 53(1): 82-93. Cao Y Q, Zhang L L, Yuan L T. Correlation analysis of normalized difference vegetation index (NDVI) and climatic factors in the vegetative growing season in Liaoning Province [J]. Chinese bulletin of botany, 2018, 53(1): 82-93.
-
期刊类型引用(1)
1. 巫曦雅,龚海波,易霈霈,傅雪军,余进祥,余智杰,孟子豪,黄滨,王淑兵. 上犹江水库鲢鳙生态增殖区与禁捕区鱼类群落营养结构差异的研究. 淡水渔业. 2024(05): 13-23 .  百度学术
百度学术
其他类型引用(1)

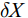
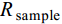

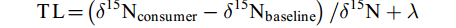

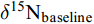
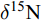
计量
- 文章访问数: 4417
- HTML全文浏览量: 722
- PDF下载量: 112
- 被引次数: 2