
THE PREDATION SELECTIVITY OF FEROCIOUS FISH (SILURUS ASOTUS AND CHANNA ARGUS) TO PROCAMBARUS CLARKII LARVAE IN POYANG LAKE
-
摘要:
自长江禁渔以来, 鄱阳湖凶猛性鱼类数量呈持续增长的态势, 为明确凶猛性鱼类的捕食活动是否会对克氏原螯虾的种群造成影响, 研究以鲶(Silurus asotus)、乌鳢(Channa argus)为捕食者, 选择8种不同种类的饵料生物, 即3种规格[小: (39.20±1.30) mm; 中: (49.82±2.13) mm; 大: (58.99±1.16) mm]的克氏原螯虾(Procambarus clarkii)幼体、鲤(Cyprinus carpio)、鲫(Carassius auratus)、赤眼鳟(Squaliobarbus curriculus)、食蚊鱼(Gambusia affinis)、鲢(Hypophthalmichthys molitrix)、麦穗鱼(Pseudorasbora parva)和日本沼虾(Macrobrachium nipponense)为猎物, 分别开展鲶和乌鳢对饵料生物种类和克氏原螯虾幼体规格的选择性研究。结果表明: (1)鲶、乌鳢均喜好在夜间捕食, 并且鲶、乌鳢的捕食高峰期均集中在为20: 00—8: 00; (2)多种饵料生物同时存在时,鲶、乌鳢均未捕食克氏原螯虾幼体, 并且对鲤、食蚊鱼和日本沼虾表现出较明显的捕食偏好性。(3)同种规格克氏原螯虾幼体的组合中, 鲶对小规格克氏原螯虾幼体存在明显的捕食偏好性, 同时乌鳢捕食小规格克氏原螯虾幼体的数量也显著高于大规格的数量(P<0.05)。(4)多种规格克氏原螯虾幼体等量混养组合中, 鲶、乌鳢对小规格克氏原螯虾幼体存在明显的捕食偏好性, 并且这种偏好性随克氏原螯虾幼体间规格差距增大而升高。综上, 在多种饵料生物共存时, 克氏原螯虾不是凶猛性鱼类的最佳猎物; 但当其他饵料生物种类及数量较少时, 较小的克氏原螯虾幼体依旧面临较高的捕食威胁。
Abstract:Since the closure of fishing in the Yangtze River, the population of ferocious fish in Poyang Lake has been steadily increasing. In order to investigate whether the Predatory activity of ferocious fish impact the population of Procambarus clarkii, we employed Silurus asotus and Channa argus as predators, using eight different types of bait organisms in this study. Three sizes of P. clarkii larvae (small: 39.20±1.30 mm; medium: 49.82±2.13 mm; large: 58.99±1.16 mm), along with Cyprinus carpio, Carassius auratu, Squaliobarbus curriculus, Mosquitofish, Hypophthalmichthys molitrix, Pseudorasbora parva and Macrobrachium nipponense, were used as prey to examine the species selectivity of S. asotus and C. argus and the size selectivity of P. clarkii larvae. The results revealed that: (1) S. asotus and C. argus exhibited a preference for nocturnal predation, with their peak predation occurring between 20:00—8:00; (2) In the presence of various food organisms simultaneously, S. asotus and C. argus did not prey on P. clarkii larvae, displaying clear predation preference for C. carpio, mosquitofish and M. nipponense. (3) When presented with P. clarkii larvae of the same size, S. asotus exhibited a pronounced predation preference for small size P. clarkii larvae, while C. argus preyed significantly more on small-sized P. clarkii larvae than on large-sized ones (P<0.05). (4) In the equal polyculture combination of various sizes of P. clarkii larvae, both S. asotus and C. argus displayed a clear preference for small size P. clarkii larvae, and this preference intensified with increasing size gap among the P. clarkii larvae. In summary, when multiple food organisms coexist, P. clarkii is not the preferred prey for ferocious fish, however, when the variety and quantity of other food organisms are limited, smaller-sized P. clarkii larvae remain at a high risk of predation.
-
Keywords:
- Poyang Lake /
- Silurus asotus /
- Channa argus /
- Prey /
- Procambarus clarkii larvae /
- Predatory selectivity
-
鄱阳湖为我国最大的淡水湖, 是连接长江的重要湖泊。赣江、抚河、信江、饶河、修水五河汇集于鄱阳湖中, 经调蓄后, 由湖口注入长江。其复杂的生境使鄱阳湖成为众多的淡水鱼类的理想栖息地[1]。自长江禁渔以来, 鄱阳湖中肉食性及杂食性鱼类数量逐渐恢复, 相关调查显示, 2021年监测到鄱阳湖中肉食性鱼类共27种, 其中肉食性及杂食性鱼类生物量比例超6成, 远高于草食性鱼类[2]。
据文献报道, 鄱阳湖南矶山保护区有着丰富的鲶(Silurus asotus)、乌鳢(Channa argus)资源[3], 2021年渔业资源调查显示鲶、乌鳢在鄱阳湖湖区及五河入湖口均有捕获[4]。鲶属鲇形目鲇科, 其分布广、适应性强, 主要以小鱼小虾及水生昆虫为食, 并好食动物内脏, 是我国重要的经济鱼类。乌鳢属鲈形目鳢科, 其生性凶猛、食量大、生长快, 乌鳢食性随其生长发生改变, 仔鱼多以浮游动物为食, 成年乌鳢摄食范围广, 包括鲫、鲤、日本沼虾等杂鱼虾[5]。
克氏原螯虾(Procambarus clarkii)属节肢动物门、甲壳亚门、十足目[6]。20世纪20年代末, 克氏原螯虾由日本传入我国, 现广泛分布于长江流域, 其在鄱阳湖中占据一定的生态位[7]并成为鄱阳湖淡水生态系统的重要组成部分。 2009—2013年鄱阳湖克氏原螯虾捕捞产量已经占到虾类捕捞总产量的50%以上, 平均达2.5×104 t[8]。但2021—2022年资源调查结果表明, 在长江禁捕后, 鄱阳湖克氏原螯虾呈现个体偏小, 资源量显著下降的趋势, 并且在多个采样点均未采集到克氏原螯虾。克氏原螯虾因其独特的生物学特性, 现阶段国内外研究多将其作为捕食者而非被捕食者[9—16], 但捕食压力对甲壳类动物的威胁仍显著存在, 如Minello等[17, 18]研究表明捕食是导致褐对虾(Penaeus aztecus)死亡的主要原因。罗鸣钟等[19]发现乌鳢、大口鲇(Silurus meridionalis)对中华绒螯蟹(Eriocheir sinensis)幼蟹的捕食是导致湖泊中华绒螯蟹捕捞率较低的重要因素之一。自然界中存在的天敌仍会对克氏原螯虾生存造成影响, 其幼体在孵化及蜕壳阶段易遭受来自肉食性鱼类、水鸟、水蛇等捕食者的威胁[20]。口裂大小是影响肉食性鱼类捕食活动的关键因素之一, 是导致捕食者对猎物规格具有显著选择性的根本原因, 当猎物体高或体宽为捕食者口裂的60%时, 为其最佳摄食规格[21, 22]。Marvin等[23]研究发现捕食者对规格较小的对虾存在明显的偏好性。此外, Tyser等[24]发现赞比亚的本地鱼类捕食红螯螯虾(Cherax quadricarinatus)时, 对红螯螯虾规格存在显著选择性。长江禁渔后鄱阳湖肉食性及杂食性鱼类逐渐恢复而克氏原螯虾资源量反而下降, 这可能是日益增加的凶猛性鱼类对克氏原螯虾的捕食活动所造成的, 但有关鄱阳湖凶猛性鱼类对克氏原螯虾的捕食机理及其对不同生长阶段的克氏原螯虾是否存在捕食偏好的研究未见报道。
本研究选择鲶和乌鳢作为试验凶猛性鱼类, 挑选8种生活习性不同的饵料生物及3种规格不同的克氏原螯虾幼体作为被捕食对象, 探讨鄱阳湖两种主要凶猛性鱼类的生物种类选择性及对克氏原螯虾幼体的规格选择性, 以期为鄱阳湖克氏原螯虾资源变动提供科学参考。
1. 材料与方法
1.1 试验材料及前期暂养
试验所需乌鳢和鲶均从鄱阳湖南叽山采样点(28.92°N, 116.29°E)捕捞获得, 克氏原螯虾、鲤(Cyprinus carpio)、鲫(Carassius auratus)、赤眼鳟(Squaliobarbus curriculus)、食蚊鱼(Gambusia affinis)、鲢(Hypophthalmichthys molitrix)、麦穗鱼(Pseudorasbora parva)和日本沼虾(Macrobrachium nipponense)8种饵料生物均来自于江西省水产科学研究所养殖基地。乌鳢、鲶和8种饵料生物分别在室内循环水养殖系统暂养7d以上, 在暂养期间持续充气, 水温为(23±1)℃、pH为7.5±0.2、溶解氧含量为(6±0.5) mg/L。在暂养期间, 乌鳢、鲶每隔1日投喂1次上述8种饵料生物各10尾, 8种饵料生物每日投喂1次商业饲料至饱。投喂后30min, 分别用抄网和虹吸管清理剩余饵料及粪便。
1.2 饵料生物种类选择性实验
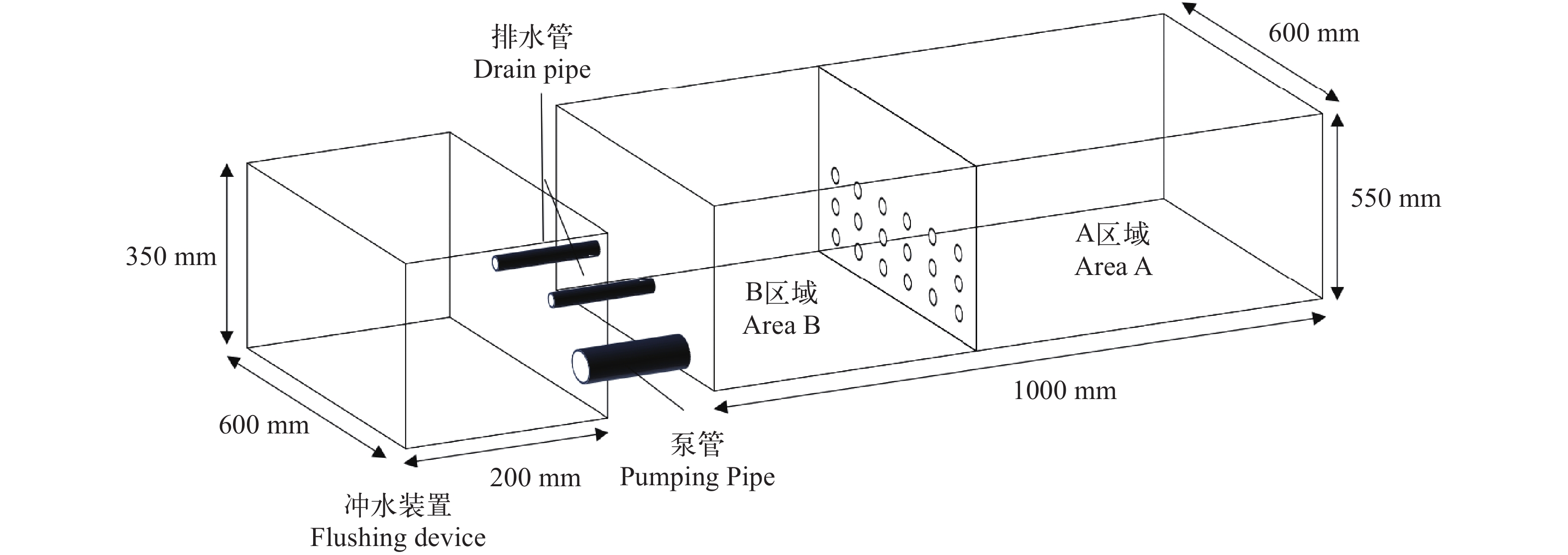
选取经暂养处理后大小均一、活力强、无病无伤的鲶、乌鳢各8尾, 试验前饥饿48h。试验在规格为(1000 mm×600 mm×550 mm), 水深为140 mm的蓝色养殖箱中进行, 在试验水体中加入适量泥沙, 并在养殖箱外接一个小型冲水装置(200 mm×600 mm×350 mm)以模拟鄱阳湖水体透明度及流速, 试验装置如图 1。试验期间8:00开灯、18:00关灯以模拟昼夜交替。在养殖箱1/3处用一块透明的带孔亚克力玻璃板分隔为大小两室, 每个蓝色养殖箱大室内分别放置单种凶猛性鱼类各1尾, 凶猛性鱼类规格见表 1。将不同种饵料生物(克氏原螯虾、鲤、鲫、赤眼鳟、食蚊鱼、鲢、麦穗鱼和日本沼虾)各2尾放置于小室内, 各饵料生物规格见表 2。各处理组重复8次, 对照组大室内不放鱼。适应2h后, 依次抽出隔板开始实验。每间隔4h观察并统计被摄食饵料生物种类及数量, 暗周期用弱光快速照射查看, 24h后试验结束并统计剩余饵料生物种类及数量。
![]() 图 1 捕食选择实验装置A区域为大室, B区域为小室Figure 1. The experimental device of predator selectionArea A is large compartment; Area B is small compartment表 1 鲶、乌鳢全长(TL)、体重(BW)、口裂宽(MB)、口裂长(OFL)Table 1. Total length (TL), body weight (BW), mouth breadth (MB) and oral fissure length (OFL) of S. asotus and C. argus
图 1 捕食选择实验装置A区域为大室, B区域为小室Figure 1. The experimental device of predator selectionArea A is large compartment; Area B is small compartment表 1 鲶、乌鳢全长(TL)、体重(BW)、口裂宽(MB)、口裂长(OFL)Table 1. Total length (TL), body weight (BW), mouth breadth (MB) and oral fissure length (OFL) of S. asotus and C. argus种类
Species数量
Number全长TL
(mm)体重BW
(g)口裂宽
MB (mm)口裂长
OFL (mm)鲶S. asotus 32 286.60±
22.54152.62±
31.1033.22±
3.5627.54±
3.66乌鳢C. argus 32 305.24±
29.57186.62±
34.8634.03±
3.0426.92±
3.27表 2 八种饵料生物全长(TL) 、体重(BW)Table 2. Total length (TL) and body weight (BW) of eight species prey种类
Species样本数
Number全长TL
(mm)体重BW
(g)赤眼鳟S. Curriculus 32 49.30±0.54 0.94±0.05 鲢H. molitrix 32 48.56±1.24 1.23±0.08 食蚊鱼G. affinis 32 31.56±1.47 0.50±0.01 鲫C. auratus 32 43.44±0.71 0.63±0.04 鲤C. carpio 32 45.05±2.14 0.93±0.14 麦穗鱼P. parva 32 49.33±2.81 0.87±0.24 克氏原螯虾P. clarkii 32 40.15±3.49 1.58±0.4 日本沼虾M. nipponense 32 36.07±2.02 1.32±0.18 1.3 克氏原螯虾幼体的规格选择性实验
随机选择21尾经暂养后规格统一、活力强、无病无伤的鲶和乌鳢, 凶猛性鱼类规格如表 1。试验虾根据其体长大小依次分为A (小)、B (中)、C (大)三种不同规格(表 3)。试验条件与试验1.2一致。如表 4所示, 3种不同规格的克氏原螯虾幼体按照不同配比进行组合, 共7种组合方式, 根据组合方式分别设置对应的空白对照组, 各组合中克氏原螯虾幼体数量一致, 为18只, 各试验组与对照组分别设置3个平行。试验开始前, 将经48h饥饿处理的单尾凶猛性鱼类放入图 1所示的养殖箱的大室内, 将不同组合的克氏原螯虾幼体放入小室内。对照组大室内不放鱼。在适应2h后, 依次抽出隔板开始实验, 每间隔12h观察克氏原螯虾幼体被捕食情况, 24h后试验结束并统计剩余克氏原螯虾幼体的数量和规格。
表 3 三种规格克氏原螯虾全长(TL)、体重(BW)、体高(BH)、体宽(BW)Table 3. Total length (TL), body weight (BW), body height (BH) and body width (BW) of P. clarkii with three sizes规格Size 样本数Number 全长TL 体重BW 体高BH 体宽BW 小(A) 252 39.20±1.30 1.74±0.55 10.90±0.02 9.35±0.45 中(B) 252 49.82±2.13 4.89±0.56 13.87±0.90 12.60±0.85 大(C) 252 58.99±1.16 7.20±0.36 15.98±0.25 14.27±0.40 注: A、B、C分别表示平均体长为(39.20±1.30)、(49.82±2.13)和(58.99±1.16) mm的克氏原螯虾幼体; 下同Note: A, B and C represent the larvae of P. clarkii with average body length of (39.20±1.30), (49.82±2.13) and (58.99±1.16) mm, respectively. The same applies below 表 4 克氏原螯虾不同规格间的组合方式Table 4. The combination of different sizes of P. clarkii种类
Species鱼数
Number of fish编号
Number虾的规格Size of P. clarkii 小Small (A) 中Middle (B) 大Large (C) 乌鳢
C. argus1 Ⅰ 18 1 Ⅱ 18 1 Ⅲ 18 1 Ⅳ 9 9 1 Ⅴ 9 9 1 Ⅵ 9 9 1 Ⅶ 6 6 6 鲶
S. asotus1 Ⅰ 18 1 Ⅱ 18 1 Ⅲ 18 1 Ⅳ 9 9 1 Ⅴ 9 9 1 Ⅵ 9 9 1 Ⅶ 6 6 6 注: Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ、Ⅶ分别表示不同规格克氏原螯虾幼虾的7种组合; 下同Note: Ⅰ, Ⅱ, Ⅲ, Ⅳ, Ⅴ, Ⅵ and Ⅶ represent seven combinations of different specifications of P. clarkii. The same applies below 1.4 数据处理与分析
鲶、乌鳢对8种饵料生物和3种规格克氏原螯虾的捕食选择性用Ivlev选择指数Mi进行评价:
$$ M_{i}=(a_{i}-e_{i})/(a_{i}+e_{i}) $$ 式中, Mi为饵料生物i的选择指数, ai为饵料生物i被摄食的量占总摄食量中的比例, ei为饵料生物i的数量占饵料总量中的比例, Mi=0, 表明对该饵料i无偏好性; 0<Mi<1, 表明对该饵料i具有偏好性; –1<Mi<0, 表明不喜好该饵料i。
克氏原螯虾的被捕食数量:
$$ N_{{\rm{a}}}=N_{0}-N_{i} $$ 克氏原螯虾的相对被捕食率:
$$ T=N_{{\rm{a}}}/(N_{0}×D)×100 $$ 式中, N0、Ni分别代表克氏原螯虾起始数量及试验结束后剩余数量, T代表克氏原螯虾的相对被捕食率, D代表捕食者数量。
试验结果以平均值±标准误(mean±SE)表示, 利用SPSS27.0软件对结果进行组内和组间方差分析, 若P<0.05, 则存在显著性差异。使用Graphpad Prism 8进行绘图。
2. 结果
2.1 两种鄱阳湖凶猛性鱼类对饵料生物的种类选择
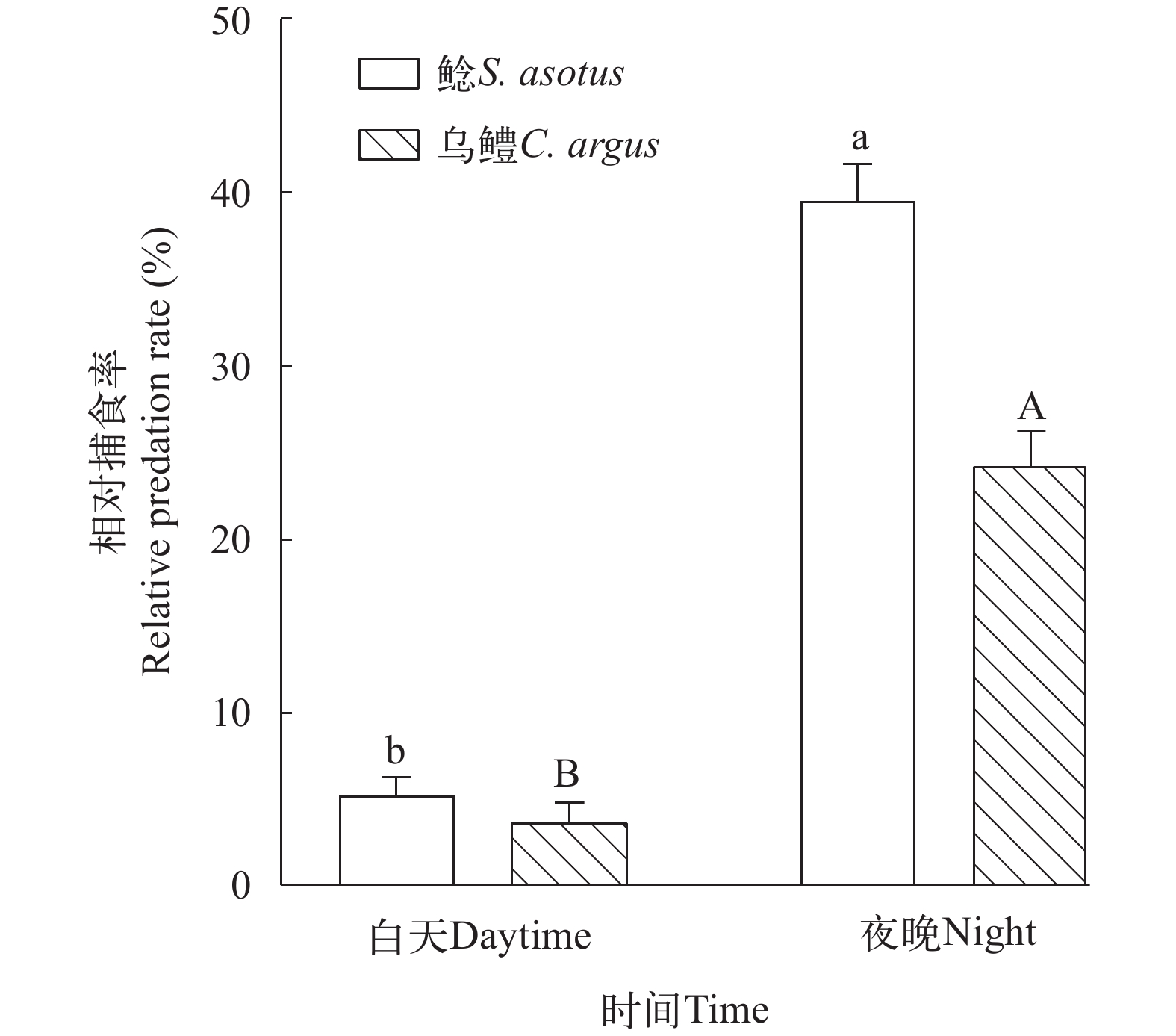
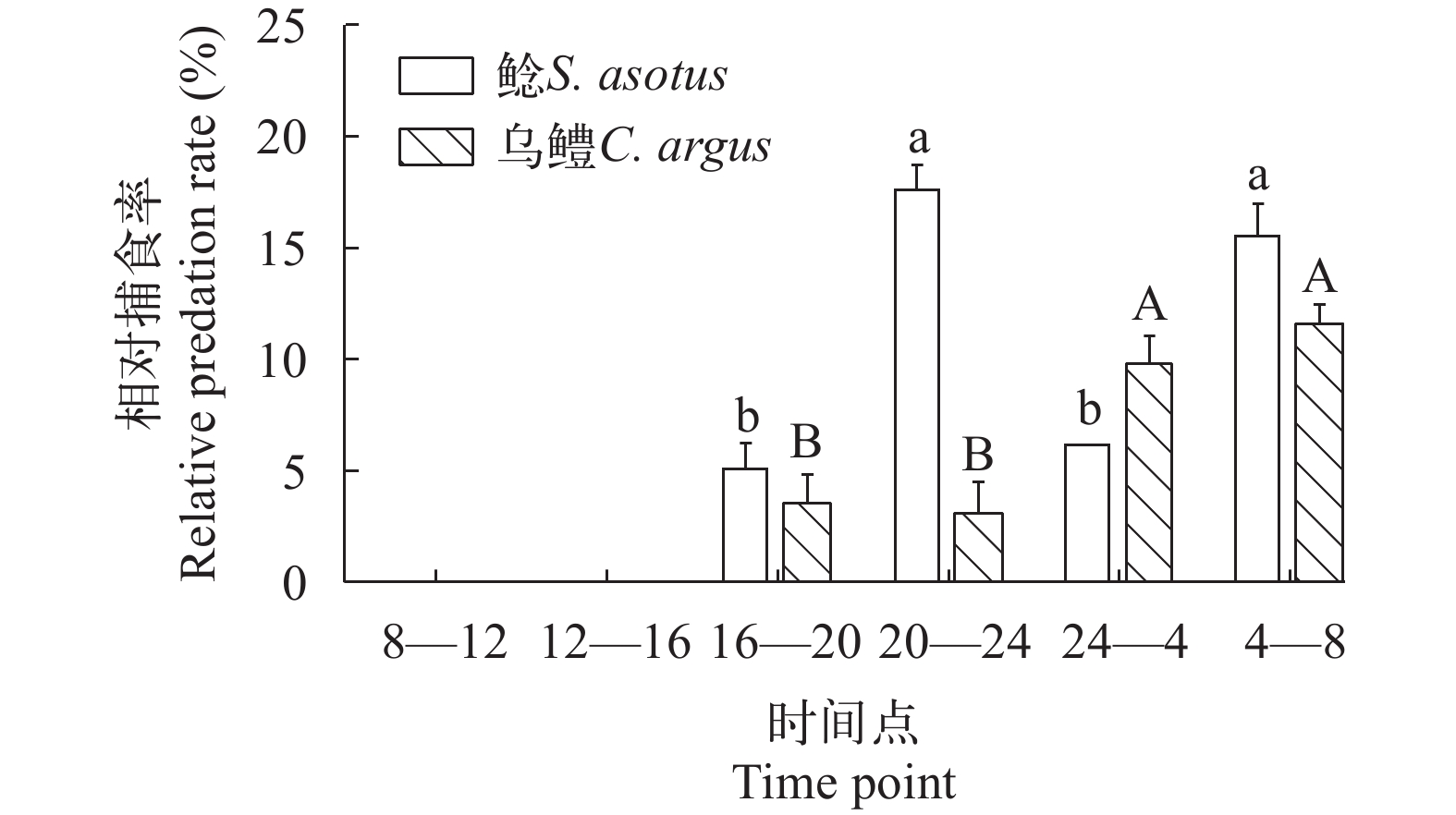
如图 2所示, 鲶、乌鳢在夜晚对饵料生物的相对捕食率显著高于白天(P<0.05)。如图 3所示, 在8:00—12:00和12:00—16:00, 鲶、乌鳢均未摄食。捕食者为鲶时, 在20:00—24:00和4:00—8:00, 鲶对饵料生物的相对捕食率显著大于16:00—20:00和24:00—4:00的捕食率(P<0.0001); 捕食者为乌鳢时, 在24:00—4:00和4:00—8:00, 乌鳢对饵料生物的相对捕食率显著大于16:00—20:00和20:00—24:00的捕食率(P<0.05)。其余时间段间, 鲶、乌鳢对饵料生物的相对捕食率均无显著性差异。
![]() 图 2 鲶、乌鳢在白天、夜晚的相对捕食率误差线表示标准误, 大小写字母不同表明组间存在显著性差异(P<0.05); 下同Figure 2. The predation number of S. asotus and C. argus in daytime and nightError lines indicate standard error, and the difference in case letters indicates that there is a significant difference between groups (P<0.05); The same applies below
图 2 鲶、乌鳢在白天、夜晚的相对捕食率误差线表示标准误, 大小写字母不同表明组间存在显著性差异(P<0.05); 下同Figure 2. The predation number of S. asotus and C. argus in daytime and nightError lines indicate standard error, and the difference in case letters indicates that there is a significant difference between groups (P<0.05); The same applies below![]() 图 3 不同时间段鲶、乌鳢的捕食数量Figure 3. The predation number of S. asotus and C. argus in different time periods
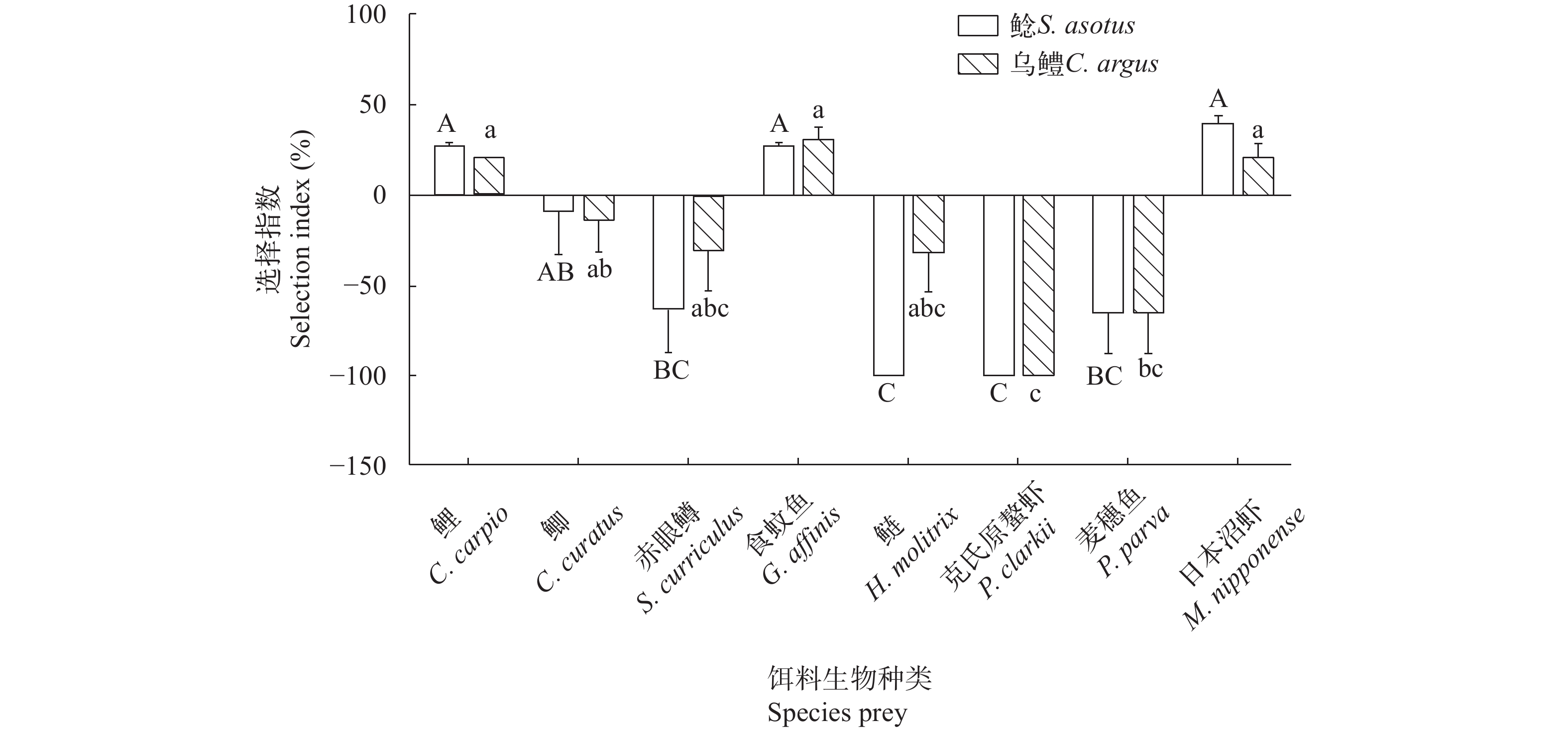
图 3 不同时间段鲶、乌鳢的捕食数量Figure 3. The predation number of S. asotus and C. argus in different time periods除克氏原螯虾之外, 鲶对其他饵料生物均存在捕食, 对食蚊鱼、鲤、日本沼虾的选择指数显著大于麦穗鱼和克氏原螯虾(P<0.05), 对鲫的选择指数显著大于克氏原螯虾(P<0.05), 其余饵料生物间选择指数无显著性差异(图 4)。
![]() 图 4 鲶、乌鳢对不同种类饵料生物的选择指数Figure 4. Selection index of S. asotus and C. argus to different prey species
图 4 鲶、乌鳢对不同种类饵料生物的选择指数Figure 4. Selection index of S. asotus and C. argus to different prey species除鲢、克氏原螯虾之外, 乌鳢对其他饵料生物均存在捕食, 且对食蚊鱼、鲤、日本沼虾的选择指数显著大于除鲫外其余饵料生物的选择指数(P<0.05), 对鲫的选择指数显著大于其对鲢、克氏原螯虾的选择指数(P<0.05), 其余饵料生物间选择指数无显著性差异。对照组中各种饵料生物数量恒定且均未出现死亡。
2.2 两种鄱阳湖凶猛性鱼类对克氏原螯虾幼体的规格选择
如表 5所示, 试验组中, 捕食者为鲶时, 放养单一规格克氏原螯虾幼体的3个组合(Ⅰ、Ⅱ和Ⅲ)间, 鲶对A规格克氏原螯虾幼体的相对捕食率均显著大于B、C两种规格(P<0.05), 其他规格间克氏原螯虾幼体的相对被捕食率均无显著性差异。任意两种规格克氏原螯虾幼体等量混养的3个组合(Ⅳ、Ⅴ和Ⅵ)间, 克氏原螯虾幼体的相对被捕食率间均无显著性差异; 组合内, 鲶对较小规格克氏原螯虾幼体的选择系数均为正, 较大规格均为负, 且组合Ⅴ中, 鲶对A规格克氏原螯虾幼体的相对捕食率显著大于C规格(P<0.05)。3种规格克氏原螯虾幼体等量共存的组合(Ⅶ)中, 鲶对A规格克氏原螯虾幼体的相对捕食率显著大于C规格(P<0.05), 即鲶对小规格克氏原螯虾幼体有较明显的捕食偏好性。
表 5 乌鳢、鲶对克氏原螯虾幼体的捕食选择性Table 5. Predation selectivity of C. argus and S. asotus on juvenile P. clarkii种类
Species组合
CombinationT±SE (%) 合计Sum Mi±SE (%) A B C A B C 鲶S. asotus Ⅰ 16.67±2.08 16.67±2.08A 0 Ⅱ 6.25±0.00 6.25±0.00BC 0 Ⅲ 4.17±2.08 4.17±2.08C 0 Ⅳ 8.33±2.08a 6.25±0.00a 14.58±2.08AB 4.76±4.76 –6.67±4.76 Ⅴ 10.41±2.08a 0.00±0.00b 10.41±2.08ABC 33.33±0.00 –100±0.00 Ⅵ 10.41±2.08a 2.08±2.08a 12.50±3.61ABC 26.98±6.34 –73.33±26.67 Ⅶ 10.41±2.08a 2.08±2.08ab 0.00±0.00b 12.50±0.00ABC 40.00±10.00 –60.00±33.33 –100±0.00 乌鳢C. argus Ⅰ 20.83±2.08 20.83±2.08A 0 Ⅱ 14.58±2.08 14.58±2.08AB 0 Ⅲ 8.33±2.08 8.33±2.08Bc 0 Ⅳ 6.25±0.00a 2.08±2.08a 8.33±2.08Bc 22.22±7.41 –66.67±33.33 Ⅴ 10.41±2.08a 0.00±0.00b 10.41±2.08Bc 33.33±0.00 –100±0.00 Ⅵ 4.17±2.08a 0.00±0.00a 4.17±2.08C 33.33±0.00 –100±0.00 Ⅶ 16.67±2.08a 4.17±2.08b 0.00±0.00b 20.83±2.08A 40.60±4.92 –38.10±31.22 –100±0.00 注: 大小写字母不同分别表明组间和组内存在显著性差异(P<0.05)Note: The difference in case letters indicate significant differences between groups and within groups respectively (P<0.05) 捕食者为乌鳢时, 放养单一规格克氏原螯虾幼体的3个组合(Ⅰ、Ⅱ和Ⅲ)间, 除A和C两种规格克氏原螯虾幼体的相对被捕食率间存在显著性差异(P<0.05), 其他规格间克氏原螯虾幼体的相对被捕食率均无显著性差异。任意两种规格克氏原螯虾幼体等量混养的3个组合(Ⅳ、Ⅴ和Ⅵ)间, 克氏原螯虾幼体的相对被捕食率均无显著性差异; 组合内, 乌鳢对较小规格克氏原螯虾幼体的选择系数均为正, 较大规格均为负, 且组合Ⅴ中, 乌鳢对A规格克氏原螯虾幼体的相对捕食率显著大于C规格(P<0.05)。3种规格克氏原螯虾幼体等量共存的组合(Ⅶ)中, 乌鳢对A规格克氏原螯虾幼体的相对捕食率显著大于B和C规格(P<0.05), 即乌鳢对小规格克氏原螯虾幼体有明显的捕食偏好性。对照组中各规格克氏原螯虾幼体数量恒定且均未出现死亡。
3. 讨论
3.1 鄱阳湖两种主要凶猛性鱼类的摄食节律
影响鱼类捕食活动的因素包括光照强度、水体溶氧量和水温昼夜周期性变化。其中光照强度会直接或间接地控制后者发生变化。不同的光照强度会促进或抑制鱼类捕食, 因此光照强度对鱼类捕食的影响更为显著[25]。在本研究中, 鲶、乌鳢的捕食活动呈现明显的昼夜节律性(鲶的捕食高峰期为20:00—24:00和4:00—8:00, 乌鳢的捕食高峰期为24:00—8:00; 而在8:00—16:00, 鲶、乌鳢均未捕食任何饵料生物)。这与谢从新等[25]所述的乌鳢幼鱼的摄食强度与光照强度呈负相关的结论保持一致。鲶、乌鳢属底层鱼类, 其对光照刺激呈负趋光性, 即较大的光照强度会抑制其行为活动, 如黄颡鱼[26]随光照强度增加, 其游动次数呈现明显的下降趋势。同时, 底层鱼类主要通过嗅觉和机械触碰对猎物进行定位, 其捕食高峰期多为夜间[27]。而一些需通过视觉进行捕食的鱼类如花鲈[28]、漠斑牙鲆[29]等多在白天摄食, 光照能帮助其更好地定位猎物位置, 提高其捕食效率。本研究结果也可能与饵料生物的反捕食能力有关, 饵料生物投入养殖箱初期, 陌生环境导致其反捕食能力增强, 被捕食难度增大; 在适应一定时间后, 饵料生物的警惕性降低, 被捕食难度减小。
3.2 鄱阳湖两种凶猛性鱼类对饵料生物种类选择
捕食者对猎物的搜寻方式、捕食效率和处理能力均会直接影响其自身的食性范围[30], 多种肉食性鱼类如鳜[31]、河川沙塘鳢[32]对猎物种类均表现出明显的捕食选择性。在本研究中, 鲶捕食了除克氏原螯虾外的7种饵料生物, 而乌鳢捕食了除鲢、克氏原螯虾外的6种饵料生物。且两捕食者对鲤、食蚊鱼、日本沼虾均表现出较强的捕食偏好性。该结果可能与克氏原螯虾[33]独特的生物学特性有关, 试验过程中发现克氏原螯虾在感受到捕食威胁后, 长期静止于试验箱角落, 该行为减少了与捕食者的接触机会; 并且克氏原螯虾自身反捕食能力较强, 在面临捕食威胁时, 会摆出防御姿势, 竖起螯足, 使得捕食难度增大[34]。饵料生物是否被捕食与其形态[35]、行为[36]等相关。Webb等[37]研究表明猎物鱼体高与其被捕食难度呈正相关关系。形态为梭形、软鳍条的饵料鱼, 其被捕食的风险越高[38]。在本研究中, 日本沼虾、食蚊鱼规格较其他饵料生物更小, 游泳速度较慢, 反捕食能力较弱, 被捕食的风险更大[39]。而鲢未被乌鳢捕食的原因可能与其习性有关, 鲢较其他种类饵料生物更活跃, 反捕食意识、反捕食能力更强。
3.3 鄱阳湖两种凶猛性鱼类对克氏原螯虾幼体的规格选择性
影响捕食者的捕食选择性的因素包括猎物的种类、运动方式、外形及规格大小。在本研究中, 随着克氏原螯虾幼体规格的增大, 其被鲶、乌鳢捕食的数量显著减少; 当2或3种不同规格的克氏原螯虾幼体等量混养时, 捕食者对小规格克氏原螯虾均表现出明显的捕食偏好性, 并且这种偏好性随克氏原螯虾幼体规格间距增大而升高, 这与乌鳢[40]、中华鳖[41]、尖嘴鲈[42]等捕食者喜好捕食规格较小的猎物的现象一致。这可能是由于随着克氏原螯虾体型增大, 其游泳速度更快, 反捕食意识与能力更强, 被捕食风险更低。从最适摄食原理[43, 44]的角度考虑, 捕食者攻击大规格克氏原螯虾幼体时, 其能力消耗量远大于获取量。而捕食小规格克氏原螯虾幼体能使捕食者的能力收益最大化, 从而导致不同规格的克氏原螯虾幼体间的被捕食量存在差异。且多种肉食性鱼类的捕食活动受其口裂限制[22], 本研究中3种不同规格的克氏原螯虾幼体虽均在鲶、乌鳢的口裂范围之内, 但较大的克氏原螯虾幼体仍可能因其规格而增加捕食者的捕食难度。
3.4 凶猛性鱼类捕食对克氏原螯虾资源的影响探究
在长江禁渔后, 鄱阳湖鱼类生态结构总体呈现以杂食性、中下层鱼类为主[45]。捕食压力对于克氏原螯虾等饵料生物的威胁显著增大。本研究涉及的两种捕食情景, 即多种饵料生物和不同规格克氏原螯虾幼体共存在鄱阳湖中均可能存在。每年4—8月是麦穗鱼[46]、赤眼鳟[47]等鱼类产卵及幼鱼生长的高峰期, 此阶段幼鱼的反捕食能力较弱, 被捕食几率较高, 而此阶段克氏原螯虾多为成虾, 其能通过较强的反捕食能力躲避捕食者的攻击, 捕食者更多地选择捕食难度较低的幼鱼而非克氏原螯虾; 9—次年3月是克氏原螯虾母体产卵及幼体生长发育的关键时期, 此阶段克氏原螯虾幼体反捕食意识及能力较弱, 而其他饵料鱼类规格较幼鱼时期更大, 反捕食能力更强。同时, 冬季鄱阳湖水位较低, 湖区鱼类密度最大, 肉食性鱼类如大口鲇、鳜(Siniperca chuatsi)为冬季鄱阳湖优势种[45], 其需捕食大量食物以应对越冬, 反捕食能力较弱的克氏原螯虾幼体成为其最佳的捕食对象之一; 春季, 鄱阳湖大多数鱼类会前往岸边产卵, 栖息于浅水、滩涂处的克氏原螯虾幼体会面临更大的捕食威胁[48]。综上, 在野外复杂的生态环境中, 捕食威胁可能是导致克氏原螯虾资源量大量损失的原因之一。
4. 结论
本试验结果表明, 当水体中存在大量不同种类的饵料生物时, 鲶、乌鳢对鲤、食蚊鱼和日本沼虾表现出较明显的捕食偏好性, 而克氏原螯虾幼体并不是鲶、乌鳢的最佳捕食对象; 但当其余饵料生物减少时, 鲶、乌鳢对小规格克氏原螯虾幼体表现出较明显的捕食偏好性, 即克氏原螯虾幼体的被捕食风险较高。鄱阳湖中肉食性与杂食性鱼类众多, 其生活习性及捕食方式不尽相同, 除鲶、乌鳢外, 鄱阳湖中其他肉食性鱼类的捕食活动也会影响克氏原螯虾的资源量, 环境因子变化等因素也可能是导致鄱阳湖克氏原螯虾资源量减少的原因。后续需开展不同水层肉食性鱼类的捕食活动对克氏原螯虾的影响研究, 对鄱阳湖水体特征及克氏原螯虾资源变化情况进行持续性跟踪, 多层次、多维度地分析克氏原螯虾资源走势, 探究出影响克氏原螯虾资源量变化的原因, 更好地预测克氏原螯虾的资源变化趋势, 为鄱阳湖克氏原螯虾可持续发展提供科学参考。
-
![]()
图 1 捕食选择实验装置
A区域为大室, B区域为小室
Figure 1. The experimental device of predator selection
Area A is large compartment; Area B is small compartment
![]()
图 2 鲶、乌鳢在白天、夜晚的相对捕食率
误差线表示标准误, 大小写字母不同表明组间存在显著性差异(P<0.05); 下同
Figure 2. The predation number of S. asotus and C. argus in daytime and night
Error lines indicate standard error, and the difference in case letters indicates that there is a significant difference between groups (P<0.05); The same applies below
![]()
图 3 不同时间段鲶、乌鳢的捕食数量
Figure 3. The predation number of S. asotus and C. argus in different time periods
![]()
图 4 鲶、乌鳢对不同种类饵料生物的选择指数
Figure 4. Selection index of S. asotus and C. argus to different prey species
表 1 鲶、乌鳢全长(TL)、体重(BW)、口裂宽(MB)、口裂长(OFL)
Table 1 Total length (TL), body weight (BW), mouth breadth (MB) and oral fissure length (OFL) of S. asotus and C. argus
种类
Species数量
Number全长TL
(mm)体重BW
(g)口裂宽
MB (mm)口裂长
OFL (mm)鲶S. asotus 32 286.60±
22.54152.62±
31.1033.22±
3.5627.54±
3.66乌鳢C. argus 32 305.24±
29.57186.62±
34.8634.03±
3.0426.92±
3.27 下载: 导出CSV
下载: 导出CSV
表 2 八种饵料生物全长(TL) 、体重(BW)
Table 2 Total length (TL) and body weight (BW) of eight species prey
种类
Species样本数
Number全长TL
(mm)体重BW
(g)赤眼鳟S. Curriculus 32 49.30±0.54 0.94±0.05 鲢H. molitrix 32 48.56±1.24 1.23±0.08 食蚊鱼G. affinis 32 31.56±1.47 0.50±0.01 鲫C. auratus 32 43.44±0.71 0.63±0.04 鲤C. carpio 32 45.05±2.14 0.93±0.14 麦穗鱼P. parva 32 49.33±2.81 0.87±0.24 克氏原螯虾P. clarkii 32 40.15±3.49 1.58±0.4 日本沼虾M. nipponense 32 36.07±2.02 1.32±0.18
下载: 导出CSV
表 3 三种规格克氏原螯虾全长(TL)、体重(BW)、体高(BH)、体宽(BW)
Table 3 Total length (TL), body weight (BW), body height (BH) and body width (BW) of P. clarkii with three sizes
规格Size 样本数Number 全长TL 体重BW 体高BH 体宽BW 小(A) 252 39.20±1.30 1.74±0.55 10.90±0.02 9.35±0.45 中(B) 252 49.82±2.13 4.89±0.56 13.87±0.90 12.60±0.85 大(C) 252 58.99±1.16 7.20±0.36 15.98±0.25 14.27±0.40 注: A、B、C分别表示平均体长为(39.20±1.30)、(49.82±2.13)和(58.99±1.16) mm的克氏原螯虾幼体; 下同Note: A, B and C represent the larvae of P. clarkii with average body length of (39.20±1.30), (49.82±2.13) and (58.99±1.16) mm, respectively. The same applies below
下载: 导出CSV
表 4 克氏原螯虾不同规格间的组合方式
Table 4 The combination of different sizes of P. clarkii
种类
Species鱼数
Number of fish编号
Number虾的规格Size of P. clarkii 小Small (A) 中Middle (B) 大Large (C) 乌鳢
C. argus1 Ⅰ 18 1 Ⅱ 18 1 Ⅲ 18 1 Ⅳ 9 9 1 Ⅴ 9 9 1 Ⅵ 9 9 1 Ⅶ 6 6 6 鲶
S. asotus1 Ⅰ 18 1 Ⅱ 18 1 Ⅲ 18 1 Ⅳ 9 9 1 Ⅴ 9 9 1 Ⅵ 9 9 1 Ⅶ 6 6 6 注: Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ、Ⅶ分别表示不同规格克氏原螯虾幼虾的7种组合; 下同Note: Ⅰ, Ⅱ, Ⅲ, Ⅳ, Ⅴ, Ⅵ and Ⅶ represent seven combinations of different specifications of P. clarkii. The same applies below
下载: 导出CSV
表 5 乌鳢、鲶对克氏原螯虾幼体的捕食选择性
Table 5 Predation selectivity of C. argus and S. asotus on juvenile P. clarkii
种类
Species组合
CombinationT±SE (%) 合计Sum Mi±SE (%) A B C A B C 鲶S. asotus Ⅰ 16.67±2.08 16.67±2.08A 0 Ⅱ 6.25±0.00 6.25±0.00BC 0 Ⅲ 4.17±2.08 4.17±2.08C 0 Ⅳ 8.33±2.08a 6.25±0.00a 14.58±2.08AB 4.76±4.76 –6.67±4.76 Ⅴ 10.41±2.08a 0.00±0.00b 10.41±2.08ABC 33.33±0.00 –100±0.00 Ⅵ 10.41±2.08a 2.08±2.08a 12.50±3.61ABC 26.98±6.34 –73.33±26.67 Ⅶ 10.41±2.08a 2.08±2.08ab 0.00±0.00b 12.50±0.00ABC 40.00±10.00 –60.00±33.33 –100±0.00 乌鳢C. argus Ⅰ 20.83±2.08 20.83±2.08A 0 Ⅱ 14.58±2.08 14.58±2.08AB 0 Ⅲ 8.33±2.08 8.33±2.08Bc 0 Ⅳ 6.25±0.00a 2.08±2.08a 8.33±2.08Bc 22.22±7.41 –66.67±33.33 Ⅴ 10.41±2.08a 0.00±0.00b 10.41±2.08Bc 33.33±0.00 –100±0.00 Ⅵ 4.17±2.08a 0.00±0.00a 4.17±2.08C 33.33±0.00 –100±0.00 Ⅶ 16.67±2.08a 4.17±2.08b 0.00±0.00b 20.83±2.08A 40.60±4.92 –38.10±31.22 –100±0.00 注: 大小写字母不同分别表明组间和组内存在显著性差异(P<0.05)Note: The difference in case letters indicate significant differences between groups and within groups respectively (P<0.05)
下载: 导出CSV
-
[1] 张堂林, 李钟杰. 鄱阳湖鱼类资源及渔业利用 [J]. 湖泊科学, 2007, 19(4): 434-444. doi: 10.3321/j.issn:1003-5427.2007.04.012 Zhang T L, Li Z J. Fish resources and fishery utilization of Lake Poyang [J]. Journal of Lake Sciences, 2007, 19(4): 434-444. doi: 10.3321/j.issn:1003-5427.2007.04.012
[2] 张燕萍, 王昌来, 傅培峰, 等. 禁捕期鄱阳湖渔业资源现状 [C]. 江西省水产学会第七次会员代表大会暨2021年学术年会摘要集. 南昌, 2021: 54. Zhang Y P, Wang C L, Fu P F, et al. Current Situation of Fishery Resources in Poyang Lake During the Fishing Ban Period [C]. Summary of the Seventh Member Congress and 2021 Annual Meeting of Jiangxi Fisheries Society. Nanchang, 2021: 54.
[3] 吴文静, 吴志强, 胡茂林. 鄱阳湖南矶山自然保护区鲶、黄颡鱼和乌鳢的同工酶分析 [J]. 水生态学杂志, 2008, 29(5): 103-105. Wu W J, Wu Z Q, Hu M L. Isozyme analysis of Silurus asotus, Pseudobagrus fulvidraco and Ophiocephalus argus in Nanjishan Wetland Natural Reserve in Poyang Lake [J]. Journal of Hydroecology, 2008, 29(5): 103-105.
[4] 吴斌, 章海鑫, 吴子君, 等. 2020-2021年鄱阳湖及其河口渔业资源调查 [J]. 水生态学杂志, 2023: 1-21. Wu B, Zhang H X, Wu Z J, et al. Survey of fisheries resources in Poyang Lake and its estuaries from 2020 to 2021 [J]. Journal of Hydroecology, 2023: 1-21.
[5] 赵永军, 徐文彦, 张慧. 鳜、鳢、鳡、鲶的生态习性 [J]. 水产科学, 2004, 23(6): 26-27. doi: 10.3969/j.issn.1003-1111.2004.06.009 Zhao Y J, Xu W Y, Zhang H. Ecology of Siniperperca Chuatsi, Ophicephalus argu, Elopichthys bambusa, and Silurus asotus [J]. Fisheries Science, 2004, 23(6): 26-27. doi: 10.3969/j.issn.1003-1111.2004.06.009
[6] 唐鑫生. 克氏原螯虾 [J]. 生物学通报, 2001, 36(9): 19-20. doi: 10.3969/j.issn.0006-3193.2001.09.013 Tang X S. Procambarus clarkii [J]. Bulletin of Biology, 2001, 36(9): 19-20. doi: 10.3969/j.issn.0006-3193.2001.09.013
[7] 邹节新, 汪雁, 钟永亮, 等. 鄱阳湖克氏原螯虾的分布现状及其群体外部形态聚类分析 [J]. 长江流域资源与环境, 2014, 23(3): 415-421. doi: 10.11870/cjlyzyyhj201403016 Zou J X, Wang Y, Zhong Y L, et al. Current distribution of Procambarus clarkii (Crustacea, Decapoda, Cambaridae) in Poyang Lake and the preliminary morphological cluster analysis of different populations [J]. Resources and Environment in the Yangtze Basin, 2014, 23(3): 415-421. doi: 10.11870/cjlyzyyhj201403016
[8] 张燕萍, 陈文静, 方春林, 等. 鄱阳湖区克氏原螯虾捕捞种群结构分析 [J]. 江西水产科技, 2014(2): 6-9. doi: 10.3969/j.issn.1006-3188.2014.02.004 Zhang Y P, Chen W J, Fang C L, et al. Analysis on the fishing population structure of Procambarus clarkii in Poyang Lake area [J]. Jiangxi Fishery Science and Technology, 2014(2): 6-9. doi: 10.3969/j.issn.1006-3188.2014.02.004
[9] 樊晓丽, 林植华. 外来捕食者克氏原螯虾对黑斑侧褶蛙不同发育阶段幼体表型与运动能力的影响 [J]. 生态学报, 2020, 40(5): 1731-1739. Fan X L, Lin Z H. Effects of exotic predator Procambarus clarkiion on the phenotype and performance of Pelophylax nigromaculatus larva [J]. Acta Ecologica Sinica, 2020, 40(5): 1731-1739.
[10] 蔡凤金. 克氏原螯虾对几种水生动物的捕食及其对克氏原螯虾的识别机制研究 [D]. 桂林: 广西师范大学, 2011: 16-21. Cai F J. Predation of Procambarus clarkii on several aquatic animals and its recognition mechanism [D]. Guilin: Guangxi Normal University, 2011: 16-21.
[11] 武正军, 蔡凤金, 贾运锋, 等. 桂林地区克氏原螯虾对泽蛙蝌蚪的捕食 [J]. 生物多样性, 2008, 16(2): 150-155. doi: 10.3321/j.issn:1005-0094.2008.02.007 Wu Z J, Cai F J, Jia Y F, et al. Predation impact of Procambarus clarkii on Rana limnocharis tadpoles in Guilin area [J]. Biodiversity Science, 2008, 16(2): 150-155. doi: 10.3321/j.issn:1005-0094.2008.02.007
[12] 毕研云. 对克氏原螯虾捕食钉螺影响因素的研究 [D]. 北京: 首都师范大学, 2007: 4-8. Bi Y Y. Study on influencing factors of crayfish preying on Oncomelania hupensis [D]. Beijing: Capital Normal University, 2007: 4-8.
[13] 戴强, 戴建洪, 李成, 等. 仙姑弹琴蛙蝌蚪对陌生捕食者克氏原螯虾的反捕反应 [J]. 生物多样性, 2004, 12(5): 481-487. doi: 10.3321/j.issn:1005-0094.2004.05.002 Dai Q, Dai J H, Li C, et al. Anti-predator behavior of tadpoles of Rana daunchina to a novel crawfish (Procambarus clarkii) [J]. Chinese Biodiversity, 2004, 12(5): 481-487. doi: 10.3321/j.issn:1005-0094.2004.05.002
[14] Chandler H C, Gorman T A, Haas C A. The effects of crayfish predation and vegetation cover on tadpole growth, survival, and nonlethal injury [J]. Journal of Herpetology, 2016, 50(2): 271-277. doi: 10.1670/14-176
[15] Axelsson E, Nyström P, Sidenmark J, et al. Crayfish predation on amphibian eggs and larvae [J]. Amphibia-Reptilia, 1997, 18(3): 217-228. doi: 10.1163/156853897X00107
[16] Crane A L, Bairos-Novak K R, Jefferson D M, et al. Survival, behaviour, and morphology of larval wood frogs, Lithobates sylvaticus, under threat from an exotic crayfish predator, Orconectes virilis [J]. Aquatic Ecology, 2019, 53(3): 383-392. doi: 10.1007/s10452-019-09696-2
[17] Minello T J, Zimmerman R J. Fish predation on juvenile brown shrimp, Penaeus aztecus Ives: the effect of simulated Spartina structure on predation rates [J]. Journal of Experimental Marine Biology and Ecology, 1983, 72(3): 211-231. doi: 10.1016/0022-0981(83)90107-7
[18] Dall W, Hill B, Rothlisberg P, et al. The biology of the Penaeidae [J]. Journal of Crustacean Biology, 1990: 1-461.
[19] 罗鸣钟, 张堂林, 张超文, 等. 七种捕食性鱼类对中华绒螯蟹幼蟹捕食风险的评估 [J]. 水生生物学报, 2012, 36(5): 932-938. Luo M Z, Zhang T L, Zhang C W, et al. Assessment of predation risks of seven predacious fish on juvenile Chinese mitten crab (Eriocheir sinensis) [J]. Acta Hydrobiologica Sinica, 2012, 36(5): 932-938.
[20] 张开松. 浅谈稻田养殖小龙虾预防天敌的方法 [J]. 河南农业, 2019(28): 52. Zhang K S. Discussion on the methods of preventing natural enemies of crayfish cultured in rice fields [J]. Agriculture of Henan, 2019(28): 52.
[21] Gill A B, Hart P J B. Feeding behaviour and prey choice of the threespine stickleback: the interacting effects of prey size, fish size and stomach fullness [J]. Animal Behaviour, 1994, 47(4): 921-932. doi: 10.1006/anbe.1994.1124
[22] Hoyle J A, Keast A. The effect of prey morphology and size on handling time in a piscivore, the largemouth bass (Micropterus salmoides) [J]. Canadian Journal of Zoology, 1987, 65(8): 1972-1977. doi: 10.1139/z87-300
[23] Mace M M, Rozas L P. Fish predation on juvenile penaeid shrimp: examining relative predator impact and size-selective predation [J]. Estuaries and Coasts, 2018, 41(7): 2128-2134. doi: 10.1007/s12237-018-0409-4
[24] Tyser A, Douthwaite R. Predation on invasive redclaw crayfish Cherax quadricarinatus by native fishes in the Kafue River, Zambia [J]. African Journal of Aquatic Science, 2014, 39(4): 473-477. doi: 10.2989/16085914.2014.980774
[25] 谢从新, 熊传喜, 周洁, 等. 不同光照强度下乌鳢幼鱼的摄食强度及动力学 [J]. 水生生物学报, 1997, 21(3): 213-218. doi: 10.3321/j.issn:1000-3207.1997.03.003 Xie C X, Xiong C X, Zhou J, et al. Feeding intensity and dynamics of juvenile northern snakehead, Channa argus, under different illumination [J]. Acta Hydrobiologica Sinica, 1997, 21(3): 213-218. doi: 10.3321/j.issn:1000-3207.1997.03.003
[26] 王武, 李伟纯, 马旭洲, 等. 水温与光照对瓦氏黄颡鱼幼鱼行为的影响 [J]. 生态学杂志, 2008, 27(5): 791-796. Wang W, Li W C, Ma X Z, et al. Effects of water temperature and light intensity on the behaviors of Pelteobagrus vachelli fingerlings [J]. Chinese Journal of Ecology, 2008, 27(5): 791-796.
[27] 乔志刚, 张国梁, 张英英, 等. 不同光照周期下鲇幼鱼的日摄食节律 [J]. 水产科学, 2008, 27(10): 511-515. doi: 10.3969/j.issn.1003-1111.2008.10.003 Qiao Z G, Zhang G L, Zhang Y Y, et al. Diel feeding rhythm of juvenile oriental sheatfish Silurus asotus in different photoperiods [J]. Fisheries Science, 2008, 27(10): 511-515. doi: 10.3969/j.issn.1003-1111.2008.10.003
[28] 竺俊全, 李明云, 吴锡科. 花鲈仔稚鱼的生长发育与摄食节律研究 [J]. 浙江海洋学院学报(自然科学版), 2002, 21(3): 210-215. Zhu J Q, Li M Y, Wu X K. Study on the growth and feeding rhythm of larval and juvenile of Lateolabrax japonicus [J]. Journal of Zhejiang Ocean University, 2002, 21(3): 210-215.
[29] 秦志清, 林越赳, 张雅芝, 等. 光照对漠斑牙鲆仔鱼摄食、生长与存活的影响 [J]. 集美大学学报(自然科学版), 2009, 14(3): 224-228. Qin Z Q, Lin Y J, Zhang Y Z, et al. Effects of light on feeding, growth and survival of larval of Paralichthys lethostigma [J]. Journal of Jimei University (Natural Science Edition), 2009, 14(3): 224-228.
[30] 尚玉昌. 行为生态学(五): 动物的觅食技巧 [J]. 生态学杂志, 1985, 4(1): 61-64. Shang Y C. Behavioral ecology (5): animals’ foraging skills [J]. Chinese Journal of Ecology, 1985, 4(1): 61-64.
[31] 崔方天, 张堂林, 刘家寿, 等. 鳜对猎物种类选择性的初步研究 [J]. 淡水渔业, 2013, 43(6): 29-34. doi: 10.3969/j.issn.1000-6907.2013.06.005 Cui F T, Zhang T L, Liu J S, et al. A preliminary study on prey type selection in Siniperca chuatsi [J]. Freshwater Fisheries, 2013, 43(6): 29-34. doi: 10.3969/j.issn.1000-6907.2013.06.005
[32] 任艳, 马琴, 刘怡然, 等. 河川沙塘鳢对猎物种类和大小选择性研究 [J]. 水生生物学报, 2022, 46(6): 865-871. doi: 10.7541/2022.2021.0374 Ren Y, Ma Q, Liu Y R, et al. Prey species and size selection of odontobutis potamophila [J]. Acta Hydrobiologica Sinica, 2022, 46(6): 865-871. doi: 10.7541/2022.2021.0374
[33] 王雪婷, 程琛, 黄成. 角落空间与克氏原螯虾生活状况的关系 [J]. 水产科学, 2010, 29(6): 348-351. doi: 10.3969/j.issn.1003-1111.2010.06.009 Wang X T, Cheng C, Huang C. Relationship between corner space and living conditions in red swamp crawfish (Procambarus clarkii) [J]. Fisheries Science, 2010, 29(6): 348-351. doi: 10.3969/j.issn.1003-1111.2010.06.009
[34] 雷景涛, 牟长军, 张保彦, 等. 克氏原螯虾生物学特性探索试验 [J]. 齐鲁渔业, 2010, 27(2): 20-23. Lei J T, Mu C J, Zhang B Y, et al. Experimental study on biological characteristics of Procambarus clarkii [J]. Shandong Fisheries, 2010, 27(2): 20-23.
[35] Moody R C, Helland J M, Stein R A. Escape tactics used by bluegills and fathead minnows to avoid predation by tiger muskellunge [J]. Environmental Biology of Fishes, 1983, 8(1): 61-65. doi: 10.1007/BF00004947
[36] Wahl D H, Stein R A. Selective predation by three esocids: the role of prey behavior and morphology [J]. Transactions of the American Fisheries Society, 1988, 117(2): 142-151. doi: 10.1577/1548-8659(1988)117<0142:SPBTET>2.3.CO;2
[37] Webb P W. Effect of body form and response threshold on the vulnerability of four species of teleost prey attacked by largemouth bass (Micropterus salmoides) [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1986, 43(4): 763-771. doi: 10.1139/f86-094
[38] Einfalt L M, Wahl D H. Prey selection by juvenile walleye as influenced by prey morphology and behavior [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1997, 54(11): 2618-2626. doi: 10.1139/f97-172
[39] Nilsson P, Brönmark C. Prey vulnerability to a gape-size limited predator: behavioural and morphological impacts on northern pike piscivory [J]. Oikos, 2000, 88(3): 539-546. doi: 10.1034/j.1600-0706.2000.880310.x
[40] 罗鸣钟, 吴小平, 张堂林, 等. 饵料鱼大小对乌鳢捕食的影响 [J]. 华中农业大学学报, 2009, 28(6): 726-730. doi: 10.3321/j.issn:1000-2421.2009.06.018 Luo M Z, Wu X P, Zhang T L, et al. Effect of prey fish size on predation of snakehead (Channa argus) [J]. Journal of Huazhong Agricultural University, 2009, 28(6): 726-730. doi: 10.3321/j.issn:1000-2421.2009.06.018
[41] 袁向阳, 朱卫东, 富裕, 等. 中华鳖对日本沼虾幼虾捕食作用的实验研究 [J]. 淡水渔业, 2016, 46(3): 76-81. doi: 10.3969/j.issn.1000-6907.2016.03.013 Yuan X Y, Zhu W D, Fu Y, et al. The experimental study on predatory effect of Trionyx sinensis on juvenile Macrobrachium nipponensis [J]. Freshwater Fisheries, 2016, 46(3): 76-81. doi: 10.3969/j.issn.1000-6907.2016.03.013
[42] Ribeiro F F, Qin J G. Prey size selection and cannibalistic behaviour of juvenile barramundi Lates calcarifer [J]. Journal of Fish Biology, 2015, 86(5): 1549-1566. doi: 10.1111/jfb.12658
[43] Gill A B. The dynamics of prey choice in fish: the importance of prey size and satiation [J]. Journal of Fish Biology, 2003, 63(s1): 105-116. doi: 10.1111/j.1095-8649.2003.00214.x
[44] Pyke G H. Optimal foraging theory: a critical review [J]. Annual Review of Ecology and Systematics, 1984, 15(1): 523-575. doi: 10.1146/annurev.es.15.110184.002515
[45] 李慧峰, 王珂, 余绪俊, 等. 禁渔初期鄱阳湖鱼类时空分布特征 [J]. 水生生物学报, 2023, 47(1): 147-157. doi: 10.7541/2023.2022.0163 Li H F, Wang K, Yu X J, et al. Hydroacoustic surveys on spatial-temporal distribution of fishes during early fishing ban period in Poyang Lake [J]. Acta Hydrobiologica Sinica, 2023, 47(1): 147-157. doi: 10.7541/2023.2022.0163
[46] 雷小青, 严保华, 姚毅, 等. 麦穗鱼生物学特性及繁殖技术研究 [J]. 江西水产科技, 2013(2): 16-18. doi: 10.3969/j.issn.1006-3188.2013.02.005 Lei X Q, Yan B H, Yao Y, et al. Study on biological characteristics and propagation techniques of wheat ear fish [J]. Jiangxi Fishery Science and Technology, 2013(2): 16-18. doi: 10.3969/j.issn.1006-3188.2013.02.005
[47] 向建国, 何福林. 赤眼鳟生物学特性研究 [J]. 淡水渔业, 2006, 36(3): 38-40. doi: 10.3969/j.issn.1000-6907.2006.03.008 Xiang J G, He F L. Study on biological characteristics of red-eyed trout [J]. Freshwater Fisheries, 2006, 36(3): 38-40. doi: 10.3969/j.issn.1000-6907.2006.03.008
[48] 杨少荣, 黎明政, 朱其广, 等. 鄱阳湖鱼类群落结构及其时空动态 [J]. 长江流域资源与环境, 2015, 24(1): 54-64. doi: 10.11870/cjlyzyyhj201501008 Yang S R, Li M Z, Zhu Q G, et al. Spatial and temporal variations of fish assemblages in Poyanghu Lake [J]. Resources and Environment in the Yangtze Basin, 2015, 24(1): 54-64. doi: 10.11870/cjlyzyyhj201501008
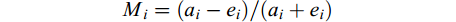
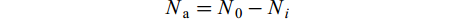
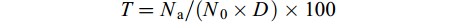
计量
- 文章访问数: 217
- HTML全文浏览量: 36
- PDF下载量: 45