
COMMUNITY STRUCTURE AND ENVIRONMENTAL ANALYSIS OF MACROZOOBENTHOS IN YICHANG-ANQING REACHES OF THE YANGTZE MAINSTEM
-
摘要: 为系统了解目前长江中下游干流大型底栖动物群落结构现状, 于2016年5—6月和10—12月对长江干流宜昌-安庆段进行2次调查。共记录大型底栖动物96种, 水生昆虫种类数最多, 占总数的49.0%。大部分物种(58.7%)的出现频率都较低(<1%), 且各江段种类组成差异较大。大型底栖动物密度为(213±58) ind./m2, 生物量为(0.202±0.066) g/m2。大型底栖动物群落结构汛前汛后差异显著, 汛后种类数和现存量均明显下降。与建坝前资料相比, 大型底栖动物群落结构发生较大改变, 种类数增加, 现存量下降。环境分析表明影响大型底栖动物分布的主要因素是流速, 底质和水质的影响不大。大型底栖动物现存量下降与建坝后干流冲刷加剧有关, 针对底栖动物保护, 建议开展水沙调节、营造缓流生境并加强洲滩保护。Abstract: To systematically understand the community structure of macrozoobethos in the mid-lower Yangtze mainstem, two field investigations were conducted on 13 sections along Yichang-Anqing reaches in May-June and October-December 2016, respectively. A total of 96 taxa of macrozoobenthos were recorded with the most abundant group of aquatic insects that account for 49.0% of the total. The frequency of occurrence was relatively low (<1%) for most species (58.7%), and species composition were obviously different among reaches. The density of macrozoobenthos was (213±58) ind./m2, and the biomass was (0.202±0.066) g/m2. Community structure was different between pre-flood and post-flood periods, where obvious decreases in both species number and standing crops were found in the latter. Compared with the pre-damming data, community structure of macrozoobenthos had increased species number and decreased standing crops. Environmental analyses showed that velocity was the key factor affecting the distribution of macrozoobenthos, while substrate and water quality parameters were of little importance. The sharp decline in standing crops of macrozoobenthos was closely related to the increased scouring effects in post-damming period. These results suggest to carry out regulation of flow and sediment, to create slow-flow habitats and to strengthen protection of sand bars with regard to macrozoobenthos conservation in the Yangtze mainstem.
-
Keywords:
- Macrozoobenthos /
- Community Structure /
- Yangtze mainstem /
- Velocity /
- Scouring
-
大银鱼(Protosalanx hyalocranius Abbott, 1901)和太湖新银鱼(Neosalanx taihuensis Chen, 1956)分别隶属于鲑形目(Salmoniformes)银鱼科(Salangidae)的大银鱼属和新银鱼属, 主要分布在我国黄河、淮河和长江中下游及其附属湖泊, 是我国重要的经济鱼类[1]。20世纪70年代以来, 2种银鱼被引种移植到全国众多湖泊和水库[2]。大银鱼主要被移植到长江以北[3], 太湖新银鱼则多被移植到黄河以南[4]。2种银鱼的生命周期均为一年, 个体小、生长快、世代离散, 种群波动显著 [1, 5]。
过度捕捞是影响鱼类资源可持续利用的重要因素, 分为生长型过度捕捞和补充型过度捕捞[6]。由于银鱼世代离散, 补充型过度捕捞导致剩余产卵群体数量不足, 进而使种群下一世代数量减少, 被认为是其资源量下降的重要原因[7]。单位补充量产卵群体生物量(Spawning Biomass-recruitment, SBR)模型根据种群的生长、死亡参数和种群资源动态, 估算捕捞后产卵群体的残存量, 进而评估种群的资源利用状况[8, 9]。当SBR残存量低于未开发状态的20%时, 通常视为补充型过度捕捞, 可能导致种群崩溃[10, 11]。平衡产量模型是渔业资源评估最常用的Beverton-Holt模型的简化形式[6, 12], 常用于分析开捕时间和捕捞强度对渔获产量的影响, 并据此制定合理的渔业管理方案[13, 14]。
洪泽湖(33°06—40′N, 118°10—52′E)是中国第四大淡水湖, 水域面积约2069 km2。大银鱼和太湖新银鱼是洪泽湖最主要的经济鱼类之一, 2010年以来年渔产量最高曾达106 kg[15], 但年际波动较大, 且近年来呈现下降趋势(江苏省洪泽湖渔业管理委员会办公室, 未发表数据)。洪泽湖银鱼捕捞采用银鱼拖网进行, 每年1月1日至8月8日为禁捕期, 其中5月21日左右允许捕捞1周。据渔民反映, 在每年12月下旬捕捞期的最后阶段2种银鱼均已很难捕捞、渔获量极低。本研究对洪泽湖大银鱼和太湖新银鱼的周年生长和资源量动态进行了分析, 评估种群捕捞利用状况, 进而优化渔业管理方式, 以期为洪泽湖银鱼的资源可持续利用管理提供参考。
1. 材料与方法
1.1 采样水域与采样方法
2015年7月至2016年6月每月中旬, 对洪泽湖大银鱼和太湖新银鱼进行周年采样(图 1)。采样使用渔民用的银鱼拖网进行, 拖网网口宽11.6 m, 高2.3 m; 网体长约25 m, 由4个网片组成, 侧面网片网目为1.2 cm×1.2 cm, 上下网片网目为1.5 cm×1.5 cm; 末端网袋长1.3 m, 网目为0.3 cm×0.3 cm。采样时在机动渔船左右两侧各设置一个拖网, 拖网上沿恰好没入水中, 渔船在固定起点(A, 33°17′19″N, 118°47′17″E)和终点(B, 33°19′58″N, 118°46′50″E)间拖行5 km, 拖速为4.0 km/h。采样在上午9:00—12:00间进行。样品采集后立即置于冰水保温箱中, 带回实验室计数, 进行生物学测定。每次采样2种银鱼生物学测定数量均在100尾以上, 当样品数量不足时, 增加拖行时间。
![]() 图 1 洪泽湖, 示大银鱼和太湖新银鱼采集时拖网拖行区间起点(A)和终点(B)Figure 1. Hongze Lake, showing the trawling area (from A to B) for P. hyalocranius and N. taihuensis collection
图 1 洪泽湖, 示大银鱼和太湖新银鱼采集时拖网拖行区间起点(A)和终点(B)Figure 1. Hongze Lake, showing the trawling area (from A to B) for P. hyalocranius and N. taihuensis collection1.2 数据获取与分析
体长-体重关系和生长率 在实验室对大银鱼和太湖新银鱼的渔获物进行计数和称重。测定个体体长(L, 0.1 mm), 称体重(W, 0.01 g), 解剖鉴定性别, 繁殖季节肉眼确定雌性性腺发育分期[16, 17]。体长-体重关系采用W=a×Lb进行拟合。采用单因子方差分析(ANOVA)检验各月2种银鱼体长和体重差异显著性, 并采用Bonferroni方法进行多重比较。分别求出2种银鱼各月的平均体长和体重, 计算相邻月份间体长和体重瞬时生长率(gl, gw)[18]:
${g_l}{\rm{ = ln}}{L_{n + 1}}{\rm{ - ln}}{L_n}$

${g_w}={\rm{ln}}{W_{n + 1}} - {\rm{ ln}}{W_n}$

式中, Ln 和Wn 分别为某月(n)大银鱼或太湖新银鱼的平均体长和体重; Ln+1 和Wn+1则为下一月份的平均体长和体重。
最适生长方程 洪泽湖大银鱼的繁殖季节为12至次年2月, 盛期为1月中旬; 太湖新银鱼春群的繁殖季节为3—4月, 盛期为3月(未发表数据)。据此, 我们将1月25日和3月25日分别设为大银鱼和太湖新银鱼的出生时间, 即0龄, 并推算2种银鱼各月采样时间对应的年龄。采用Logistic生长方程(Logistic growth function, Logistic GF)、Gompertz生长方程(Gompertz growth function, Gompertz GF)和von Bertalanffy生长方程(Von Bertalanffy Growth Function, VBGF)拟合2种银鱼的年龄与体长生长关系, 使用最小二乘法估算各生长方程的参数, 依据赤井信息准则(Akaike’s information criterion, AIC)选择最适生长方程[19]。3种生长方程分别为:
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
式中, t表示样本年龄; t0为理论生长起点, 即体长Lt=0时的起始年龄; L∞为渐近体长; K为生长系数。生长方程拟合采用R-3.3.1软件进行[20]。
种群死亡系数 定义单位捕捞努力量渔获量(Catch per Unit Effort, CPUE)为拖网拖行1 km2水域面积的渔获物个体数量和生物量。使用CPUE逐月数量变动数据, 结合洪泽湖当前的银鱼捕捞管理方式(8月9日—12月31日为捕捞期, 同时5月21—28日可对银鱼资源进行为期一周的捕捞), 求解以下方程组, 计算捕捞死亡系数F和自然死亡系数M [21]:
${N_{{{t_2}}}} = \left\{ {\begin{array}{*{20}{c}}\!\!\!\! {{N_{{{t_1}}}}{{\rm{e}}^{ - M({{t_2}} - {{t_1}})}}\;\;\left( \text{当不存在人为捕捞作用时} \right)}\\\!\!\!\! {{N_{{{t_1}}}}{{\rm{e}}^{ - Z({{t_2}} - {{t_1}})}}\;\;\left( \text{当存在人为捕捞作用时} \right)}\end{array}} \right.$

This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
模型构建 使用平衡产量模型(The equilibrium output model)估算种群单位补充量渔产量随开捕年龄和捕捞强度的变化[6, 13]:
$\begin{aligned}Y = & F \times {N_0} \times {{\rm{e}}^{ - M \times {{r}}}} \times {W_\infty } \times \\ & \left( {\frac{1}{Z} - \frac{{3{{\rm{e}}^{ - K \times {{r}}}}}}{{Z + K}} + \frac{{3{{\rm{e}}^{ - 2K \times {{r}}}}}}{{Z + 2K}} - \frac{{{{\rm{e}}^{ - 3K \times {{r}}}}}}{{Z + 3K}}} \right)\end{aligned}$

This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
使用SBR模型对洪泽湖当前银鱼捕捞管理方式和依据平衡产量模型制定的新的管理方式进行评估。假定2种方案下银鱼的捕捞死亡系数和自然死亡系数不变, 将模型中的时间步长设置为1周, 即0.0192 year[10], SBR模型公式为[8]:
$S \! B \! R \!\!=\!\! \sum\limits_{t = 0}^{{t_{\max }}} \!{\exp [( \!-\! (F \! \times \!\! {S_t}} \! \times \! {A_t}) \!-\! M) \!\! \times \!\! t] \!\! \times \!\! a \!\! \times \!\! {({L_t})^b} \!\! \times\! \! {G_t}$

式中, tmax表示样本的最大年龄; St和Gt分别表示刀刃式选择假设下的网具和性成熟选择性, 当t<50%性成熟体长时,Gt=0, 反之, Gt=1; 当t<初次开捕年龄时,St=0, 反之, St=1[22]; At对应于t龄时是否处于渔业开捕状态, 若是, 则At=1, 否则At=0。
对于SBR模型中使用的50%性成熟体长, 针对2种银鱼1年生且1次性成熟的特点, 以繁殖期性成熟的最小10尾雌性个体的体长均值Lm表示[23], 将其代入最适生长方程中, 计算对应的性成熟年龄Tm。
2. 结果
2.1 种群结构与生长
共测定了洪泽湖1250尾大银鱼和1818尾太湖新银鱼的体长和体重。大银鱼体长和体重分别为29.0—182.6 mm和0.10—34.79 g; 太湖新银鱼体长和体重分别为20.4—82.7 mm和0.04—3.40 g。银鱼的体长-体重关系方程分别为:
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
![]() 图 2 洪泽湖大银鱼(a)和太湖新银鱼(b)的体长频率分布逐月变化Figure 2. Monthly variations in body length frequency distribution for P. hyalocranius (a) and N. taihuensis (b) in Hongze Lake
图 2 洪泽湖大银鱼(a)和太湖新银鱼(b)的体长频率分布逐月变化Figure 2. Monthly variations in body length frequency distribution for P. hyalocranius (a) and N. taihuensis (b) in Hongze Lake大银鱼一个世代在渔获物中出现的时间从4月到翌年2月, 9月以后大银鱼个体间体长差异逐渐增大、体长范围逐渐增加, 12月后大个体在种群中所占比例下降(图 2a)。世代周期中存在4—6月和8—11月2个快速生长阶段。这2个阶段相邻月份的体长和体重均差异显著(P<0.05)。体长和体重均在11月达到最大值。体长和体重瞬时生长率总体均呈下降趋势, 12月后为负值(图 3、图 4)。
太湖新银鱼一个世代在渔获物中出现的时间从5月到翌年4月, 为春季繁殖群体(春群)的后代; 9月也采集到少量性成熟个体(秋群), 其后代于10月开始出现在渔获物中, 但数量较少(图 2b)。本研究仅考虑春群。太湖新银鱼整个世代周期中持续生长, 无明显的阶段性。5—12月间相邻月份间体长差异显著(P<0.05), 但体重差异不显著(P>0.05)。太湖新银鱼体长和体重瞬时生长率变化较大银鱼平稳, 总体呈下降趋势, 12月后近乎于零(图 3、图 4)。
![]() 图 3 洪泽湖大银鱼和太湖新银鱼(不包括秋群)的体长与其瞬时生长率的逐月变化(误差线代表标准差; *表示两个相邻月份间体长差异显著, P<0.05)Figure 3. Monthly fluctuations in body length and its instantaneous growth rate for P. hyalocranius and N. taihuensis in Hongze Lake (Error bars represent ± standard deviation; * indicates significant difference in body length between two adjacent months, P<0.05)
图 3 洪泽湖大银鱼和太湖新银鱼(不包括秋群)的体长与其瞬时生长率的逐月变化(误差线代表标准差; *表示两个相邻月份间体长差异显著, P<0.05)Figure 3. Monthly fluctuations in body length and its instantaneous growth rate for P. hyalocranius and N. taihuensis in Hongze Lake (Error bars represent ± standard deviation; * indicates significant difference in body length between two adjacent months, P<0.05)![]() 图 4 洪泽湖大银鱼和太湖新银鱼(不包括秋群)的体重与其瞬时生长率的逐月变化(误差线代表标准差; *表示两个相邻月份间体重差异显著, P<0.05)Figure 4. Monthly fluctuations in body weight and its instantaneous growth rate for P. hyalocranius and N. taihuensis in Hongze Lake (Error bars represent ± standard deviation; * indicates significant difference in body weight between two adjacent months, P<0.05)
图 4 洪泽湖大银鱼和太湖新银鱼(不包括秋群)的体重与其瞬时生长率的逐月变化(误差线代表标准差; *表示两个相邻月份间体重差异显著, P<0.05)Figure 4. Monthly fluctuations in body weight and its instantaneous growth rate for P. hyalocranius and N. taihuensis in Hongze Lake (Error bars represent ± standard deviation; * indicates significant difference in body weight between two adjacent months, P<0.05)2.2 最适生长方程与性成熟大小
在3种生长方程中, VBGF对大银鱼生长拟合的AIC值最小, Logistic GF对太湖新银鱼生长拟合的AIC值最小(表 1), 分别为2种鱼类的最适生长方程(图 5)。大银鱼和太湖新银鱼卵巢发育至Ⅳ期的10尾最小个体的体长均值Lm分别为99.9和61.1 mm, 代入对应的生长方程, 得出大银鱼和太湖新银鱼的Tm分别为0.528 year和0.564 year。
表 1 三种生长方程对大银鱼和太湖新银鱼生长拟合的方程参数Table 1. Estimated parameters of three different growth functions for P. hyalocranius and N. taihuensis in Hongze Lake参数Parameter 大银鱼P. hyalocranius 太湖新银鱼N. taihuensis Logistic GF Gompertz GF VBGF Logistic GF Gompertz GF VBGF L∞ (mm) 145.86 153.63 173.35 66.82 67.24 67.87 K 5.013 3.536 1.972 5.386 4.569 3.748 t0 (year) 0.376 0.290 0.092 0.124 0.054 –0.042 AIC 7237.47 7220.83 7202.66 12424.96 12428.04 12432.73 ![]() 图 5 洪泽湖大银鱼(a)和太湖新银鱼(b)的最适生长方程曲线Figure 5. The most fitted growth functions for P. hyalocranius (a) and N. taihuensis (b) in Hongze Lake
图 5 洪泽湖大银鱼(a)和太湖新银鱼(b)的最适生长方程曲线Figure 5. The most fitted growth functions for P. hyalocranius (a) and N. taihuensis (b) in Hongze Lake2.3 CPUE逐月变动与种群死亡系数
单位水域面积(1 km2)中大银鱼和太湖新银鱼渔获物数量均在5月最高, 分别为31809和78098 ind./km2。经历了5月中旬为期1周的捕捞后, 6月2种银鱼的资源数量均明显下降; 6月至8月间下降相对平缓; 8月捕捞开始后下降明显, 到12月时渔获物数量均降至500 ind./km2以下。单位水域面积(1 km2)大银鱼和太湖新银鱼渔获物生物量的最高值分别出现在6月和8月, 为73.07和49.33 kg/km2。大银鱼生物量在8—9月之间下降最快,太湖新银鱼生物量在9—10月间下降最快。
根据洪泽湖现行的捕捞管理方式, 5—6月间和8月以后, 2种银鱼的CPUE下降受捕捞和自然死亡的共同影响; 6—8月间, CPUE的下降只受自然死亡的影响。据此求出大银鱼F和M分别为8.583/year和3.292/year,太湖新银鱼F和M分别为7.006/year和1.146/year。
![]() 图 6 洪泽湖大银鱼和太湖新银鱼(春群)单位水域面积(1 km2)渔获物数量(a)和生物量(b)的逐月变动4月之前2种银鱼的个体小、渔获数量少, 均未计算CPUEFigure 6. Monthly variations in number of individuals (a) and biomass (b) for P. hyalocranius and N. taihuensis harvested across an area of 1 km2 by trawl in Hongze LakeIn April, size of both species was small, and numbers of catch for both species was very low. Thus, data in April was not included
图 6 洪泽湖大银鱼和太湖新银鱼(春群)单位水域面积(1 km2)渔获物数量(a)和生物量(b)的逐月变动4月之前2种银鱼的个体小、渔获数量少, 均未计算CPUEFigure 6. Monthly variations in number of individuals (a) and biomass (b) for P. hyalocranius and N. taihuensis harvested across an area of 1 km2 by trawl in Hongze LakeIn April, size of both species was small, and numbers of catch for both species was very low. Thus, data in April was not included2.4 平衡产量模型分析
使用以上获得的生长、繁殖和死亡等相关参数(表 2), 分别构建了大银鱼和太湖新银鱼单位补充量渔产量随开捕年龄(图 6a)和捕捞死亡系数(图 6b)变化的曲线。结果显示, 大银鱼和太湖新银鱼的单位补充量渔产量均随开捕年龄的增大呈先升后降的倒U型曲线关系。当大银鱼开捕年龄tc=0.525 year时, 能够取得相应最大的单位补充量渔产量Ymax=1.078 kg, 开捕年龄过早或过晚均不能获得足够的渔产量。就洪泽湖当前渔业管理模式来说, 大银鱼的开捕年龄约为0.538 year, 略大于tc, 此时的单位补充量渔产量Y=1.077 kg, 接近Ymax。当太湖新银鱼开捕年龄tc=0.525 year时, 能够取得相应最大的单位补充量渔产量Ymax=0.684 kg。就当前渔业管理模式来说, 太湖新银鱼的开捕年龄约为0.365 year, 此时的单位补充量渔产量Y=0.607 kg<Ymax。将开捕时间向后推移20d (即8月29日), 即大银鱼和太湖新银鱼的开捕年龄分别为0.593 year和0.420 year时, 能够获得最大的总单位补充量渔产量1.708 kg。
![]() 图 7 平衡产量模型估算的洪泽湖大银鱼和太湖新银鱼单位补充量渔产量随开捕年龄(a)和捕捞死亡系数(b)变化的曲线垂直虚线表示当前捕捞管理模式下2种银鱼的开捕年龄和捕捞死亡系数Figure 7. Relationships of yield per-recruitment to the age at first capture (a) and fishing mortality coefficient (b) estimated from the equilibrium output mode for P. hyalocranius and N. taihuensis in Hongze LakeThe dotted vertical lines locate the current age at first capture (a) and fishing mortality (b) for both species
图 7 平衡产量模型估算的洪泽湖大银鱼和太湖新银鱼单位补充量渔产量随开捕年龄(a)和捕捞死亡系数(b)变化的曲线垂直虚线表示当前捕捞管理模式下2种银鱼的开捕年龄和捕捞死亡系数Figure 7. Relationships of yield per-recruitment to the age at first capture (a) and fishing mortality coefficient (b) estimated from the equilibrium output mode for P. hyalocranius and N. taihuensis in Hongze LakeThe dotted vertical lines locate the current age at first capture (a) and fishing mortality (b) for both species大银鱼和太湖新银鱼的单位补充量渔产量均先随捕捞死亡系数的增加而快速增高, 当捕捞死亡系数增大到一定程度后, 渔产量上升速度减慢, 之后不再升高甚至出现下降趋势。大银鱼捕捞死亡系数F=24.38/year时, 获得最大单位补充量渔产量Ymax=1.114 kg; 当前洪泽湖大银鱼捕捞死亡系数为8.583/year, 相应渔产量Y=1.077 kg, 接近Ymax且尚未开发过度。太湖新银鱼捕捞死亡系数F=4.72/year时, 获得最大的单位补充量渔产量Ymax=0.616 kg; 而当前洪泽湖太湖新银鱼捕捞死亡系数为7.006/year, 已大于最适捕捞强度, 且相应的渔产量Y=0.607 kg, 小于Ymax。
2.5 基于SBR模型分析的资源状况评价
使用以上获得的生长、繁殖和死亡等相关参数(表 2), 分别构建了2种不同渔业管理方案下大银鱼和太湖新银鱼种群的SBR随捕捞死亡系数变动的曲线:
方案1: 洪泽湖现行的渔业管理方式。8月9日—12月31日为捕捞期, 同时5月可对银鱼资源进行为期1周的捕捞。
方案2: 假定对两种银鱼的捕捞强度不发生变化(即F恒定), 将开捕时间向后推迟20d, 8月29日—12月31日为捕捞期, 同时取消5月为期1周的捕捞。
大银鱼在当前渔业管理方案1下, 相对于未开发状态(100%), SBR残存量为20.234%, 即3.326 g, 接近于补充型过度捕捞评价阈值20%, 种群有可能因产卵亲体不足而导致资源衰退; 在方案2下, SBR残存量相当于未开发状态的36.721%, 即6.036 g, 远大于补充型过度捕捞评价阈值20%, 大银鱼渔业资源和产卵群体得到了较好的保护(图 7)。在2种不同管理方案下, 太湖新银鱼SBR残存量分别相当于未开发状态的7.503%和12.862%, 即0.972 g和1.666 g (图 8)。在优化的管理方案2下, 太湖新银鱼的产卵亲体资源虽然得到了一定程度的保护, 但其渔业资源仍处于补充型过度捕捞水平。
表 2 洪泽湖大银鱼和太湖新银鱼平衡产量模型和单位补充量产卵群体生物量模型参数Table 2. Growth and mortality parameters fitted into the equilib-rium output model and SBR model for P. hyalocranius and N. taihuensis in Hongze Lake参数
Parameter大银鱼
P. hyalocranius太湖新银鱼
N. taihuensisa 1.082×10–6 2.113×10–6 b 3.279 3.215 K 1.972 5.386 L∞ (mm) 173.35 66.82 W∞ (g) 23.75 1.56 t0 (year) 0.092 0.124 Z (/year) 11.875 8.152 M (/year) 3.292 1.146 F (/year) 8.583 7.006 tmax (year) 1 1 Lm (mm) 99.9 61.1 Tm (year) 0.528 0.564 tc (year) 0.538 0.365 ![]() 图 8 两种不同捕捞管理方案下洪泽湖大银鱼(a)和太湖新银鱼(b)单位补充量产卵群体生物量随捕捞死亡系数F变化的曲线当F=0时, 大银鱼和太湖新银鱼的单位补充量产卵群体生物量分别为16.44 g和12.95 g。捕捞管理方案1: 洪泽湖现行的渔业管理方式, 即8月9日—12月31日为捕捞期, 同时5月可对银鱼资源进行为期1周的捕捞; 捕捞管理方案2: 假定对两种银鱼的捕捞强度不变, 将开捕时间向后推迟20d, 即8月29日—12月31日为捕捞期, 同时取消5月为期1周的捕捞。虚线示两种管理方案下对应的捕捞死亡系数Figure 8. Relationships of spawner biomass per-recruitment to fishing mortality coefficients for P. hyalocranius (a) and N. taihuensis (b) under two different fishery management schemes in Hongze LakeThe spawner biomass per-recruitment at F=0 is 16.44 g for P. hyalocranius and 12.95 g for N. taihuensis. Fishery management scheme 1 is the current status, with fishing season of one week in Mid-May and between August 9 and December 31; the scheme 2 is an adjusted status, with fishing season between August 29 and December 31. The dotted vertical lines represent the corresponding fishing mortality coefficients under these two fishery management schemes
图 8 两种不同捕捞管理方案下洪泽湖大银鱼(a)和太湖新银鱼(b)单位补充量产卵群体生物量随捕捞死亡系数F变化的曲线当F=0时, 大银鱼和太湖新银鱼的单位补充量产卵群体生物量分别为16.44 g和12.95 g。捕捞管理方案1: 洪泽湖现行的渔业管理方式, 即8月9日—12月31日为捕捞期, 同时5月可对银鱼资源进行为期1周的捕捞; 捕捞管理方案2: 假定对两种银鱼的捕捞强度不变, 将开捕时间向后推迟20d, 即8月29日—12月31日为捕捞期, 同时取消5月为期1周的捕捞。虚线示两种管理方案下对应的捕捞死亡系数Figure 8. Relationships of spawner biomass per-recruitment to fishing mortality coefficients for P. hyalocranius (a) and N. taihuensis (b) under two different fishery management schemes in Hongze LakeThe spawner biomass per-recruitment at F=0 is 16.44 g for P. hyalocranius and 12.95 g for N. taihuensis. Fishery management scheme 1 is the current status, with fishing season of one week in Mid-May and between August 9 and December 31; the scheme 2 is an adjusted status, with fishing season between August 29 and December 31. The dotted vertical lines represent the corresponding fishing mortality coefficients under these two fishery management schemes3. 讨论
3.1 大银鱼和太湖新银鱼的阶段性生长差异
鱼类的生长具有阶段性, 生活史的不同阶段, 个体的生理特征、食性以及环境因素等差异, 会引起生长速度的改变[24, 25]。本研究发现, 洪泽湖大银鱼存在2个快速生长阶段, 分别在4—6月和8—11月间。鱼类一般在早期生活史阶段和性成熟前体长生长较快[26], 这一阶段的快速生长可能是鱼类维持种群数量的一种生态适应方式, 有利于降低其被捕食的压力、快速达到性成熟体长[27, 28]。洪泽湖大银鱼繁殖期在12—2月(作者未发表数据), 4—6月间的第一个快速生长阶段, 在个体性成熟之前; 同时, 该时期水体温度逐步上升, 初级生产力升高, 水体中浮游动物等大银鱼的主要饵料生物丰富, 是这一阶段大银鱼快速生长的营养保障[29]。8—11月间第二阶段快速生长可能主要源于大银鱼的食性转化。唐富江发现兴凯湖大银鱼的体长大于60 mm以后, 逐渐由摄食浮游动物转为摄食鱼虾类[30]; 本文作者也发现, 自8月起虾类和鱼类逐渐取代枝角类和桡足类成为洪泽湖大银鱼的主要食物(作者未发表数据)。鱼虾类相对于浮游动物而言具有体积大、能量高、动物蛋白丰富等特点, 更易于被鱼类吸收和利用, 作为生长和繁殖的能量供应[31]。因此, 大银鱼在8—11月的快速生长很可能是因为有更多的能量来源投入到生长中。12月后, 大银鱼种群体长和体重均表现为负增长, 这可能与其进入繁殖期后大个体较早的繁殖和死亡有关[32], 需进一步研究确证。
太湖新银鱼个体相对较小, 其体长在5—10月呈快速持续生长状态, 无明显阶段性。太湖新银鱼终生摄食浮游动物, 与大银鱼不同, 没有明显的食性转化[33]。太湖新银鱼这一食性特点可能是其生长过程没有明显阶段性的主要原因。11月以后, 太湖新银鱼个体体长无明显的增长趋势。洪泽湖太湖新银鱼繁殖季节主要在3—4月, 11月之后, 繁殖季节前, 是性腺发育能量积累的主要阶段[34]。因此, 能量投入到性腺发育中, 可能是太湖新银鱼11月后体长停止增长的重要原因。
3.2 洪泽湖银鱼资源利用评估与管理
鱼类种群可提供渔获量的多少取决于种群数量、生长、存活、开捕年龄和捕捞强度等, 其中仅有捕捞强度(捕捞死亡系数)和初次捕捞年龄(初次开捕体长)能够被人为控制, 因此渔业管理中常通过调整这2个因素以获取最大可持续渔产量[6]。依据平衡产量模型分析, 洪泽湖大银鱼和太湖新银鱼在当前捕捞强度下的渔产量均已接近其资源群体所能提供的最大渔产量, 调整捕捞强度对进一步提高渔产量的影响不大; 而调整2种银鱼的初次开捕年龄, 更有利于获得较高的总渔产量。
在洪泽湖当前的捕捞管理方式下, 大银鱼和太湖新银鱼的SBR残存量分别为未开发状态的20.23%和7.50%, 表明对2种银鱼的捕捞均处于补充型过度捕捞水平, 且太湖新银鱼捕捞后的SBR残余量已远低于种群崩溃的警戒线。由此看来, 较大的捕捞强度和补充型过度捕捞, 可能是导致近年来洪泽湖大银鱼和太湖新银鱼资源波动明显的重要原因。因此, 很有必要对洪泽湖当前银鱼资源捕捞管理方式进行优化, 避免过度捕捞的发生。
本研究构建平衡产量模型时发现, 将洪泽湖银鱼开捕时间推迟20d可获得大银鱼和太湖新银鱼的最大总渔产量。在渔业管理中, 这种开捕时间的推迟也将影响最终捕捞后的产卵亲体生物量[8, 9]。对洪泽湖的银鱼捕捞管理方式进行优化后, 大银鱼的SBR残存量提高至未开发状态的36.72%, 既满足了获得最高渔产量的需求, 又不会带来太大的渔业风险; 优化管理后太湖新银鱼的SBR残存量亦提高至未开发状态的12.86%。可见, 优化的渔业管理方式实现了获取最高渔产量和保护产卵亲体资源上的“双赢”。同时也应注意到, 优化的渔业管理方案对大银鱼产卵群体的保护作用更为显著; 在未改变当前捕捞强度的情况下, 尽管太湖新银鱼在捕捞后的产卵亲体资源量也得到了一定程度的提升, 但依然低于补充型过度捕捞评价阈值20%, 故还需减小对太湖新银鱼的捕捞强度。因此, 针对洪泽湖太湖新银鱼的渔业资源管理, 在延迟开捕时间的基础上, 建议还可以通过限制渔获物的起捕体长、增大捕捞网具的网目规格等方式控制捕捞强度, 同时可采取人工放流等措施提高其下一代的资源补充能力, 以避免种群资源的进一步衰退。
-
![]()
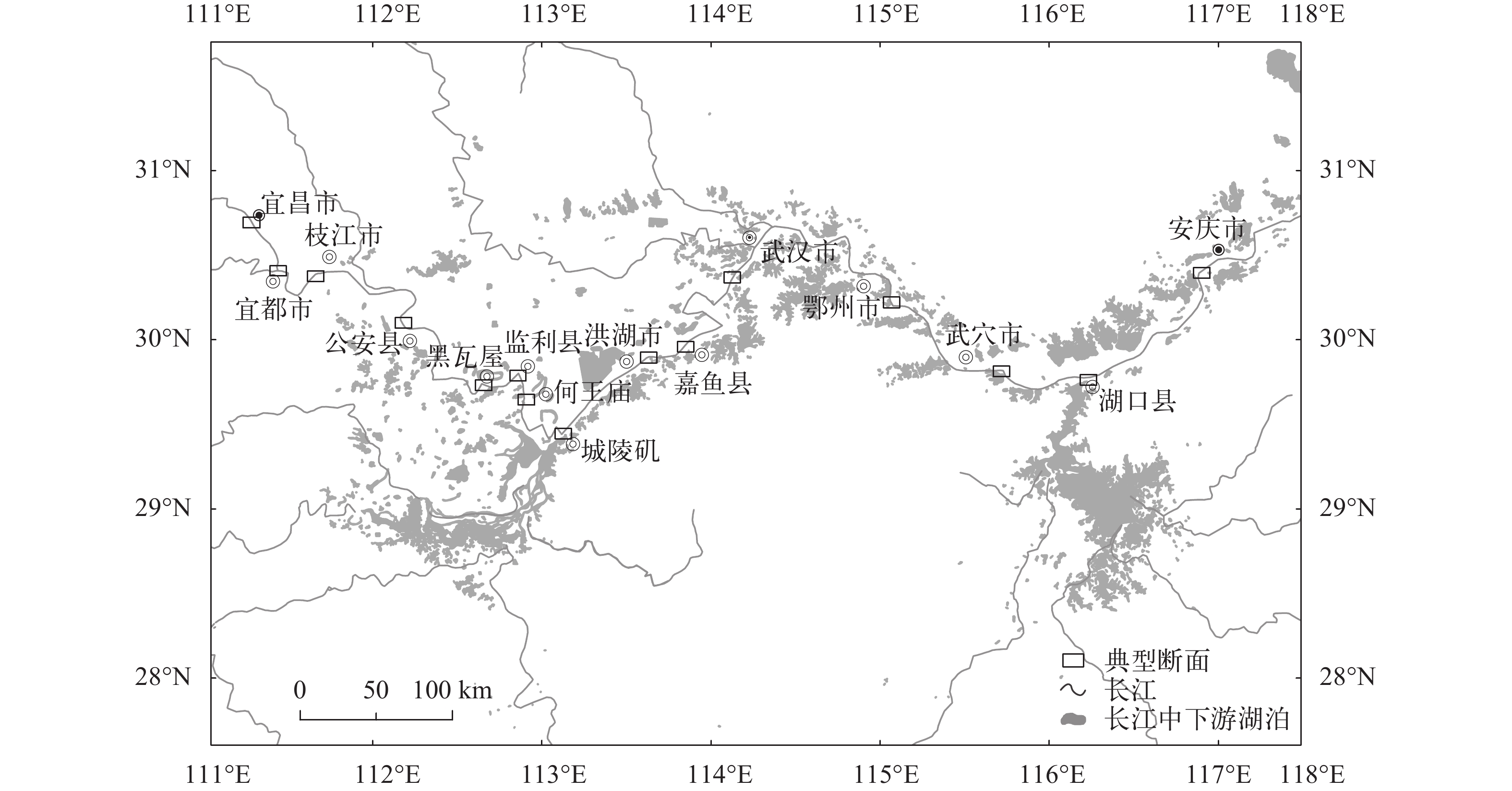
图 1 长江干流大型底栖动物采集江段
Figure 1. Sampling reaches of macrozoobenthos of the Yangtze mainstem
![]()
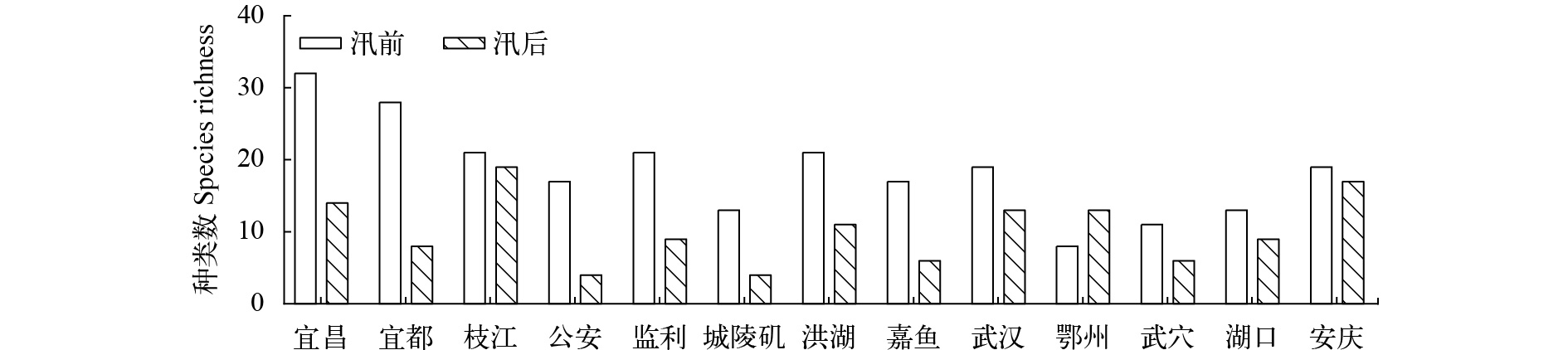
图 2 长江干流各江段大型底栖动物种类数
Figure 2. Species richness of macrozoobenthos in the Yangtze mainstem
![]()
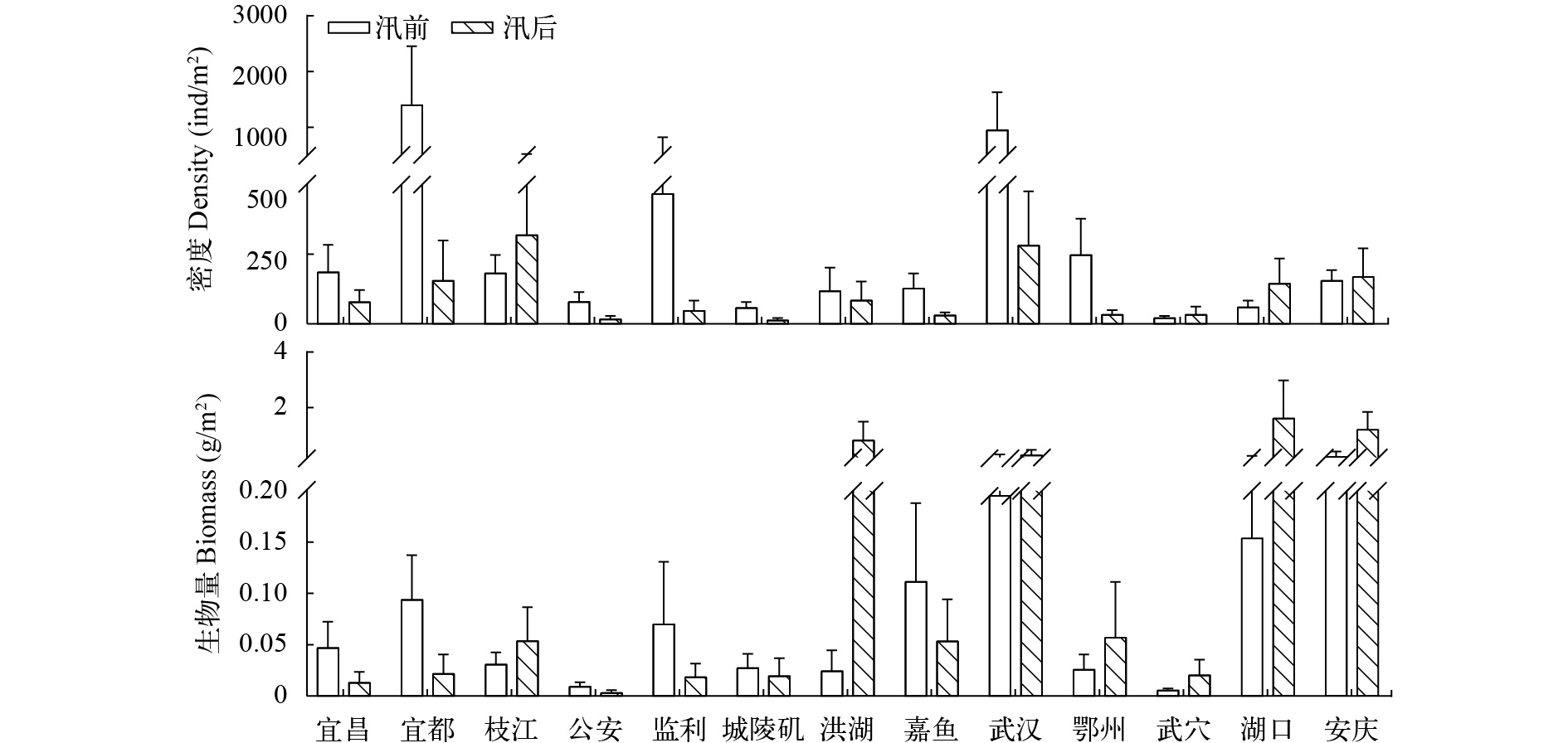
图 3 长江干流各江段大型底栖动物现存量
Figure 3. Standing crops of macrozoobenthos in the Yangtze mainstem
![]()
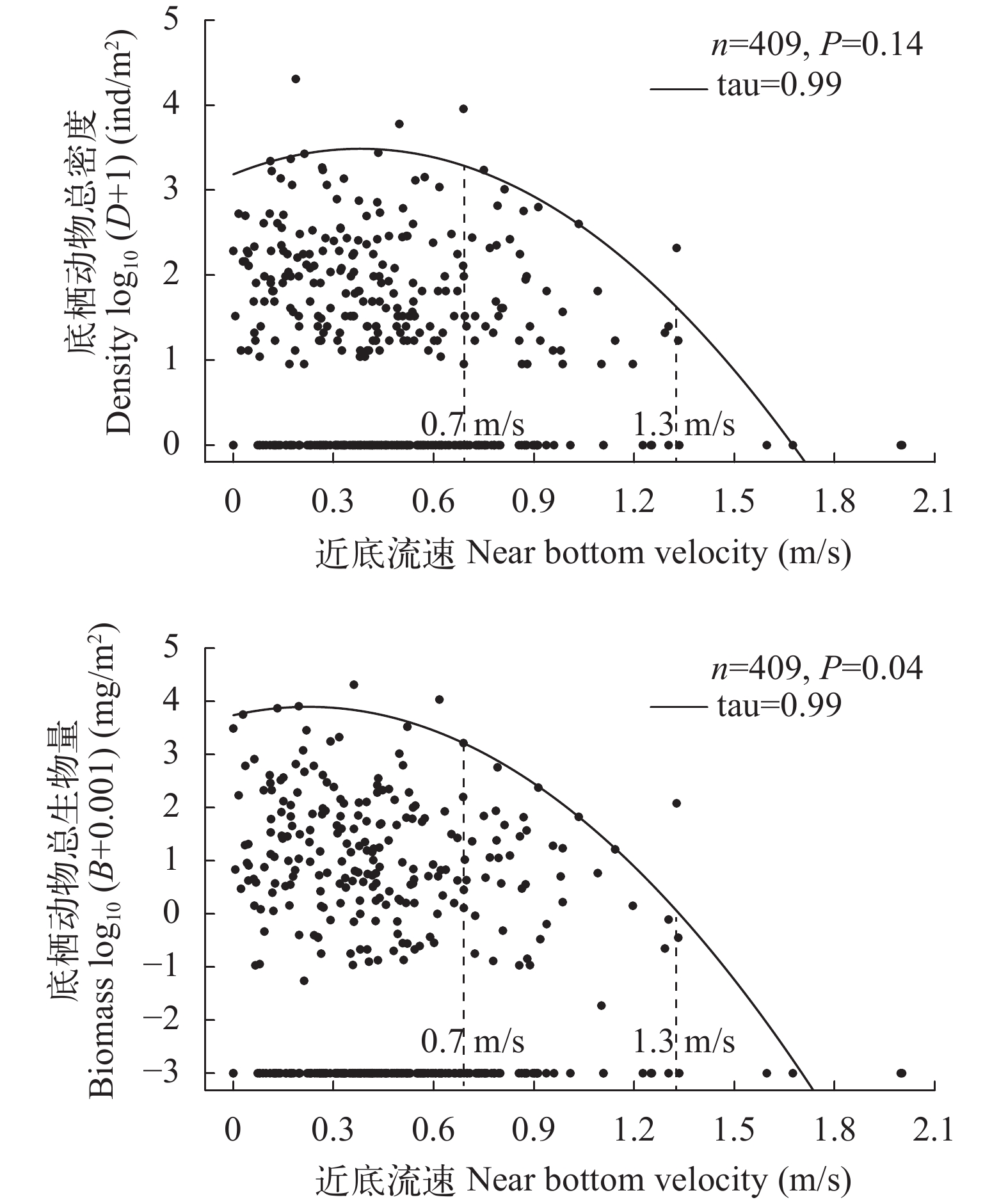
图 4 大型底栖动物现存量与近底流速的关系
Figure 4. The relationships between standing crops of macrozoobenthos and near bottom velocity
![]()
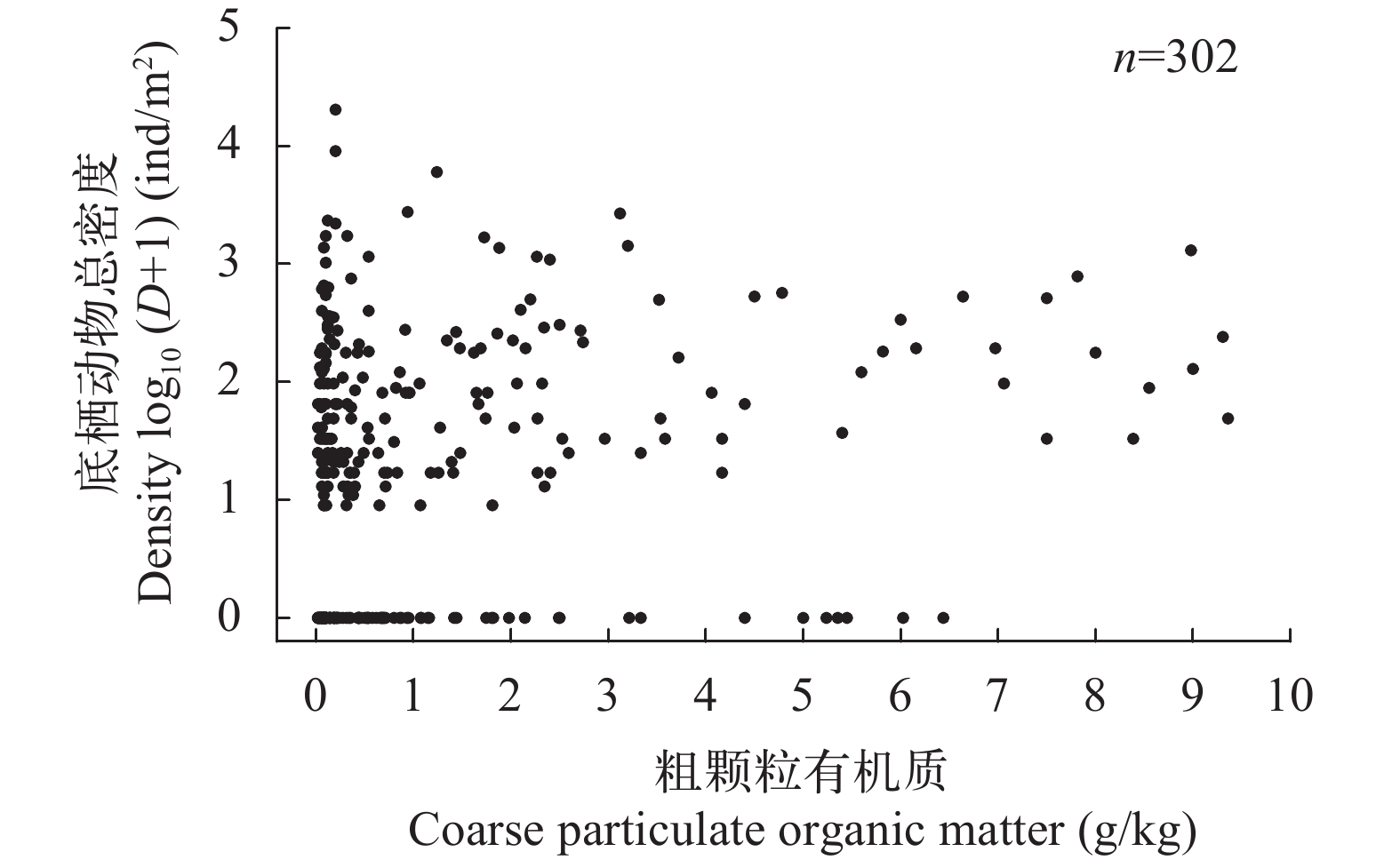
图 5 大型底栖动物密度与粗颗粒有机质的关系
Figure 5. The relationship between density of macrozoobenthos and coarse particulate organic matter of substrate
![]()
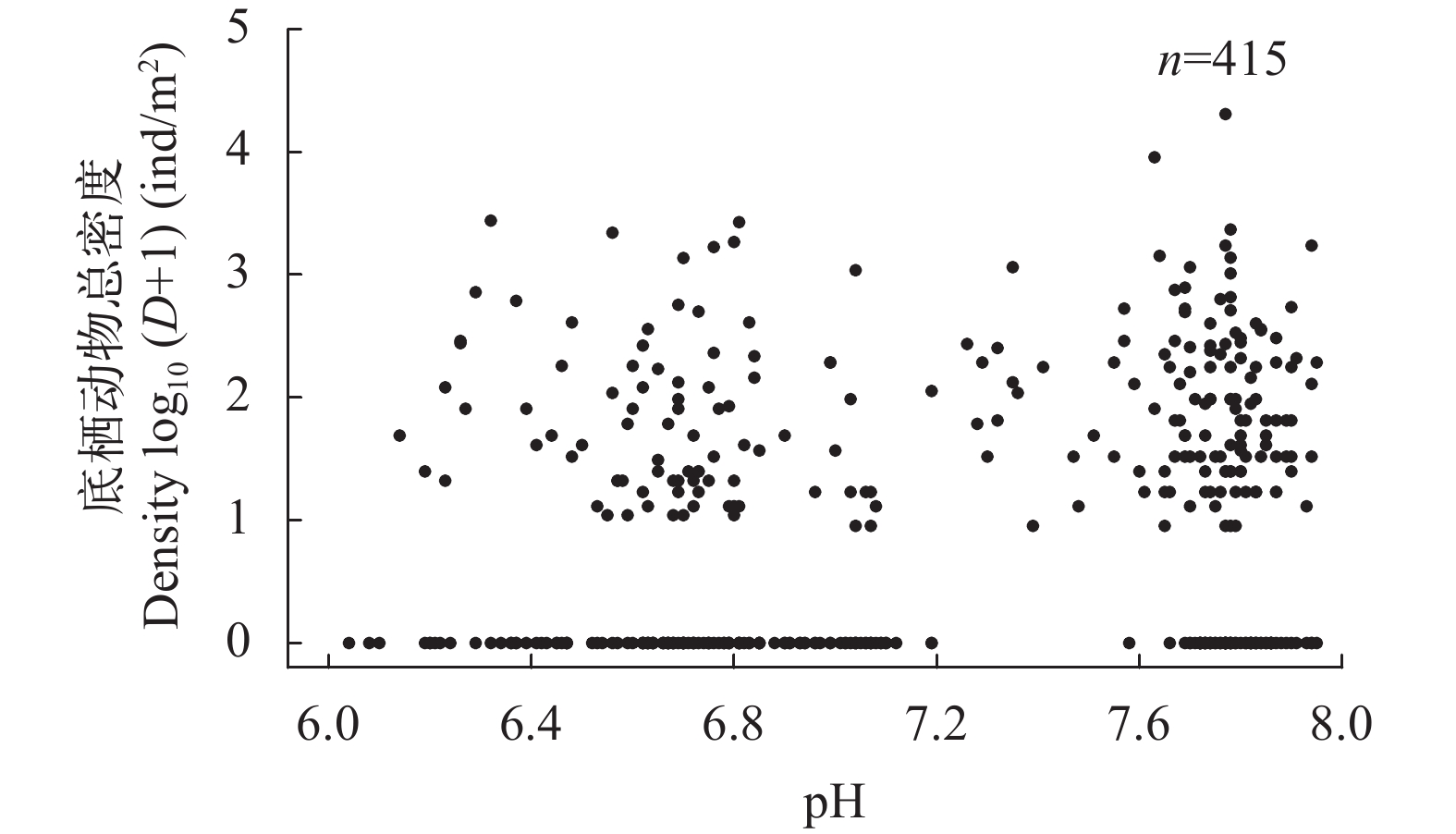
图 6 大型底栖动物密度与表层水pH的关系
Figure 6. The relationship between density of macrozoobenthos and pH of surface water
表 1 长江干流宜昌-安庆段的环境参数
Table 1 Environmental parameters of Yichang-Anqing reaches of the Yangtze mainstem
环境参数Environmental parameters 汛前 汛后 Wilcoxon秩和检验 平均值(最小值—最大值) 平均值(最小值—最大值) W P 水深Water depth (m) 6.4 (0.5—22.5) 4.9 (0.1—16.7) 132 0.01 表层流速Surface velocity (m/s) 0.433 (0—1.755) 0.348 (0—1.943) 109 0.23 近底流速Near bottom velocity (m/s) 0.517 (0—1.597) 0.445 (0—2.002) 119 0.08 平均流速Mean velocity (m/s) 0.561 (0—1.721) 0.512 (0—2.069) 106 0.29 剪切流速Shear velocity (m/s) 0.02 (0—0.061) 0.019 (0—0.075) 106 0.29 中值粒径Medium grain diameter (mm) — 0.63 (0.01—1.1) — — 总氮Total nitrogen (g/kg) — 0.46 (0.08—2.35) — — 总磷Total phosphorus (g/kg) — 0.57 (0.28—0.91) — — 总有机质Total organic matter (g/kg) — 9.3 (0—36.4) — — 粗颗粒有机质Coarse particulate organic matter (g/kg) 1.1 (0—9.4) 0.8 (0—6.4) 96 0.17 透明度Secchi depth (cm) 70 (10—210) 54 (5—170) 101 0.42 水温Water temperature (℃) 20.7 (15.3—25.6) 18.1 (12.8—22.7) 139 0.17 溶解氧Dissolved oxygen (mg/L) 7.4 (5.6—9.4) 8.8 (7.6—10.5) 8 <0.01 电导率Conductivity (s/cm) 282.9 (111.8—367.2) 248.2 (95.6—350.8) 129 0.02 pH 7.74 (7.04—7.95) 6.69 (6.04—7.03) 169 <0.01 注: — 为未测量或未计算, 粗体为达到显著水平(P<0.05)Note: — indicates not measured or not calculated, bold indicates significant correlations (P<0.05)  下载: 导出CSV
下载: 导出CSV
表 2 长江干流大型底栖动物现存量与环境参数的Spearman秩相关关系(达到显著水平以粗体显示, *P<0.05, **P<0.01)
Table 2 Spearman rank correlations between standing crops of macrozoobenthos and environmental parameters (significant correlations in bold, *P<0.05, **P<0.01)
WD SV NBV MV SHV ST D50 TN TP TOM CPOM SD WT DO Cond pH WD SV 0.25** NBV 0.42** 0.41** MV 0.47** 0.42** 0.88** SHV 0.39** 0.42** 0.87** 0.99** ST 0.16** 0.15** 0.13** 0.16** 0.20** D50 –0.06 0.14 0.23 0.24* 0.25* 0.74** TN 0.13 –0.20 –0.30* –0.33** –0.34** –0.72** –0.80** TP 0.00 0.09 –0.04 –0.07 –0.01 –0.03 0.00 0.11 TOM 0.16 –0.29* –0.43** –0.40** –0.41** –0.61** –0.73** 0.79** 0.07 CPOM –0.15** –0.12* –0.16** –0.18** –0.18** –0.34** –0.40** 0.53** 0.01 0.43** SD 0.17** 0.09 0.12* 0.13** 0.16** 0.44** 0.26* –0.20 0.22 –0.19 –0.15* WT 0.02 0.08 0.01 –0.03 –0.01 0.04 0.43** –0.35** 0.33** –0.26* 0.07 0.11* DO –0.10* –0.16** –0.06 –0.03 –0.04 –0.05 –0.49** 0.41** –0.29* 0.36** –0.07 –0.14** –0.77** Cond 0.07 0.18** 0.10 0.12* 0.13** 0.25** 0.39** –0.34** 0.29* –0.36** –0.18** 0.64** 0.18** –0.35** pH 0.11* 0.14** 0.14** 0.10* 0.09 –0.05 –0.06 0.11 –0.08 0.12 –0.13* 0.24** 0.13** –0.50** 0.55** Dtotal –0.08 –0.09 –0.19** –0.21** –0.22** –0.27** –0.29* 0.39** 0.00 0.48** 0.21** –0.02 0.05 –0.17** 0.04 0.17** Btotal –0.09 –0.07 –0.20** –0.22** –0.23** –0.28** –0.36** 0.40** –0.03 0.50** 0.22** –0.08 0.03 –0.14** –0.04 0.13** Doligochaeta –0.07 –0.16** –0.20** –0.20** –0.22** –0.21** –0.23 0.35** –0.10 0.43** 0.16** –0.02 –0.04 –0.08 0.12* 0.11* Boligochaeta –0.06 –0.16** –0.20** –0.20** –0.22** –0.23** –0.23* 0.34** –0.09 0.43** 0.16** –0.06 –0.02 –0.08 0.09 0.09 Dinsect –0.11* –0.12* –0.17** –0.19** –0.19** –0.20** 0.01 0.13 0.23 0.19 0.18** 0.14** 0.14** –0.26** 0.19** 0.29** Binsect –0.10* –0.09 –0.14** –0.17** –0.17** –0.18** 0.01 0.13 0.18 0.18 0.17** 0.15** 0.15** –0.27** 0.19** 0.30** 注: WD. 水深water depth, m; SV. 表层流速surface velocity, m/s; NBV. 近底流速near bottom velocity, m/s; MV. 平均流速mean velocity, m/s; SHV. 剪切流速shear velocity, m/s; ST. 底质类型substrate type; D50. 中值粒径median grain size, mm; TN. 总氮total nitrogen, g/kg; TP. 总磷total phosphorus, g/kg; TOM. 总有机质total organic matter, g/kg; CPOM. 粗颗粒有机质coarse particulate organic matter, g/kg; SD. 透明度Secchi depth, cm; WT. 水温water temperature, ℃; DO. 溶解氧dissolved oxygen, mg/L; Cond. 电导率conductivity, s/cm
下载: 导出CSV
-
[1] Poff N L R, Allan J D, Bain M B, et al. The natural flow regime [J]. BioScience, 1997, 47(11): 769—784 doi: 10.2307/1313099
[2] Lytle D A, Poff N L R. Adaptation to natural flow regimes [J]. Trends in Ecology & Evolution, 2004, 19(2): 94—100
[3] 中国工程院三峡工程阶段性评估项目组. 三峡工程阶段性评估报告(综合卷), 北京: 中国水利水电出版社. 2010, 1—400 The Project Team of Three Gorges Project Stage Assessment of Chinese Academy of Engineering. Three Gorges Project Stage Assessment Report (Integrated Volume) [M]. Beijing: China Water Power Press. 2010, 1—400
[4] 施勇. 三峡工程运用后对长江中下游防洪的影响评价研究. 武汉: 长江出版社. 2011, 1—167 Shi Y. Study on the Impact Assessment of Flood Control of the Mid-Lower Reaches of the Yangtze River after the Application of the Three Gorges Project [M]. Wuhan: Changjiang Press. 2011, 1—167
[5] Thomson J R, Lake P S, Downes B J. The effect of hydrological disturbance on the impact of a benthic invertebrate predator [J]. Ecology, 2002, 83(3): 628—642 doi: 10.1890/0012-9658(2002)083[0628:TEOHDO]2.0.CO;2
[6] Holt C R, Pfitzer D, Scalley C, et al. Macroinvertebrate community responses to annual flow variation from river regulation: an 11- year study [J]. River Research and Applications, 2015, 31(7): 798—807 doi: 10.1002/rra.v31.7
[7] 梁彦龄, 吴天惠, 谢志才. 保安湖底栖动物现状及渔业评价-草型湖泊资源环境与渔业生态学管理. 北京: 科学出版社. 1995, 178—192 Liang Y L, Wu T H, Xie Z C. Status of Macrozoobenthos and Assessment of Fisheries in Baoan Lake - Management of Grass - Type Lake Resources, Environment and Fishery Ecology [M]. Beijing: Science Press. 1995, 178—192
[8] Covich A P, Palmer M A, Crowl T A. The role of benthic invertebrate species in freshwater ecosystems: zoobenthic species influence energy flows and nutrient cycling [J]. BioScience, 1999, 49(2): 119—127 doi: 10.2307/1313537
[9] 舒凤月, 王海军, 潘保柱, 等. 长江中下游湖泊贝类物种濒危状况评估. 水生生物学报, 2009, 33(6): 1051—1058 Shu F Y, Wang H J, Pan B Z, et al. Assessment of species status of Mollusca in the min-lower Yangtze Lakes [J]. Acta Hydrobiologica Sinica, 2009, 33(6): 1051—1058
[10] Xie Z C, Liang Y L, Wang J, et al. Preliminary studies of macroinvertebrates of the mainstream of the Changjiang (Yangtze) River [J]. Acta Hydrobiologica Sinica, 1999, 23(supp1.): 148—157
[11] 段学花, 王兆印, 余国安. 以底栖动物为指示物种对长江流域水生态进行评价. 长江流域资源与环境, 2009, 18(3): 241—247 doi: 10.3969/j.issn.1004-8227.2009.03.007 Duan X H, Wang Z Y, Yu G A. Ecological assessment of the Yangtze River eco-system with benthic invertebrate as indicator species [J]. Resources and Environment in The Yangtze Basin, 2009, 18(3): 241—247 doi: 10.3969/j.issn.1004-8227.2009.03.007
[12] Pan B Z, Wang H Z, Ban X, et al. An exploratory analysis of ecological water requirements of macroinvertebrates in the Wuhan branch of the Yangtze River [J]. Quaternary International, 2015, 380—381: 256—261
[13] 李莎, 熊飞, 王珂, 等. 长江中游透水框架护岸工程对底栖动物群落结构的影响. 水生态学杂志, 2015, 36(6): 72—79 Li S, Xiong F, Wang K, et al. Effect of tetrahedron permeable frames on the community structure of benthic macroinvertebrates in the middle Yangtze River [J]. Journal of Hydroecology, 2015, 36(6): 72—79
[14] 陈秀粉, 夏炜, 潘保柱, 等. 长江中游宜昌至武汉段底栖动物群落结构特征研究. 北京大学学报(自然科学版), 2017, 53(5): 973—981 Chen X F, Xia W, Pan B Z, et al. Community structure characteristics of macroinvertebrate from Yichang to Wuhan in the middle reaches of the Yangtze River [J]. Acta Scientiarum Naturalium Universutatis Pskiensis, 2017, 53(5): 973—981
[15] 赵伟华. 中国河流底栖动物宏观格局及黄河下游生态需水研究. 博士学位论文, 中国科学院水生生物研究所, 武汉. 2010 Zhao W H. Macroecological patterns of macrozoobenthos in rivers of China and environmental flow requirements in lower reaches of the Yellow River [D]. Thesis for Doctor of Science, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan. 2010
[16] 《中国河湖大典》编纂委员会. 中国河湖大典-长江卷(上). 北京: 中国水利水电出版社. 2010. 1—510 Editorial Committee of Encyclopedia of River and Lakes in China. Encyclopedia of River and Lakes in China - Section of Changjiang River Basin (Vol. One) [M]. Beijing: China Water Power Press. 2010. 1—510
[17] 中华人民共和国水利部. 2016年中国河流泥沙公报. 北京: 中国水利水电出版社. 2016, 1—23 The Ministry of Water Resources of the People’s Republic of China. Chinese River Sediment Bulletin of 2016 [M]. Beijing: China Water Power Press. 2016, 1—23
[18] 大连水产学院. 淡水生物学-上册(分类学部分). 北京: 农业出版社. 1982, 1—346 Dalian Institute of Aguaculture. Freshwater Biology-Volume 1 (Taxonomy) [M]. Beijing: China Agriculture Press. 1982, 1—346
[19] 何雪宝. 西藏和四条大型河流水栖寡毛类区系研究. 博士学位论文, 中国科学院水生生物研究所, 武汉. 2011 He X B. Studies on faunae of aquatic Oligochaeta (Annelida) in Tibet and four large rivers of China [D]. Thesis for Doctor of Science, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan. 2011
[20] Morse J C, Yang L F, Tian L X. Aquatic Insects of China Useful for Monitoring Water Quality-Volume 13 [M]. Jiangsu: Hohai University Press. 1994, 1—570
[21] Epler J H. Identification Manual for the Larval Chironomidae (Diptera) of North and South Carolina [M]. A Guide to the Taxonomy of the Midges of the Southeastern United States, Including Florida. Special Publication SJ2001-SP13. North Carolina Department of Environment and Natural Resources, Raleigh, NC, and St. Johns River Water Management District, Palatka, FL. 2001, 1—526
[22] 周长发, 苏翠荣, 归鸿. 中国蜉蝣概述. 北京: 科学出版社. 2015, 1—310 Zhou C F, Su C R, Gui H. Outline of Chinese Mayflies [M]. Beijing: Science Press. 2015, 1—310
[23] 刘月英. 中国经济动物志-淡水软体动物. 北京: 科学出版社. 1979, 1—134 Liu Y Y. Chinese Economic Animal - Freshwater Mollusca [M]. Beijing: Science Press. 1979, 1—134
[24] 刘月英, 张文珍, 王耀先. 医学贝类学. 北京: 海洋出版社. 1993, 1—156 Liu Y Y, Zhang W Z, Wang Y X. Medical Shellfish [M]. Beijing: China Ocean Press. 1993, 1—156
[25] 宋大祥, 冯钟琪. 蚂蟥. 科学出版社. 1978, 1—176 Song D X, Feng Z Q. Grasshopper [M]. Beijing: Science Press. 1978, 1—176
[26] 阎云君, 梁彦龄. 大型底栖动物体长体重关系和生化组成的研究. 华中科技大学学报(自然科学版), 2002, 30(11): 114—116 doi: 10.3321/j.issn:1671-4512.2002.11.038 Yan Y J, Liang Y L. On the relationships body-length and body-weight and the biochemical composition of macrozoobenthos [J]. Journal of Huazhong University of Science and Technology (
Nature Science Edition ) , 2002, 30(11): 114—116 doi: 10.3321/j.issn:1671-4512.2002.11.038 [27] Smith I R. Turbulence in Lakes and Rivers [M]. Amsterdam: Freshwater Biol. Assoc. 1975, 1—79
[28] Statzner B, Gore J A, Resh V H. Hydraulic stream ecology: observed patterns and potential applications [J]. Journal of the North American Benthological Society, 1988, 7(4): 307—360 doi: 10.2307/1467296
[29] 卢金友. 长江泥沙起动流速公式探讨. 长江科学院院报, 1991, 8(4): 57—64 Lu J Y. Study on incipient velocity of sediment transport in the Yangtze River [J]. Journal of Yangtze River Scientific Research Institute, 1991, 8(4): 57—64
[30] 邵学军, 王兴奎. 河流动力学概论. 清华大学出版社. 2005, 1—248 Shao X J, Wang X K. Introduction to River Mechanics[M]. Beijing: Tsinghua University Press. 2005, 1—248
[31] 段学花, 王兆印, 田世民. 河床底质对大型底栖动物多样性影响的野外试验. 生态学报, 2007, 47(9): 1553—1556 Duan X H, Wang Z Y, Tian S M. Benthic macroinvertebrates communities and biodiversity in various stream substrata [J]. Acta Ecologica Sinica, 2007, 47(9): 1553—1556
[32] 潘保柱, 王兆印, 何雪宝. 西江大型底栖动物群落特征研究. 水生生物学报, 2011, 35(5): 851—856 Pan B Z, Wang Z Y, He X B. Studies on assemblage characteristics of macrozoobenthos in the West River [J]. Acta Hydrobiologica Sinica, 2011, 35(5): 851—856
[33] Pan B Z, Wang H Z, Wang H J. A floodplain-scale lake classification based on characteristics of macroinvertebrate assemblages and corresponding environmental properties [J]. Limnologica-Ecology and Management of Inland Waters, 2014, 49: 10—17 doi: 10.1016/j.limno.2014.07.003
[34] Dodds W K. Freshwater Ecology: Concepts and Environmental Applications [M]. Amsterdam: Academic Press. 2002, 1—569
[35] 丁宝清, 刘焕章. 长江流域鱼类食性同资源集团组成特征分析. 四川动物, 2011, 30(1): 31—35 doi: 10.3969/j.issn.1000-7083.2011.01.007 Ding B Q, Liu H Z. Analysis of the fish feeding guiding composition in the Yangtze River [J]. Sichuan Journal of Zoology, 2011, 30(1): 31—35 doi: 10.3969/j.issn.1000-7083.2011.01.007
[36] 鹿志创, 田甲申, 王召会, 等. 应用碳氮稳定同位素技术研究江豚(Neophocaena asiaeorientalis ssp. sunameri). 食性. 生态学报, 2016, 36(1): 69—76 Lu Z C, Tian J S, Wang Z H, et al. Using stable isotope technique to study feeding habits of the finless porpoise (Neophocaena asiaeorientalis ssp. sunameri) [J]. Acta Ecologica Sinica, 2016, 36(1): 69—76
[37] Xie S, Li Z, Liu J, et al. Fisheries of the Yangtze River show immediate impacts of the Three Gorges Dam [J]. Fisheries, 2007, 32(7): 343—344
[38] Zhang G, Wu L, Li H, et al. Preliminary evidence of delayed spawning and suppressed larval growth and condition of the major carps in the Yangtze River below the Three Gorges Dam [J]. Environmental Biology of Fishes, 2012, 93(3): 439—447 doi: 10.1007/s10641-011-9934-8
-
期刊类型引用(8)
1. 公莉,过龙根,尹成杰,杨姣姣,杨雅兰. 洱海西太公鱼和太湖新银鱼生长特性及种群调控效果研究. 水生态学杂志. 2022(01): 117-123 .  百度学术
百度学术
2. 李大命,唐晟凯,刘燕山,谷先坤,刘小维,殷稼雯,张彤晴,潘建林. 基于Cytb基因的江苏省大银鱼种群遗传多样性和遗传结构分析. 上海海洋大学学报. 2021(03): 416-425 . 百度学术
3. 赵旭昊,徐东坡,任泷,徐跑. 基于Ecopath模型的太湖鲢鳙生态容量评估. 中国水产科学. 2021(06): 785-795 . 百度学术
4. 王煜恒,陈军,李华. 大银鱼仔鱼的人工培育技术探究. 南方农业. 2021(29): 178-179 . 百度学术
5. 李大命,唐晟凯,刘燕山,谷先坤,刘小维,李强,高军,张伟,穆新武,张彤晴. 骆马湖大银鱼、太湖新银鱼线粒体Cytb和COⅠ基因序列多态性分析. 江苏农业科学. 2020(11): 47-52 . 百度学术
6. 李大命,唐晟凯,刘燕山,谷先坤,刘小维,殷稼雯,张彤晴,潘建林. 江苏省4个太湖新银鱼种群遗传多样性和遗传结构分析. 渔业科学进展. 2020(05): 52-60 . 百度学术
7. 唐富江,高文燕,李慧琴,刘伟. 大银鱼生物学与渔业生态学研究进展. 水产学报. 2020(12): 2100-2111 . 百度学术
8. 毛志刚,谷孝鸿,龚志军,曾庆飞,陈辉辉,李红敏,张胜宇,穆欢. 洪泽湖鱼类群落结构及其资源变化. 湖泊科学. 2019(04): 1109-1119 . 百度学术
其他类型引用(7)








计量
- 文章访问数: 1930
- HTML全文浏览量: 537
- PDF下载量: 98
- 被引次数: 15